
Cycloartenyl Ferulate Is the Predominant Compound in Brown Rice Conferring Cytoprotective Potential against Oxidative Stress-Induced Cytotoxicity
 and
and
Abstract
:1. Introduction
2. Results
2.1. Estimation of the Contribution of Major Phytochemicals to Antioxidant Capacity in Brown Rice
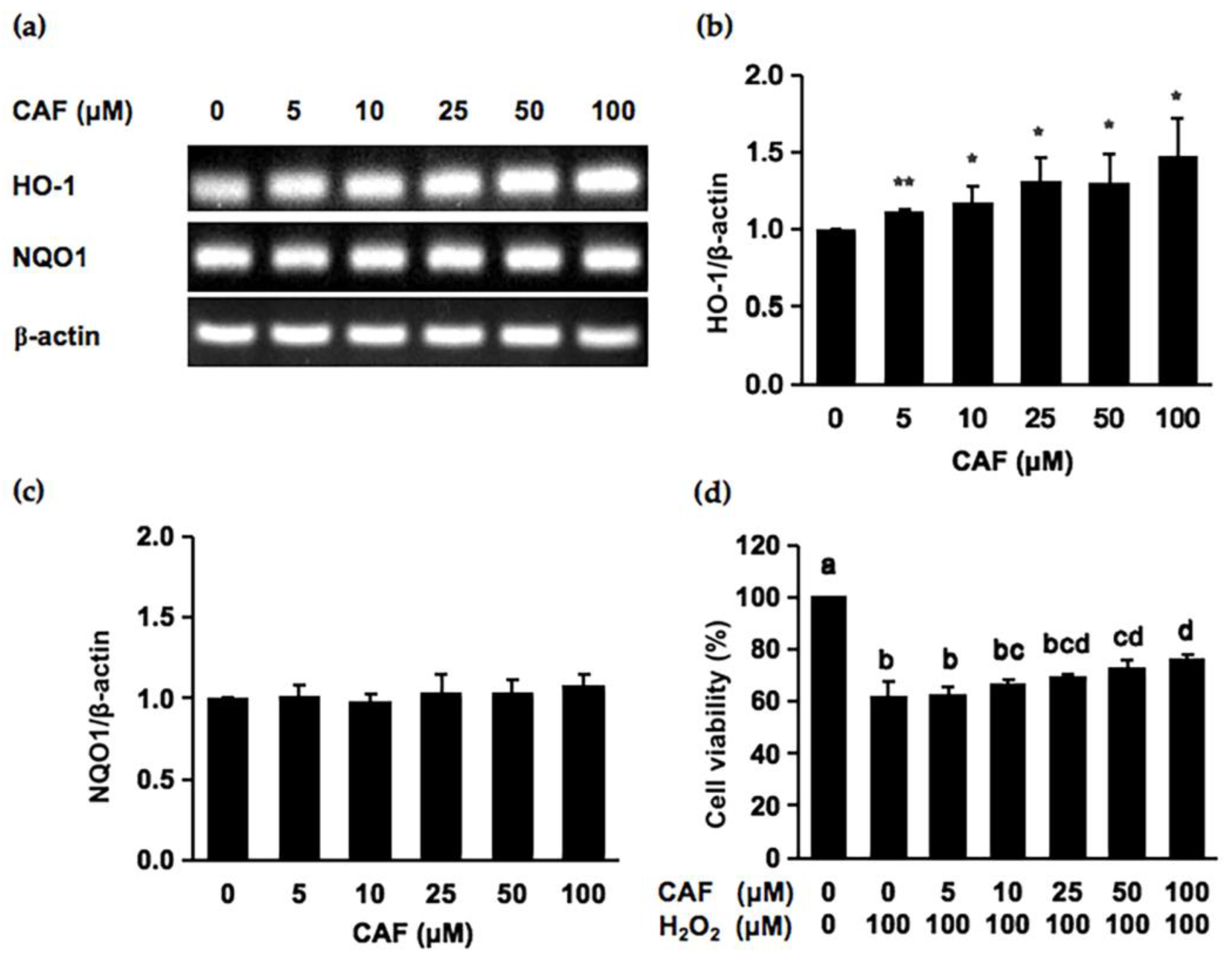
2.2. Cycloartenyl Ferulate (CAF) Is a Potential Cytoprotector against Hydrogen Peroxide
2.3. The Cytoprotective Effect of CAF Was Counteracted by an Heme Oxygenase-1 (HO-1) Inhibitor, Tin Protoporphyrin IX (SnPP)
2.4. Cycloartenyl Moiety Mainly Contributes to the Cytoprotective Potential of CAF
3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Preparation of the Rice Extract
4.3. DPPH Assay
4.4. HPLC Determination of Major Compounds in the Brown Rice
4.5. Cell Cultures
4.6. MTT Assay for Cell Viability Determination
4.7. RNA Extraction and Reverse Transcription-PCR Analysis
4.8. Western Blotting Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cordero-Lara, K.I. Temperate japonica rice (Oryza sativa L.) breeding: History, present and future challenges. Chil. J. Agric. Res. 2020, 80, 303–314. [Google Scholar] [CrossRef]
- Golzarand, M.; Toolabi, K.; Eskandari Delfan, S.; Mirmiran, P. The effect of brown rice compared to white rice on adiposity indices, lipid profile, and glycemic markers: A systematic review and meta-analysis of randomized controlled trials. Crit. Rev. Food Sci. Nutr. 2022, 62, 7395–7412. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, M.; Hossain, S.; Hussain, J.; Hasan, M.; Bhowmick, S.; Basunia, M.A.; Hashimoto, M. Cholesterol lowering and antioxidative effect of pregerminated brown rice in hypercholesterolemic rats. J. Nutr. Sci. Vitaminol. 2019, 65, S93–S99. [Google Scholar] [CrossRef] [Green Version]
- Feng, M.; Wang, X.; Xiong, H.; Qiu, T.; Zhang, H.; Guo, F.; Jiang, L.; Sun, Y. Anti-inflammatory effects of three selenium-enriched brown rice protein hydrolysates in LPS-induced RAW264. 7 macrophages via NF-κB/MAPKs signaling pathways. J. Funct. Foods 2021, 76, 104320. [Google Scholar] [CrossRef]
- Wu, H.; Nakamura, T.; Guo, Y.; Hirooka, M.; Zhang, G.; Munemasa, S.; Murata, Y.; Fujita, A.; Nakamura, Y. White rice ethanol extract is qualitatively, but not quantitatively, equivalent to that of brown rice as an antioxidant source. Biosci. Biotech. Biochem. 2021, 85, 2161–2168. [Google Scholar] [CrossRef]
- Wu, H.; Nakamura, T.; Nakamura, Y. Predominant rice phytochemicals and their disease-preventive effects. JESSS 2021, 10, MR01_p1–MR01_p4. [Google Scholar] [CrossRef]
