
S100A6 Protein—Expression and Function in Norm and Pathology


Abstract
:1. Introduction
2. Localization and Expression of S100A6
3. Intracellular S100A6 Ligands
4. Structural Aspects of S100A6—Ligand Interactions
4.1. Intracellular S100A6—Involvement in Cell Proliferation and Differentiation
4.2. Intracellular S100A6—Involvement in Cellular Stress Response
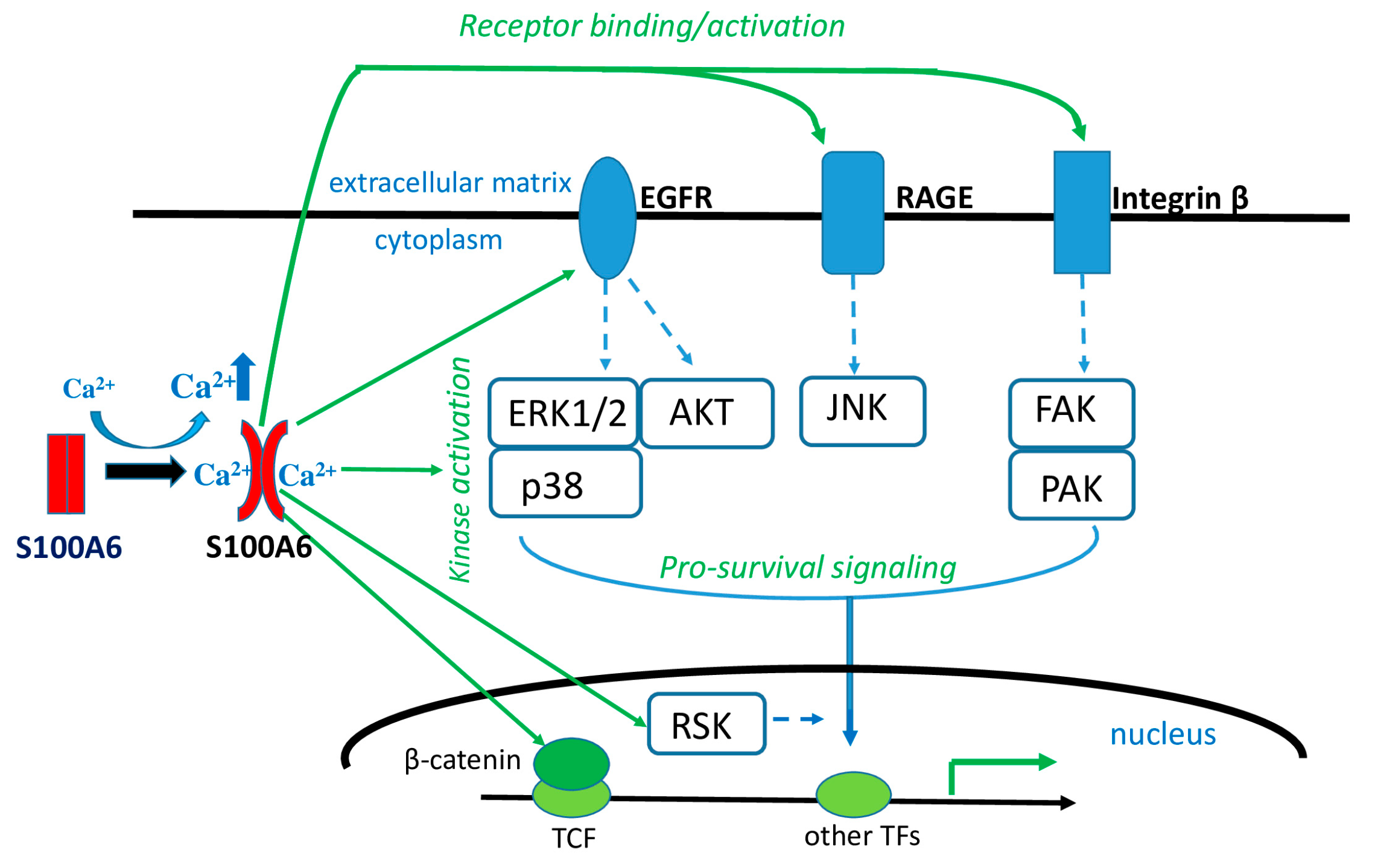
5. Extracellular S100A6—A Marker of Pathological States
6. S100A6 in Various Pathologies
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Gonzalez, L.L.; Garrie, K.; Turner, M.D. Role of S100 proteins in health and disease. Biochim. Biophys. Acta 2020, 1867, 118677. [Google Scholar] [CrossRef]
- Schäfer, B.W.; Wicki, R.; Engelkamp, D.; Mattei, M.G.; Heizmann, C.W. Isolation of a YAC clone covering a cluster of nine S100 genes on human chromosome 1q21: Rationale for a new nomenclature of the S100 calcium-binding protein family. Genomics 1995, 25, 638–643. [Google Scholar] [CrossRef] [PubMed]
- Ridinger, K.; Ilg, E.C.; Niggli, F.K.; Heizmann, C.W.; Schäfer, B.W. Clustered organization of S100 genes in human and mouse. Biochim. Biophys. Acta 1998, 1448, 254–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuźnicki, J.; Filipek, A. Purification and properties of a novel Ca2+-binding protein (10.5 kDa) from Ehrlich-ascites-tumour cells. Biochem. J. 1987, 247, 663–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuźnicki, J.; Kordowska, J.; Puzianowska, M.; Woźniewicz, B.M. Calcyclin as a marker of human epithelial cells and fibroblasts. Exp. Cell Res. 1992, 200, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Lesniak, W.; Filipek, A. Ca2+-dependent interaction of calcyclin with membrane. Biochem. Biophys. Res. Commun. 1996, 220, 269–273. [Google Scholar] [CrossRef]
- Stradal, T.B.; Gimona, M. Ca(2+)-dependent association of S100A6 (Calcyclin) with the plasma membrane and the nuclear envelope. J. Biol. Chem. 1999, 274, 31593–31596. [Google Scholar] [CrossRef] [Green Version]
- Vimalachandran, D.; Greenhalf, W.; Thompson, C.; Lüttges, J.; Prime, W.; Campbell, F.; Dodson, A.; Watson, R.; Crnogorac-Jurcevic, T.; Lemoine, N.; et al. High nuclear S100A6 (Calcyclin) is significantly associated with poor survival in pancreatic cancer patients. Cancer Res. 2005, 65, 3218–3225. [Google Scholar] [CrossRef] [Green Version]
- Allen, B.G.; Andrea, J.E.; Sutherland, C.; Schönekess, B.O.; Walsh, M.P. Molecular cloning of chicken calcyclin (S100A6) and identification of putative isoforms. Biochem. Cell Biol. 1997, 75, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Otterbein, L.R.; Kordowska, J.; Witte-Hoffmann, C.; Wang, C.L.; Dominguez, R. Crystal structures of S100A6 in the Ca(2+)-free and Ca(2+)-bound states: The calcium sensor mechanism of S100 proteins revealed at atomic resolution. Structure 2002, 10, 557–567. [Google Scholar] [CrossRef]
- Wojda, U.; Kuźnicki, J. Calcyclin from mouse Ehrlich ascites tumor cells and rabbit lung form non-covalent dimers. Biochim. Biophys. Acta 1994, 1209, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; O’Hanlon, D.; Heizmann, C.W.; Marks, A. Demonstration of heterodimer formation between S100B and S100A6 in the yeast two-hybrid system and human melanoma. Exp. Cell Res. 1999, 246, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Deloulme, J.C.; Assard, N.; Mbele, G.O.; Mangin, C.; Kuwano, R.; Baudier, J. S100A6 and S100A11 are specific targets of the calcium- and zinc-binding S100B protein in vivo. J. Biol. Chem. 2000, 275, 35302–35310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sastry, M.; Ketchem, R.R.; Crescenzi, O.; Weber, C.; Lubienski, M.J.; Hidaka, H.; Chazin, W.J. The three-dimensional structure of Ca(2+)-bound calcyclin: Implications for Ca(2+)-signal transduction by S100 proteins. Structure 1998, 6, 223–231. [Google Scholar] [CrossRef] [Green Version]
- Filipek, A.; Heizmann, C.W. Kuźnicki J. Calcyclin is a calcium and zinc binding protein. FEBS Lett. 1990, 264, 263–266. [Google Scholar] [CrossRef] [Green Version]
- Moroz, O.V.; Wilson, K.S.; Bronstein, I.B. The role of zinc in the S100 proteins: Insights from the X-ray structures. Amino Acids 2011, 41, 761–772. [Google Scholar] [CrossRef]
- Filipek, A.; Puzianowska, M.; Cieślak, B.; Kuźnicki, J. Calcyclin--Ca(2+)-binding protein homologous to glial S-100 beta is present in neurones. Neuroreport 1993, 4, 383–386. [Google Scholar] [CrossRef]
- Yamashita, N.; Ilg, E.C.; Schäfer, B.W.; Heizmann, C.W.; Kosaka, T. Distribution of a specific calcium-binding protein of the S100 protein family, S100A6 (calcyclin), in subpopulations of neurons and glial cells of the adult rat nervous system. J. Comp. Neurol. 1999, 404, 235–257. [Google Scholar] [CrossRef]
- Mandinova, A.; Atar, D.; Schäfer, B.W.; Spiess, M.; Aebi, U.; Heizmann, C.W. Distinct subcellular localization of calcium binding S100 proteins in human smooth muscle cells and their relocation in response to rises in intracellular calcium. J. Cell Sci. 1998, 111 Pt 14, 2043–2054. [Google Scholar] [CrossRef]
- Engelkamp, D.; Schäfer, B.W.; Erne, P.; Heizmann, C.W. S100 alpha, CAPL, and CACY: Molecular cloning and expression analysis of three calcium-binding proteins from human heart. Biochemistry 1992, 31, 10258–10264. [Google Scholar] [CrossRef]
- Tomida, Y.; Terasawa, M.; Kobayashi, R.; Hidaka, H. Calcyclin and calvasculin exist in human platelets. Biochem. Biophys. Res. Commun. 1992, 189, 1310–1316. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Tagliafico, E.; Manfredini, R.; Grande, A.; Rossi, E.; Zucchini, P.; Torelli, G.; Torelli, U. Abundance of the primary transcript and its processed product of growth-related genes in normal and leukemic cells during proliferation and differentiation. Cancer Res. 1992, 52, 11–16. [Google Scholar] [PubMed]
- Leśniak, W.; Filipek, A. S100A6 as a Constituent and Potential Marker of Adult and Cancer Stem Cells. Stem Cell Rev. Rep. 2022, 18, 2699–2708. [Google Scholar] [CrossRef] [PubMed]
- Orre, L.M.; Pernemalm, M.; Lengqvist, J.; Lewensohn, R.; Lehtiö, J. Up-regulation, modification, and translocation of S100A6 induced by exposure to ionizing radiation revealed by proteomics profiling. Mol. Cell. Proteom. 2007, 6, 2122–2131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Xu, X.; Khan, A.Q.; Ling, W. High Expression of S100A6 Predicts Unfavorable Prognosis of Lung Squamous Cell Cancer. Med. Sci. Monit. 2017, 23, 5011–5017. [Google Scholar] [CrossRef] [Green Version]
- Brinck, U.; Gabius, H.J.; Zeng, F.Y.; Gerke, V.; Lazarou, D.; Zografakis, C.; Tsambaos, D.; Berger, H. Differential expression of calcyclin and its accessible ligands in various types of cutaneous tumors. J. Dermatol. Sci. 1995, 10, 181–190. [Google Scholar] [CrossRef]
- Ghezzo, F.; Valpreda, S.; De Riel, J.K.; Baserga, R. Identification of serum-responsive elements in the promoter of human calcyclin, a growth-regulated gene. DNA 1989, 8, 171–177. [Google Scholar] [CrossRef]
- Tsoporis, J.N.; Izhar, S.; Parker, T. Expression of S100A6 in cardiac myocytes limits apoptosis induced by tumor necrosis factor-alpha. J. Biol. Chem. 2008, 283, 30174–30183. [Google Scholar] [CrossRef] [Green Version]
- Tonini, G.; Casalaro, A.; Cara, A.; Di Martino, D. Inducible expression of calcyclin, a gene with strong homology to S-100 protein, during neuroblastoma cell differentiation and its prevalent expression in Schwann-like cell lines. Cancer Res. 1991, 51, 1733–1737. [Google Scholar]
- Hong, E.J.; Park, S.H.; Choi, K.C.; Leung, P.C.; Jeung, E.B. Identification of estrogen-regulated genes by microarray analysis of the uterus of immature rats exposed to endocrine disrupting chemicals. Reprod. Biol. Endocrinol. 2006, 4, 49. [Google Scholar] [CrossRef] [Green Version]
- Busch, A.; Cordery, D.; Denyer, G.S.; Biden, T.J. Expression profiling of palmitate- and oleate-regulated genes provides novel insights into the effects of chronic lipid exposure on pancreatic beta-cell function. Diabetes 2002, 51, 977–987. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, P.; Liu, D.; Gao, M.; Wang, J.; Wang, X.; Liu, Y.; Zhang, X. c-Myc Upregulated by High Glucose Inhibits HaCaT Differentiation by S100A6 Transcriptional Activation. Front. Endocrinol. 2021, 12, 676403. [Google Scholar] [CrossRef] [PubMed]
- Courtois-Coutry, N.; Le Moellic, C.; Boulkroun, S.; Fay, M.; Cluzeaud, F.; Escoubet, B.; Farman, N.; Blot-Chabaud, M. Calcyclin is an early vasopressin-induced gene in the renal collecting duct. Role in the long term regulation of ion transport. J. Biol. Chem. 2002, 277, 25728–25734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucharczak, J.; Pannequin, J.; Camby, I.; Decaestecker, C.; Kiss, R.; Martinez, J. Gastrin induces over-expression of genes involved in human U373 glioblastoma cell migration. Oncogene 2001, 20, 7021–7028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Ismaeel, Q.; Neal, C.P.; Al-Mahmoodi, H.; Almutairi, Z.; Al-Shamarti, I.; Straatman, K.; Jaunbocus, N.; Irvine, A.; Issa, E.; Moreman, C.; et al. ZEB1 and IL-6/11-STAT3 signalling cooperate to define invasive potential of pancreatic cancer cells via differential regulation of the expression of S100 proteins. Br. J. Cancer 2019, 121, 65–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leśniak, W.; Jezierska, A.; Kuźnicki, J. Upstream stimulatory factor is involved in the regulation of the human calcyclin (S100A6) gene. Biochim. Biophys. Acta 2000, 1517, 73–81. [Google Scholar] [CrossRef]
- Joo, J.H.; Kim, J.W.; Lee, Y.; Yoon, S.Y.; Kim, J.H.; Paik, S.G.; Choe, I.S. Involvement of NF-kappaB in the regulation of S100A6 gene expression in human hepatoblastoma cell line HepG2. Biochem. Biophys. Res. Commun. 2003, 307, 274–280. [Google Scholar] [CrossRef]
- Króliczak, W.; Pietrzak, M.; