
Disulfiram Ophthalmic Solution Inhibited Macrophage Infiltration by Suppressing Macrophage Pseudopodia Formation in a Rat Corneal Alkali Burn Model
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
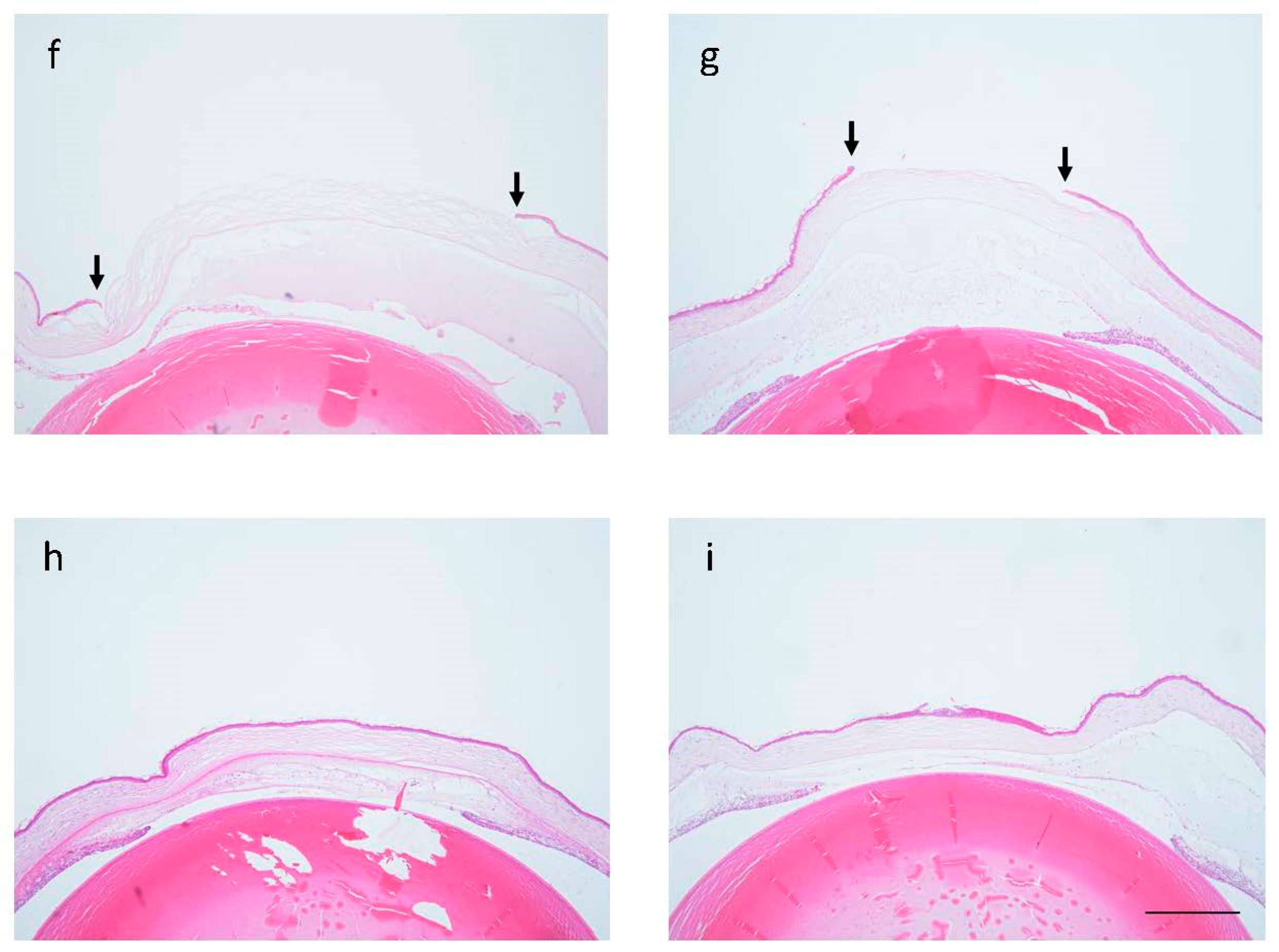
2.1. Corneal Scarring after Alkali Burn
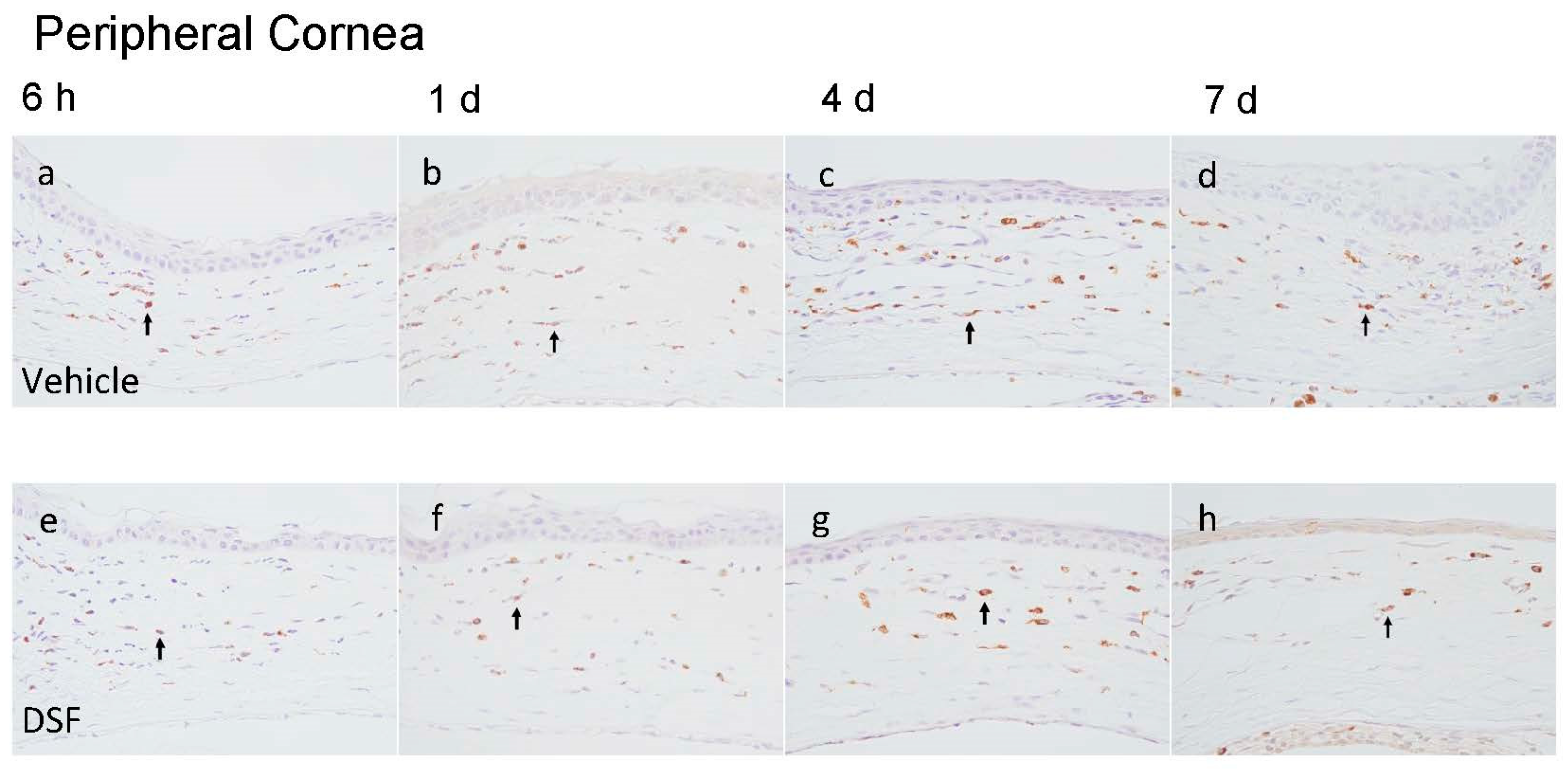
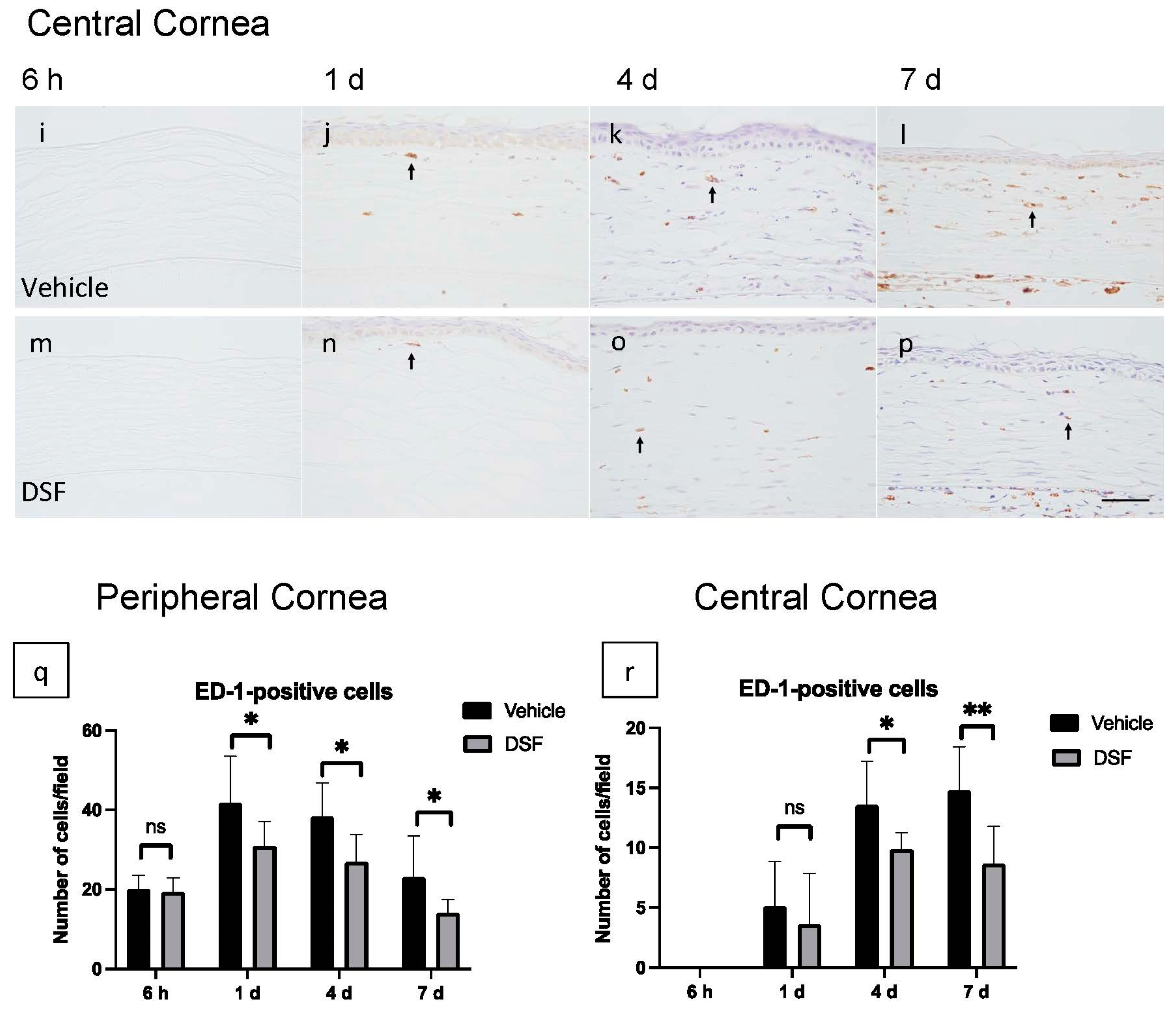
2.2. Evaluation of Macrophage Infiltration with Disulfiram Eye Drops
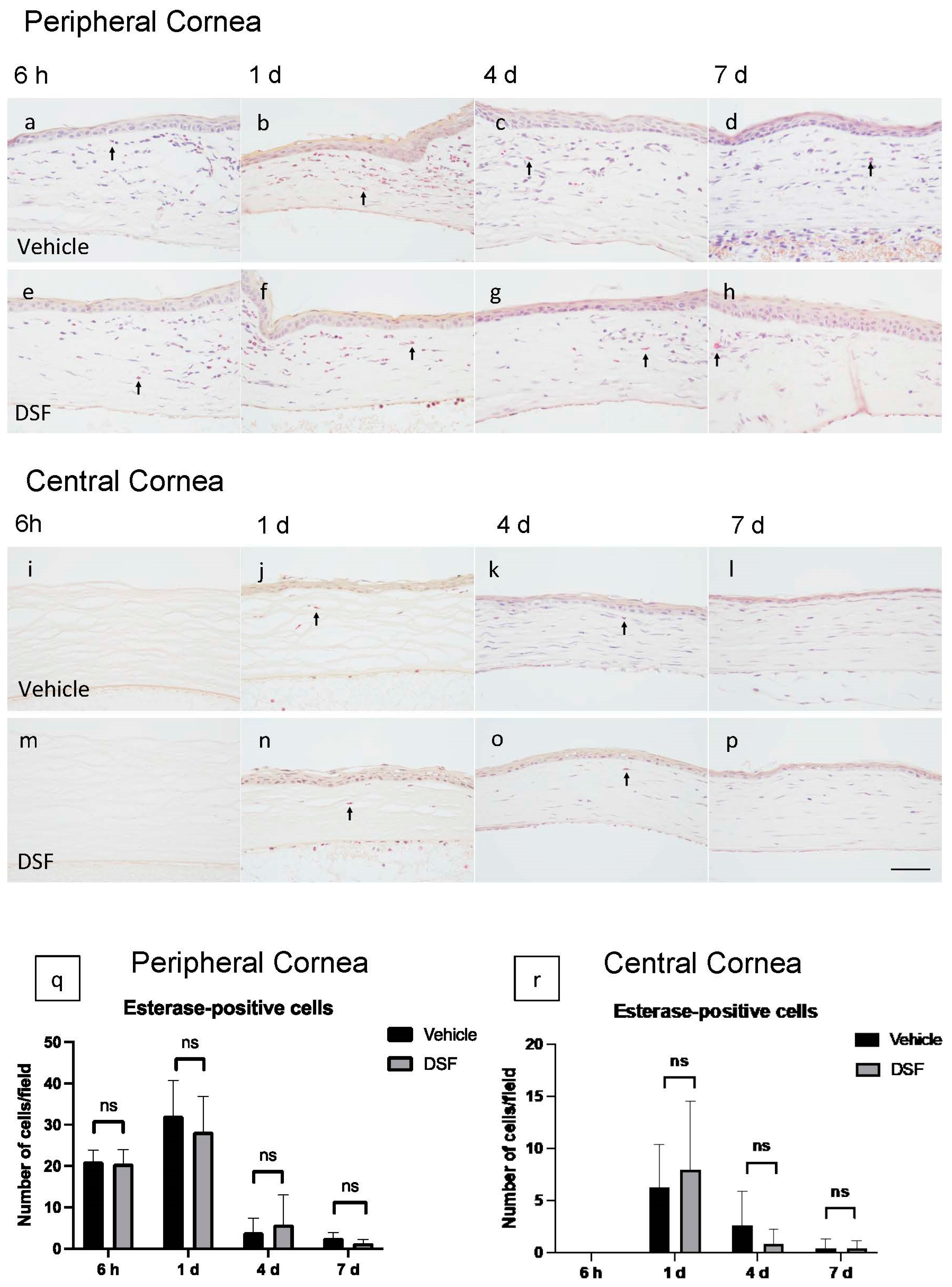
2.3. Evaluation of Neutrophil Infiltration with Disulfiram Eye Drops
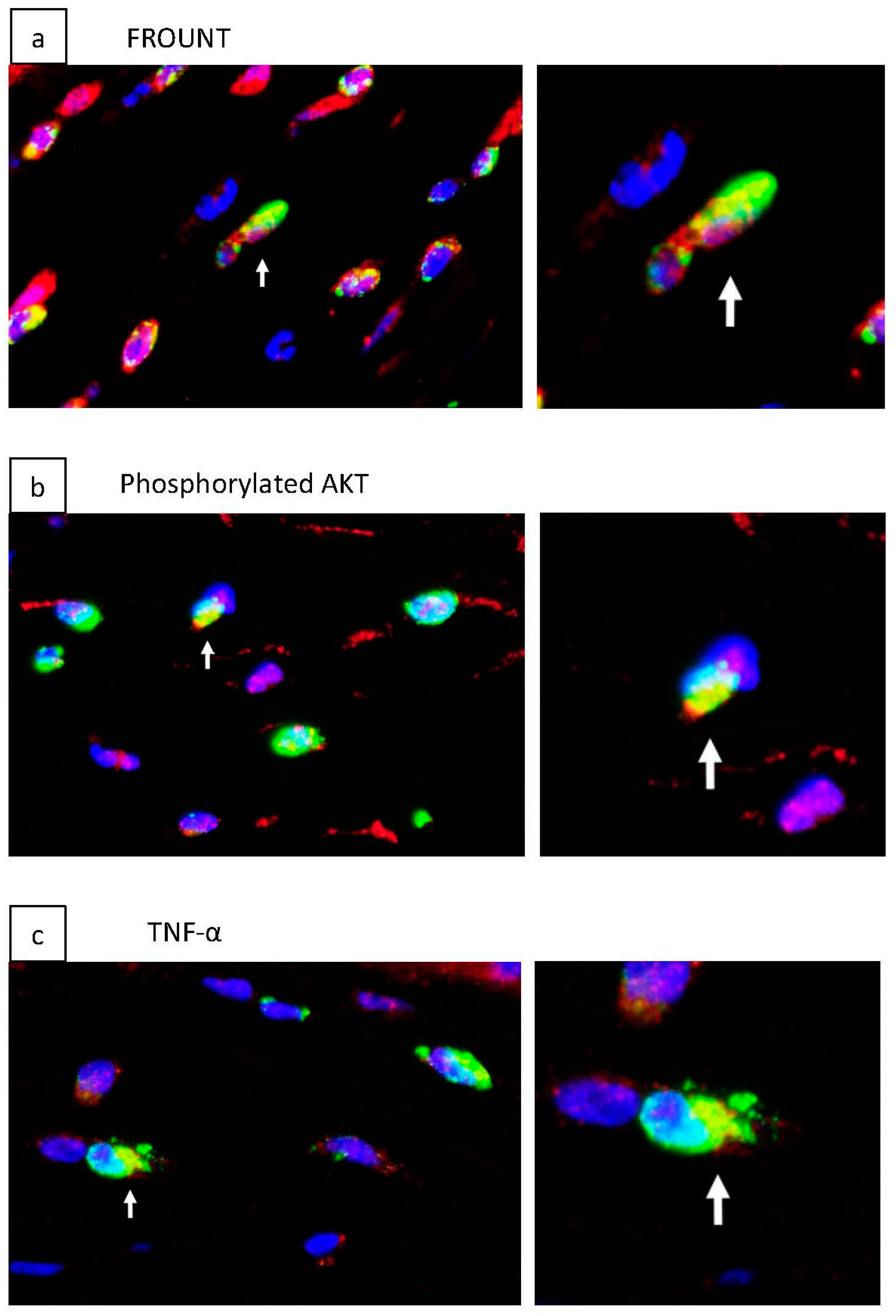
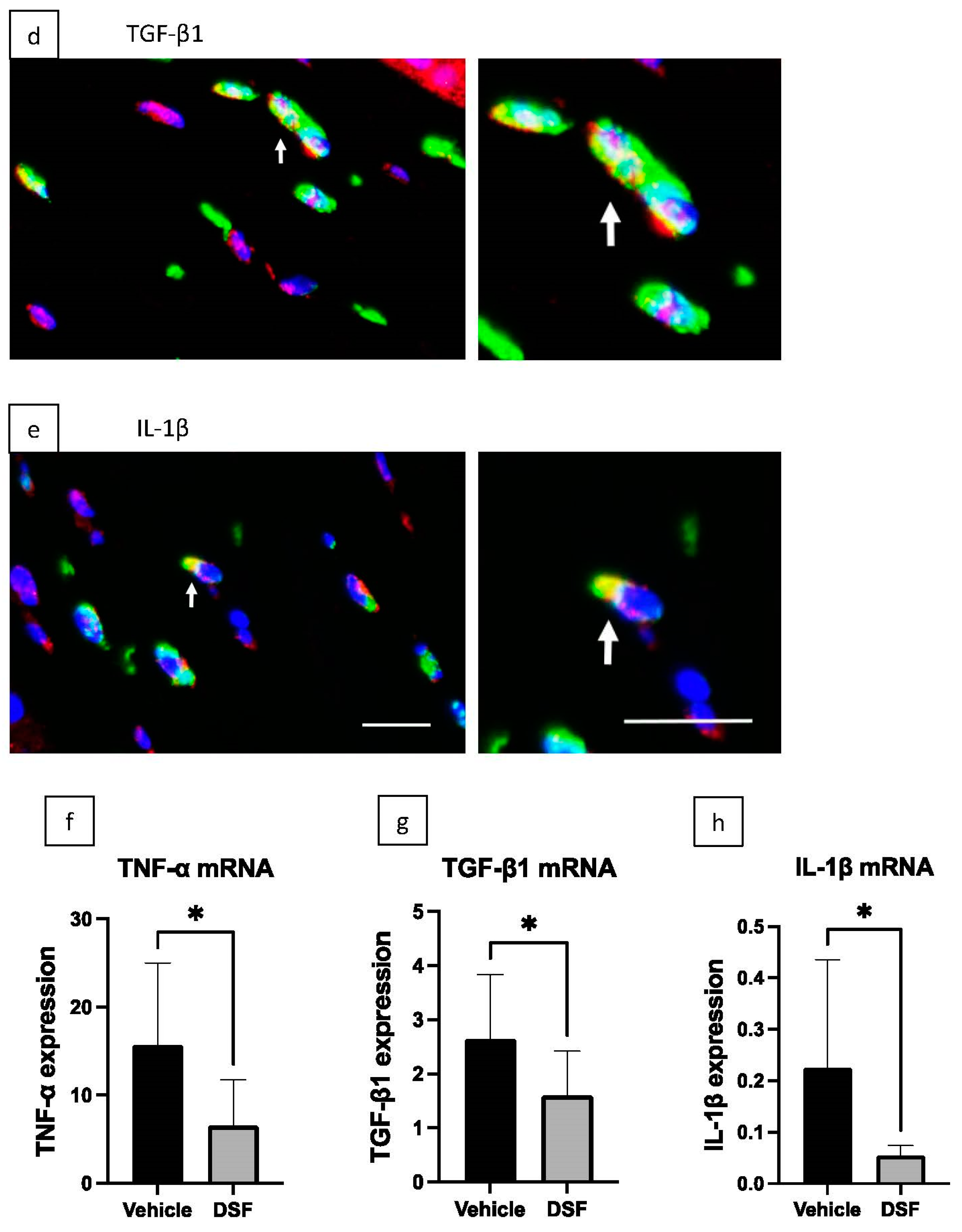
2.4. Inhibition of Macrophage-Associated Cytokines by Disulfiram Eye Drops
2.5. Inhibition of Corneal Scarring by Disulfiram Eye Drops
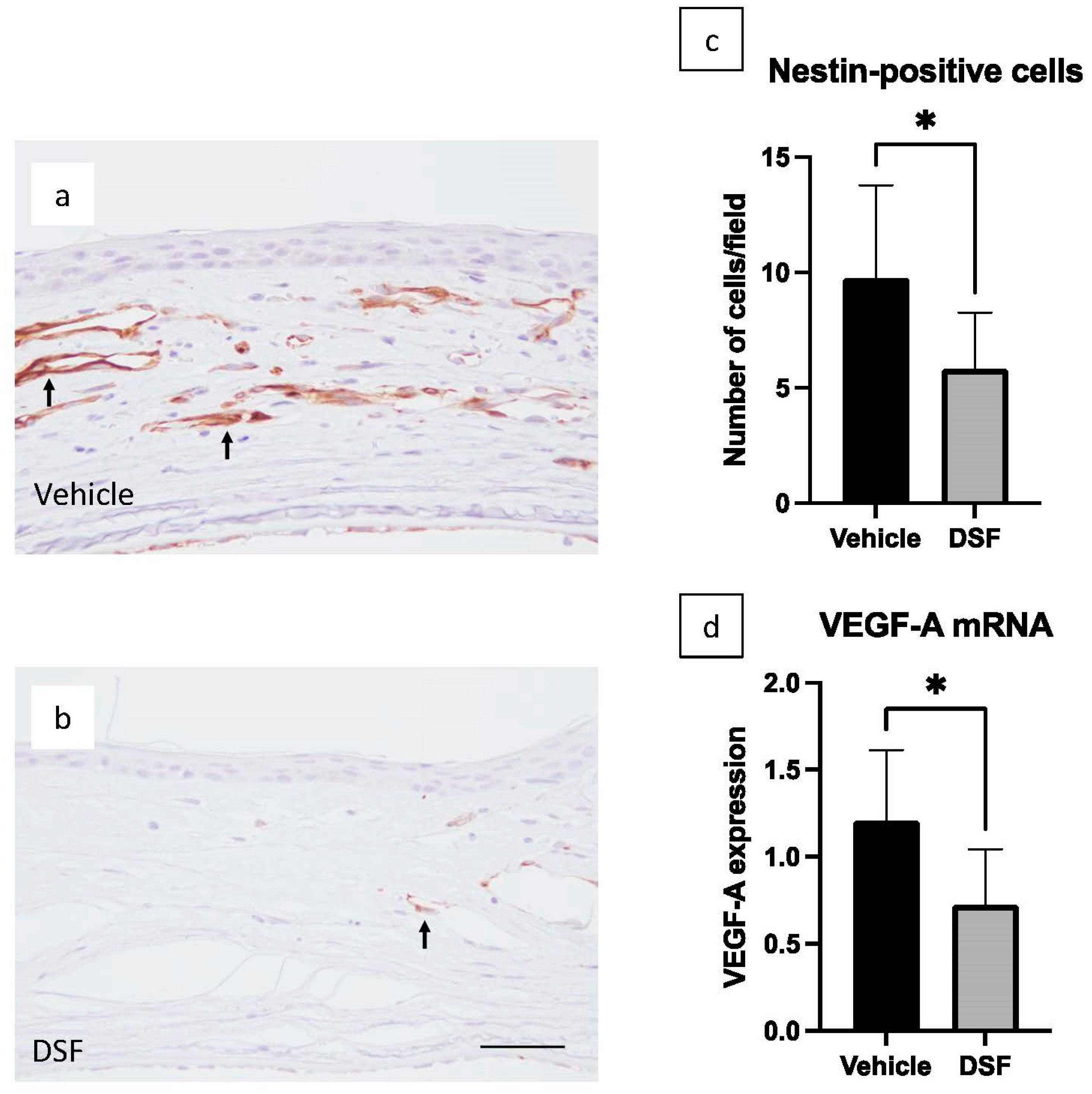
2.6. Inhibition of Angiogenesis by Disulfiram Eye Drops
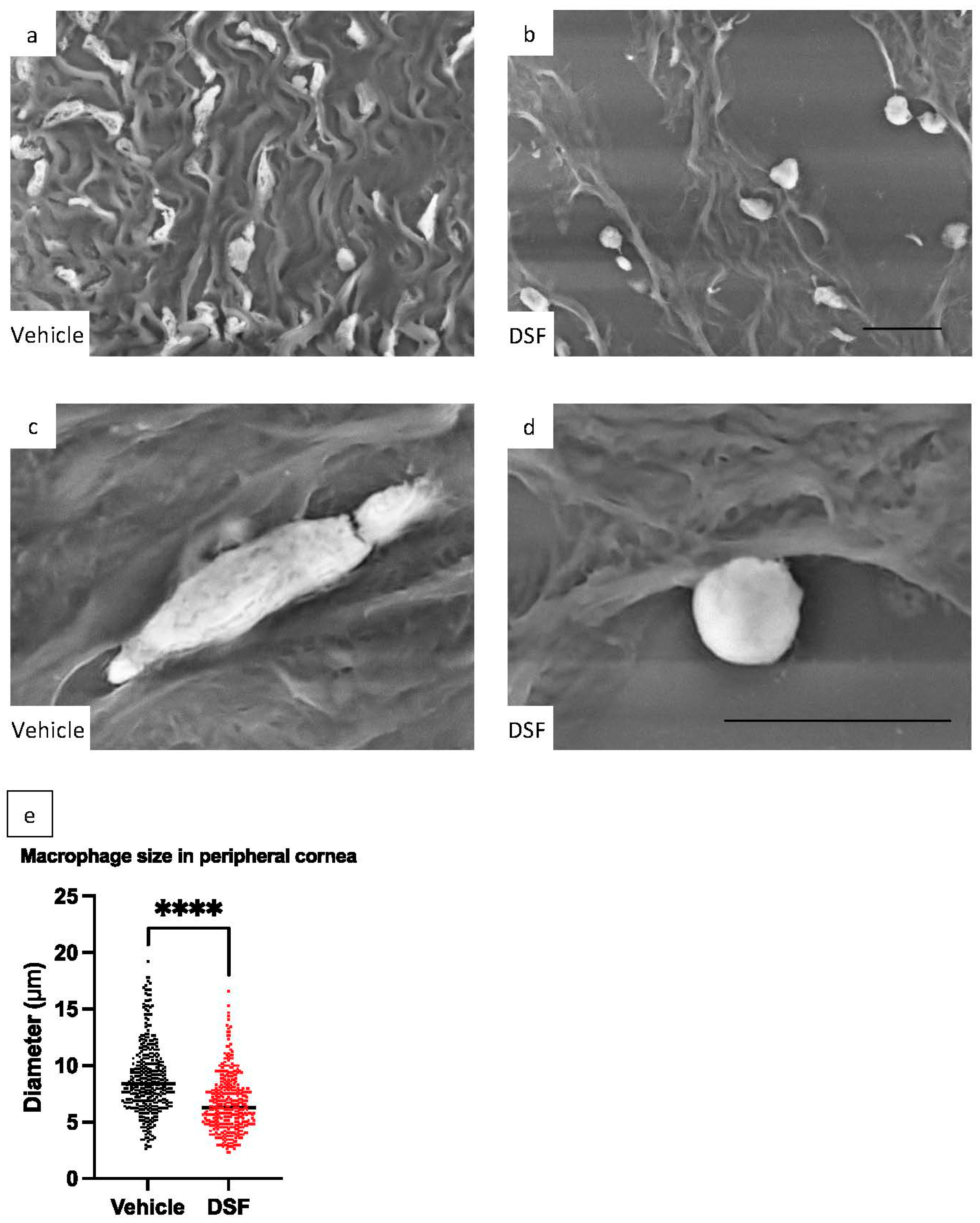
2.7. Evaluation of Macrophage Pseudopodia Formation Using LV-SEM
3. Discussion
4. Materials and Methods
4.1. Animals and Ethics Statement
4.2. Experimental Procedures
4.3. Histological and Immunohistochemical Analyses
4.4. Real-Time RT-PCR
4.5. Low-Vacuum Scanning Electron Microscopy Imaging
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bachelerie, F.; Ben-Baruch, A.; Burkhardt, A.M.; Combadiere, C.; Farber, J.M.; Graham, G.J.; Horuk, R.; Sparre-Ulrich, A.H.; Locati, M.; Luster, A.D.; et al. International Union of Pharmacology. LXXXIX. Update on the extended family of chemokine receptors and introducing a new nomenclature for atypical chemokine receptors. Pharmacol. Rev. 2013, 66, 1–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braga, T.T.; Correa-Costa, M.; Silva, R.C.; Cruz, M.C.; Hiyane, M.I.; da Silva, J.S.; Perez, K.R.; Cuccovia, I.M.; Camara, N.O.S. CCR2 contributes to the recruitment of monocytes and leads to kidney inflammation and fibrosis development. Inflammopharmacology 2018, 26, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Seo, M.J.; Deci, M.B.; Weil, B.R.; Canty, J.M.; Nguyen, J. Effect of CCR2 inhibitor-loaded lipid micelles on inflammatory cell migration and cardiac function after myocardial infarction. Int. J. Nanomed. 2018, 13, 6441–6451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Zhong, B.; He, J.; Yang, X.; He, M.; Zeng, W.; Pan, J.; Fang, Z.; Jia, J.; Liu, H. Single-cell transcriptomes identifies characteristic features of mouse macrophages in liver Mallory-Denk bodies formation. Exp. Mol. Pathol. 2022, 127, 104811. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-K.; Na, Y.H.; Nguyen, H.T.; Nguyen, L.P.; Hurh, S.; Seong, J.Y.; Lee, C.S.; Ham, B.-J.; Hwang, J.-I. Analysis of CCR2 splice variant expression patterns and functional properties. Cell Biosci. 2022, 12, 59. [Google Scholar] [CrossRef] [PubMed]
- She, S.; Ren, L.; Chen, P.; Wang, M.; Chen, D.; Wang, Y.; Chen, H. Functional roles of chemokine receptor CCR2 and its ligands in liver disease. Front Immunol. 2022, 13, 812431. [Google Scholar] [CrossRef]
- Kurihara, T.; Warr, G.; Loy, J.; Bravo, R. Defects in macrophage recruitment and host defense in mice lacking the CCR2 chemokine receptor. J. Exp. Med. 1997, 186, 1757–1762. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Xue, Y.; Dong, D.; Xiao, C.; Lin, C.; Wang, H.; Song, F.; Fu, T.; Wang, Z.; Chen, J.; et al. CCR2- and CCR2+ corneal macrophages exhibit distinct characteristics and balance inflammatory responses after epithelial abrasion. Mucosal Immunol. 2017, 10, 1145–1159. [Google Scholar] [CrossRef] [Green Version]
- Goyal, S.; Chauhan, S.K.; Zhang, Q.; Dana, R. Amelioration of murine dry eye disease by topical antagonist to chemokine receptor 2. Arch. Ophthalmol. 2009, 127, 882–887. [Google Scholar] [CrossRef] [Green Version]
- Oshima, T.; Sonoda, K.-H.; Tsutsumi-Miyahara, C.; Qiao, H.; Hisatomi, T.; Nakao, S.; Hamano, S.; Egashira, K.; Charo, I.F.; Ishibashi, T. Analysis of corneal inflammation induced by cauterisation in CCR2 and MCP-1 knockout mice. Br. J. Ophthalmol. 2006, 90, 218–222. [Google Scholar] [CrossRef]
- Lu, P.; Li, L.; Wu, Y.; Mukaida, N.; Zhang, X. Essential contribution of CCL3 to alkali-induced corneal neovascularization by regulating vascular endothelial growth factor production by macrophages. Mol. Vis. 2008, 14, 1614–1622. [Google Scholar] [PubMed]
- Terashima, Y.; Onai, N.; Murai, M.; Enomoto, M.; Poonpiriya, V.; Hamada, T.; Motomura, K.; Suwa, M.; Ezaki, T.; Haga, T.; et al. Pivotal function for cytoplasmic protein FROUNT in CCR2-mediated monocyte chemotaxis. Nat. Immunol. 2005, 6, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Toda, E.; Terashima, Y.; Esaki, K.; Yoshinaga, S.; Sugihara, M.; Kofuku, Y.; Shimada, I.; Suwa, M.; Kanegasaki, S.; Terasawa, H.; et al. Identification of a binding element for the cytoplasmic regulator FROUNT in the membrane-proximal C-terminal region of chemokine receptors CCR2 and CCR5. Biochem. J. 2014, 457, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Esaki, K.; Yoshinaga, S.; Tsuji, T.; Toda, E.; Terashima, Y.; Saitoh, T.; Kohda, D.; Kohno, T.; Osawa, M.; Ueda, T.; et al. Structural basis for the binding of the membrane-proximal C-terminal region of chemokine receptor CCR2 with the cytosolic regulator FROUNT. FEBS J. 2014, 281, 5552–5566. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, A.; Yoshinaga, S.; Yunoki, K.; Ezaki, S.; Yano, K.; Takeda, M.; Toda, E.; Terashima, Y.; Matsushima, K.; Terasawa, H. Identification and preparation of a novel chemokine receptor-binding domain in the cytoplasmic regulator FROUNT. Mol. Biotechnol. 2017, 59, 141–150. [Google Scholar] [CrossRef]
- Yoshinaga, S.; Ishida, N.; Tsuji, T.; Sonoda, A.; Yunoki, K.; Takeda, M.; Toda, E.; Terashima, Y.; Matsushima, K.; Terasawa, H. 1H, 13C and 15N resonance assignments for a chemokine receptor-binding domain of FROUNT, a cytoplasmic regulator of chemotaxis. Biomol. NMR Assign. 2018, 12, 259–262. [Google Scholar] [CrossRef]
- Terashima, Y.; Toda, E.; Itakura, M.; Otsuji, M.; Yoshinaga, S.; Okumura, K.; Shand, F.H.; Komohara, Y.; Takeda, M.; Kokubo, K.; et al. Targeting FROUNT with disulfiram suppresses macrophage accumulation and its tumor-promoting properties. Nat. Commun. 2020, 11, 609. [Google Scholar] [CrossRef] [Green Version]
- Skrott, Z.; Mistrik, M.; Andersen, K.K.; Friis, S.; Majera, D.; Gursky, J.; Ozdian, T.; Bartkova, J.; Turi, Z.; Moudry, P.; et al. Alcohol-abuse drug disulfiram targets cancer via p97 segregase adaptor NPL4. Nature 2017, 552, 194–199. [Google Scholar] [CrossRef]
- Chen, D.; Cui, Q.C.; Yang, H.; Dou, Q.P. Disulfiram, a clinically used anti-alcoholism drug and copper-binding agent, induces apoptotic cell death in breast cancer cultures and xenografts via inhibition of the proteasome activity. Cancer Res. 2006, 66, 10425–10433. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.J.; Liu, X.; Xia, S.; Zhang, Z.; Zhang, Y.; Zhao, J.; Ruan, J.; Luo, X.; Lou, X.; Bai, Y.; et al. FDA-approved disulfiram inhibits pyroptosis by blocking gasdermin D pore formation. Nat. Immunol. 2020, 21, 736–745. [Google Scholar] [CrossRef]
- Toda, E.; Sawada, A.; Takeuchi, K.; Wakamatsu, K.; Ishikawa, A.; Kuwahara, N.; Sawa, Y.; Hatanaka, S.; Kokubo, K.; Makino, K.; et al. Inhibition of the chemokine signal regulator FROUNT by disulfiram ameliorates crescentic glomerulonephritis. Kidney Int. 2022, 102, 1276–1290. [Google Scholar] [CrossRef] [PubMed]
- Kanai, K.; Ito, Y.; Nagai, N.; Itoh, N.; Hori, Y.; Chikazawa, S.; Hoshi, F.; Higuchi, S. Effects of instillation of eyedrops containing disulfiram and hydroxypropyl-β-cyclodextrin inclusion complex on endotoxin-induced uveitis in rats. Curr. Eye Res. 2012, 37, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Phillips, C.; Oh, J.Y.; Stock, E.M.; Kim, D.-K.; Won, J.-K.; Fulcher, S. Comprehensive modeling of corneal alkali injury in the rat eye. Curr. Eye Res. 2017, 42, 1348–1357. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Kaur, M.; Agarwal, T.; Sangwan, V.S.; Vajpayee, R.B. Treatment of acute ocular chemical burns. Surv. Ophthalmol. 2018, 63, 214–235. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, M.; Shimizu, A.; Masuda, Y.; Nagasaka, S.; Fukuda, Y.; Takahashi, H. An ophthalmic solution of a peroxisome proliferator-activated receptor gamma agonist prevents corneal inflammation in a rat alkali burn model. Mol. Vis. 2013, 19, 2135–2150. [Google Scholar] [PubMed]
- Tanaka, K.; Inagaki, K. Enhancement of the BSE signal from hydrous SEM samples by use of platinum blue. J. Electron. Microsc. 1993, 42, 255. [Google Scholar]
- Inaga, S.; Hirashima, S.; Tanaka, K.; Katsumoto, T.; Kameie, T.; Nakane, H.; Naguro, T. Low vacuum scanning electron microscopy for paraffin sections utilizing the differential stainability of cells and tissues with platinum blue. Arch. Histol. Cytol. 2009, 72, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Arima, T.; Uchiyama, M.; Shimizu, A.; Takahashi, H. Observation of corneal wound healing and angiogenesis using low-vacuum scanning electron microscopy. Transl. Vis. Sci. Technol. 2020, 9, 14. [Google Scholar] [CrossRef]
- Arai, Y.; Takeuchi, K.; Hatanaka, S.; Ishikawa, A.; Inoue, T.; Takakuma, S.; Kajimoto, Y.; Toda, E.; Kunugi, S.; Terasaki, M.; et al. Heavy metal enhancement technique for diaminobenzidine in immunohistochemistry enables ultrastructural observation by low-vacuum scanning electron microscopy. J. Histochem. Cytochem. 2022, 70, 427–436. [Google Scholar] [CrossRef]
- Roberti, G.; Oddone, F.; Agnifili, L.; Katsanos, A.; Michelessi, M.; Mastropasqua, L.; Quaranta, L.; Riva, I.; Tanga, L.; Manni, G. Steroid-induced glaucoma: Epidemiology, pathophysiology, and clinical management. Surv. Ophthalmol. 2020, 65, 458–472. [Google Scholar] [CrossRef]
- Metzemaekers, M.; Gouwy, M.; Proost, P. Neutrophil chemoattractant receptors in health and disease: Double-edged swords. Cell. Mol. Immunol. 2020, 17, 433–450. [Google Scholar] [CrossRef]
- Kamil, S.; Mohan, R.R. Corneal stromal wound healing: Major regulators and therapeutic targets. Ocul. Surf. 2021, 19, 290–306. [Google Scholar] [CrossRef]
- De Oliveira, R.C.; Wilson, S.E. Fibrocytes, Wound Healing, and Corneal Fibrosis. Investig. Ophthalmol. Vis. Sci. 2020, 61, 28. [Google Scholar] [CrossRef] [Green Version]
- Tandon, A.; Tovey, J.C.; Sharma, A.; Gupta, R.; Mohan, R.R. Role of transforming growth factor beta in corneal function, biology and pathology. Curr. Mol. Med. 2010, 10, 565–578. [Google Scholar]
- Myrna, K.E.; Mendonsa, R.; Russell, P.; Pot, S.A.; Liliensiek, S.J.; Jester, J.V.; Nealey, P.F.; Brown, D.; Murphy, C.J. Substratum topography modulates corneal fibroblast to myofibroblast transformation. Investig. Ophthalmol. Vis. Sci. 2012, 53, 811–816. [Google Scholar] [CrossRef] [Green Version]
- Kaji, Y.; Soya, K.; Amano, S.; Oshika, T.; Yamashita, H. Relation between corneal haze and transforming growth factor-beta1 after photorefractive keratectomy and laser in situ keratomileusis. J. Cataract. Refract. Surg. 2001, 27, 1840–1846. [Google Scholar] [CrossRef]
- Stepp, M.A.; Menko, A.S. Immune responses to injury and their links to eye disease. Transl. Res. 2021, 236, 52–71. [Google Scholar] [CrossRef]
- Murray, P.J.; Wynn, T.A. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef] [Green Version]
- Hesketh, M.; Sahin, K.B.; West, Z.E.; Murray, R.Z. Macrophage phenotypes regulate scar formation and chronic wound healing. Int. J. Mol. Sci. 2017, 18, 1545. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, Y.; Hagio, M.; Ishiwata, T. Nestin: A novel angiogenesis marker and possible target for tumor angiogenesis. World J. Gastroenterol. 2013, 19, 42–48. [Google Scholar] [CrossRef]
- Cursiefen, C.; Chen, L.; Borges, L.P.; Jackson, D.; Cao, J.; Radziejewski, C.; D’Amore, P.A.; Dana, M.R.; Wiegand, S.J.; Streilein, J.W. VEGF-A stimulates lymphangiogenesis and hemangiogenesis in inflammatory neovascularization via macrophage recruitment. J. Clin. Investig. 2004, 113, 1040–1050. [Google Scholar] [CrossRef] [Green Version]
- Shakiba, Y.; Mansouri, K.; Arshadi, D.; Rezaei, N. Corneal neovascularization: Molecular events and therapeutic options. Recent Pat. Inflamm. Allergy Drug Discov. 2009, 3, 221–231. [Google Scholar] [CrossRef]
- Lu, P.; Li, L.; Liu, G.; Baba, T.; Ishida, Y.; Nosaka, M.; Kondo, T.; Zhang, X.; Mukaida, N. Critical role of TNF-alpha-induced macrophage VEGF and iNOS production in the experimental corneal neo-vascularization. Investig. Ophthalmol. Vis. Sci. 2012, 53, 3516–3526. [Google Scholar] [CrossRef] [Green Version]
- Mantsounga, C.S.; Lee, C.; Neverson, J.; Sharma, S.; Healy, A.; Berus, J.M.; Parry, C.; Ceneri, N.M.; López-Giráldez, F.; Chun, H.J.; et al. Macrophage IL-1β promotes arteriogenesis by autocrine STAT3- and NF-κB-mediated transcription of pro-angiogenic VEGF-A. Cell Rep. 2022, 38, 110309. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (5′-3′) |
|---|---|---|
| β-actin | GCAGGAGTACGATGAGTCCG | ACGCAGCTCAGTAACAGTCC |
| TNF-α | AAATGGGCTCCCTCTCATCAGTTC | TCTGCTTGGTGGTTTGCTACGAC |
| TGF-b1 | TGGCCAGATCCTGTCCAAAC | GTTGTACAAAGCGAGCACCG |
| IL-1b | TACCTATGTCTTGCCCGTGGAG | ATCATCCCACGAGTCACAGAGG |
| VEGF-A | GCAGCGACAAGGCAGACTAT | GCAACCTCTCCAAACCGTTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ikebukuro, T.; Arima, T.; Kasamatsu, M.; Nakano, Y.; Tobita, Y.; Uchiyama, M.; Terashima, Y.; Toda, E.; Shimizu, A.; Takahashi, H. Disulfiram Ophthalmic Solution Inhibited Macrophage Infiltration by Suppressing Macrophage Pseudopodia Formation in a Rat Corneal Alkali Burn Model. Int. J. Mol. Sci. 2023, 24, 735. https://doi.org/10.3390/ijms24010735
Ikebukuro T, Arima T, Kasamatsu M, Nakano Y, Tobita Y, Uchiyama M, Terashima Y, Toda E, Shimizu A, Takahashi H. Disulfiram Ophthalmic Solution Inhibited Macrophage Infiltration by Suppressing Macrophage Pseudopodia Formation in a Rat Corneal Alkali Burn Model. International Journal of Molecular Sciences. 2023; 24(1):735. https://doi.org/10.3390/ijms24010735
Chicago/Turabian StyleIkebukuro, Toyo, Takeshi Arima, Momoko Kasamatsu, Yuji Nakano, Yutaro Tobita, Masaaki Uchiyama, Yuya Terashima, Etsuko Toda, Akira Shimizu, and Hiroshi Takahashi. 2023. "Disulfiram Ophthalmic Solution Inhibited Macrophage Infiltration by Suppressing Macrophage Pseudopodia Formation in a Rat Corneal Alkali Burn Model" International Journal of Molecular Sciences 24, no. 1: 735. https://doi.org/10.3390/ijms24010735
APA StyleIkebukuro, T., Arima, T., Kasamatsu, M., Nakano, Y., Tobita, Y., Uchiyama, M., Terashima, Y., Toda, E., Shimizu, A., & Takahashi, H. (2023). Disulfiram Ophthalmic Solution Inhibited Macrophage Infiltration by Suppressing Macrophage Pseudopodia Formation in a Rat Corneal Alkali Burn Model. International Journal of Molecular Sciences, 24(1), 735. https://doi.org/10.3390/ijms24010735