- Gong, E.S.; Luo, S.J.; Li, T.; Liu, C.M.; Zhang, G.W.; Chen, J.; Zeng, Z.C.; Liu, R.H. Phytochemical profiles and antioxidant activity of brown rice varieties. Food Chem. 2017, 227, 432–443. [Google Scholar] [CrossRef]
- Miller, A.; Engel, K.H. Content of γ-oryzanol and composition of steryl ferulates in brown rice (Oryza sativa L.) of European origin. J. Agric. Food Chem. 2006, 54, 8127–8133. [Google Scholar] [CrossRef]
- Pahrudin Arrozi, A.; Shukri, S.N.S.; Wan Ngah, W.Z.; Mohd Yusof, Y.A.; Ahmad Damanhuri, M.H.; Jaafar, F.; Makpol, S. Comparative effects of alpha-and gamma-tocopherol on mitochondrial functions in Alzheimer’s disease in vitro model. Sci. Rep. 2020, 10, 8962. [Google Scholar] [CrossRef]
- Zhang, C.; Liang, W.; Wang, H.; Yang, Y.; Wang, T.; Wang, S.; Wang, X.; Wang, Y.; Feng, H. γ-Oryzanol mitigates oxidative stress and prevents mutant SOD1-Related neurotoxicity in Drosophila and cell models of amyotrophic lateral sclerosis. Neuropharmacology 2019, 160, 107777. [Google Scholar] [CrossRef]
- Wang, O.; Liu, J.; Cheng, Q.; Guo, X.; Wang, Y.; Zhao, L.; Zhou, F.; Ji, B. Effects of ferulic acid and γ-oryzanol on high-fat and high-fructose diet-induced metabolic syndrome in rats. PloS ONE 2015, 10, e0118135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Xi, X.; Liu, Y.; Lu, Y.; Che, F.; Gu, Y.; Yu, Y.; Li, H.; Liu, J.; Wei, Y. Isolation of four major compounds of γ-oryzanol from rice bran oil by ionic liquids modified high-speed countercurrent chromatography and antimicrobial activity and neuroprotective effect of cycloartenyl ferulate in vitro. Chromatographia 2021, 84, 635–644. [Google Scholar] [CrossRef]
- Chotimarkorn, C.; Ushio, H. The effect of trans-ferulic acid and gamma-oryzanol on ethanol-induced liver injury in C57BL mouse. Phytomedicine 2008, 15, 951–958. [Google Scholar] [CrossRef]
- Kruk, J.; Aboul-Enein, H.Y.; Kładna, A.; Bowser, J.E. Oxidative stress in biological systems and its relation with pathophysiological functions: The effect of physical activity on cellular redox homeostasis. Free Radic. Res. 2019, 53, 497–521. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chang, X.; Zhan, H.; Zhang, Q.; Li, C.; Gao, Q.; Yang, M.; Luo, Z.; Li, S.; Sun, Y. Curcumin and Baicalin ameliorate ethanol-induced liver oxidative damage via the Nrf2/HO-1 pathway. J. Food Biochem. 2020, 44, e13425. [Google Scholar] [CrossRef]
- Wei, J.; Zhao, Q.; Zhang, Y.; Shi, W.; Wang, H.; Zheng, Z.; Meng, L.; Xin, Y.; Jiang, X. Sulforaphane-mediated Nrf2 activation prevents radiation-induced skin injury through inhibiting the oxidative stress-activated DNA damage and NLRP3 inflammasome. Antioxidants 2021, 10, 1850. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, H.L.; Xing, G.D.; Qian, Y.; Zhong, J.F.; Chen, K.L. S-allyl cysteine ameliorates heat stress-induced oxidative stress by activating Nrf2/HO-1 signaling pathway in BMECs. Toxicol. Appl. Pharmacol. 2021, 416, 115469. [Google Scholar] [CrossRef]
- Nakamura, Y.; Miyoshi, N. Electrophiles in foods: The current status of isothiocyanates and their chemical biology. Biosci. Biotech. Biochem. 2010, 74, 242–255. [Google Scholar] [CrossRef] [Green Version]
- Yodpitak, S.; Mahatheeranont, S.; Boonyawan, D.; Sookwong, P.; Roytrakul, S.; Norkaew, O. Cold plasma treatment to improve germination and enhance the bioactive phytochemical content of germinated brown rice. Food Chem. 2019, 289, 328–339. [Google Scholar] [CrossRef]
- Liu, Y.; Yamanaka, M.; Abe-Kanoh, N.; Liu, X.; Zhu, B.; Munemasa, S.; Nakamura, T.; Murata, Y.; Nakamura, Y. Benzyl isothiocyanate ameliorates acetaldehyde-induced cytotoxicity by enhancing aldehyde dehydrogenase activity in murine hepatoma Hepa1c1c7 cells. Food Chem. Toxicol. 2017, 108, 305–313. [Google Scholar] [CrossRef]
- Sudtasarn, G.; Homsombat, W.; Chotechuen, S.; Chamarerk, V. Quantification of tocopherols, tocotrienols and γ-oryzanol contents of local rice varieties in northeastern Thailand. J. Nutr. Sci. Vitaminol. 2019, 65, S125–S128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goufo, P.; Trindade, H. Rice antioxidants: Phenolic acids, flavonoids, anthocyanins, proanthocyanidins, tocopherols, tocotrienols, γ-oryzanol, and phytic acid. Food Sci. Nutr. 2014, 2, 75–104. [Google Scholar] [CrossRef]
- Cho, J.Y.; Lee, H.J.; Kim, G.A.; Kim, G.D.; Lee, Y.S.; Shin, S.C.; Park, K.H.; Moon, J.H. Quantitative analyses of individual γ-oryzanol (steryl ferulates) in conventional and organic brown rice (Oryza sativa L.). J. Cereal Sci. 2012, 55, 337–343. [Google Scholar] [CrossRef]
- Yu, L.; Li, G.; Li, M.; Xu, F.; Beta, T.; Bao, J. Genotypic variation in phenolic acids, vitamin E and fatty acids in whole grain rice. Food Chem. 2016, 197, 776–782. [Google Scholar] [CrossRef] [PubMed]
- Minatel, I.O.; Francisqueti, F.V.; Corrêa, C.R.; Lima, G.P. Antioxidant activity of γ-oryzanol: A complex network of interactions. Int. J. Mol. Sci. 2016, 17, 1107. [Google Scholar] [CrossRef] [Green Version]
- Averilla, J.N.; Oh, J.; Kim, J.S. Carbon monoxide partially mediates protective effect of resveratrol against UVB-Induced oxidative stress in human keratinocytes. Antioxidants 2019, 8, 432. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Yu, J.; Zhang, Y.; Li, Z.; Gong, L.; Dong, S.; Mu, R. Phosphatidylinositol 3-kinase-mediated HO-1/CO represses Fis1 levels and alleviates lipopolysaccharide-induced oxidative injury in alveolar macrophages. Exp. Ther. Med. 2018, 16, 2735–2742. [Google Scholar] [CrossRef] [Green Version]
- Keum, H.; Kim, T.W.; Kim, Y.; Seo, C.; Son, Y.; Kim, J.; Kim, D.; Jung, W.; Whang, C.; Jon, S. Bilirubin nanomedicine alleviates psoriatic skin inflammation by reducing oxidative stress and suppressing pathogenic signaling. J. Control. Release 2020, 325, 359–369. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, D.Y.; Kang, S.; Miao, W.; Kim, H.; Lee, Y.; Jon, S. Bilirubin nanoparticle preconditioning protects against hepatic ischemia-reperfusion injury. Biomaterials 2017, 133, 1–10. [Google Scholar] [CrossRef]
- Zhou, Y.; Xu, X.; Wu, J.; Xu, L.; Zhang, M.; Li, Z.; Wang, D. Allyl isothiocyanate treatment alleviates chronic obstructive pulmonary disease through the Nrf2-Notch1 signaling and upregulation of MRP1. Life Sci. 2020, 243, 117291. [Google Scholar] [CrossRef]
- Bucolo, C.; Drago, F.; Maisto, R.; Romano, G.L.; D’Agata, V.; Maugeri, G.; Giunta, S. Curcumin prevents high glucose damage in retinal pigment epithelial cells through ERK1/2-mediated activation of the Nrf2/HO-1 pathway. J. Cell. Physiol. 2019, 234, 17295–17304. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Huang, Z.L.; Li, L.; Yang, Y.; Wang, C.H.; Wang, Z.T.; Ji, L.L. Quercetin attenuates toosendanin-induced hepatotoxicity through inducing the Nrf2/GCL/GSH antioxidant signaling pathway. Acta Pharmacol. Sin. 2019, 40, 75–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, G.; Qiu, Y.; Hao, J.; Fu, Q.; Deng, X. γ-Oryzanol alleviates acetaminophen-induced liver injury: Roles of modulating AMPK/GSK3β/Nrf2 and NF-κB signaling pathways. Food Funct. 2019, 10, 6858–6872. [Google Scholar] [CrossRef]
- Ahn, J.; Son, H.J.; Seo, H.D.; Ha, T.Y.; Ahn, J.; Lee, H.; Shin, S.H.; Jung, C.H.; Jang, Y.J. γ-Oryzanol improves exercise endurance and muscle strength by upregulating PPARδ and ERRγ activity in aged mice. Mol. Nutr. Food Res. 2021, 65, 2000652. [Google Scholar] [CrossRef]
- Zhu, L.; Xiang, S.; Ou, Z.; Ma, C.; Yi, X.; Huang, L.; Ma, Y.; Liang, Z.; Yuan, Z.; Wen, L.; et al. Gamma-oryzanol prevents ethanol-induced liver injury by ameliorating oxidative stress and modulating apoptosis-related protein expression in mice. J. Funct. Foods 2019, 62, 103532. [Google Scholar] [CrossRef]
- Oka, T.; Fujimoto, M.; Nagasaka, R.; Ushio, H.; Hori, M.; Ozaki, H. Cycloartenyl ferulate, a component of rice bran oil-derived γ-oryzanol, attenuates mast cell degranulation. Phytomedicine 2010, 17, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, S.; Sowa, Y.; Hashimoto, H.; Nakagami, T.; Tsuno, T.; Sakai, T. Cycloartenyl ferulate and β-sitosteryl ferulate-steryl ferulates of γ-oryzanol-suppress intracellular reactive oxygen species in cell-based system. J. Oleo Sci. 2019, 68, 765–768. [Google Scholar] [CrossRef] [Green Version]
- Hong, G.L.; Liu, J.M.; Zhao, G.J.; Tan, J.P.; Wu, B.; Li, M.F.; Liang, G.; Qiu, Q.M.; Lu, Z.Q. Cycloartenyl ferulate inhibits paraquat-induced apoptosis in HK-2 cells with the involvement of ABCC1. J. Cell. Biochem. 2016, 117, 872–880. [Google Scholar] [CrossRef]
- Jha, A.B.; Panchal, S.S. Neuroprotection and cognitive enhancement by treatment with γ-oryzanol in sporadic Alzheimer’s disease. J. Appl. Biomed. 2017, 15, 265–281. [Google Scholar] [CrossRef]
- Sawada, K.; Rahmania, H.; Matsuki, M.; Hashimoto, H.; Ito, J.; Miyazawa, T.; Nakagawa, K. Absorption and metabolism of γ-oryzanol, a characteristic functional ingredient in rice bran. J. Nutr. Sci. Vitaminol. 2019, 65, S180–S184. [Google Scholar] [CrossRef]
- Yao, Y.; Wang, H.; Xu, F.; Zhang, Y.; Li, Z.; Ju, X.; Wang, L. Insoluble-bound polyphenols of adlay seed ameliorate H2O2-induced oxidative stress in HepG2 cells via Nrf2 signalling. Food Chem. 2020, 325, 126865. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A. Anti-hypertensive effect of cereal antioxidant ferulic acid and its mechanism of action. Front. Nutr. 2019, 6, 121. [Google Scholar] [CrossRef]
- Muthusamy, G.; Gunaseelan, S.; Prasad, N.R. Ferulic acid reverses P-glycoprotein-mediated multidrug resistance via inhibition of PI3K/Akt/NF-κB signaling pathway. J. Nutr. Biochem. 2019, 63, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Yoshida, H.; Matsuki, N.; Ono, K.; Nagasaka, R.; Ushio, H.; Guo, Y.; Hiramatsu, T.; Hosoya, T.; Murata, T.; et al. Antioxidant, free radical–scavenging, and NF-κB–inhibitory activities of phytosteryl ferulates: Structure–activity studies. J. Pharmacol. Sci. 2009, 111, 328–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, P.C.; Lai, M.H.; Hsu, K.P.; Kuo, Y.H.; Chen, J.; Tsai, M.C.; Li, C.X.; Yin, X.J.; Jeyashoke, N.; Chao, L.K.P. Identification of β-sitosterol as in vitro anti-inflammatory constituent in Moringa oleifera. J. Agric. Food Chem. 2018, 66, 10748–10759. [Google Scholar] [CrossRef]
- Sultana, S.; Alam, A.; Khan, N.; Sharma, S. Inhibition of benzoyl peroxide and ultraviolet-B radiation induced oxidative stress and tumor promotion markers by cycloartenol in murine skin. Redox Rep. 2003, 8, 105–112. [Google Scholar] [CrossRef]
- Niu, H.; Li, X.; Yang, A.; Jin, Z.; Wang, X.; Wang, Q.; Yu, C.; Wei, Z.; Dou, C. Cycloartenol exerts anti-proliferative effects on Glioma U87 cells via induction of cell cycle arrest and p38 MAPK-mediated apoptosis. J. BUON 2018, 23, 1840–1845. Available online: https://www.jbuon.com/archive/23-6-1840.pdf (accessed on 1 March 2021).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rice Sample | αT (nmol/g) (Percent Contribution to Antioxidant Capacity of Rice) | γT (nmol/g) (Percent Contribution to Antioxidant Capacity of Rice) | αT3 (nmol/g) (Percent Contribution to Antioxidant Capacity of Rice) | γT3 (nmol/g) (Percent Contribution to Antioxidant Capacity of Rice) | FA (nmol/g) (Percent Contribution to Antioxidant Capacity of Rice) | CAF (nmol/g) (Percent Contribution to Antioxidant Capacity of Rice) |
|---|---|---|---|---|---|---|
| BR | 19.73 ± 2.28 (3.01 ± 0.22%) | 3.81 ± 0.40 (0.59 ± 0.03%) | 16.02 ± 1.29 (2.91 ± 0.22%) | 18.24 ± 1.41 (3.33 ± 0.26%) | N.D b | 89.00 ± 8.07 (3.09 ± 0.16%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Nakamura, T.; Guo, Y.; Matsumoto, R.; Munemasa, S.; Murata, Y.; Nakamura, Y. Cycloartenyl Ferulate Is the Predominant Compound in Brown Rice Conferring Cytoprotective Potential against Oxidative Stress-Induced Cytotoxicity. Int. J. Mol. Sci. 2023, 24, 822. https://doi.org/10.3390/ijms24010822
Wu H, Nakamura T, Guo Y, Matsumoto R, Munemasa S, Murata Y, Nakamura Y. Cycloartenyl Ferulate Is the Predominant Compound in Brown Rice Conferring Cytoprotective Potential against Oxidative Stress-Induced Cytotoxicity. International Journal of Molecular Sciences. 2023; 24(1):822. https://doi.org/10.3390/ijms24010822
Chicago/Turabian StyleWu, Hongyan, Toshiyuki Nakamura, Yingnan Guo, Riho Matsumoto, Shintaro Munemasa, Yoshiyuki Murata, and Yoshimasa Nakamura. 2023. "Cycloartenyl Ferulate Is the Predominant Compound in Brown Rice Conferring Cytoprotective Potential against Oxidative Stress-Induced Cytotoxicity" International Journal of Molecular Sciences 24, no. 1: 822. https://doi.org/10.3390/ijms24010822
APA StyleWu, H., Nakamura, T., Guo, Y., Matsumoto, R., Munemasa, S., Murata, Y., & Nakamura, Y. (2023). Cycloartenyl Ferulate Is the Predominant Compound in Brown Rice Conferring Cytoprotective Potential against Oxidative Stress-Induced Cytotoxicity. International Journal of Molecular Sciences, 24(1), 822. https://doi.org/10.3390/ijms24010822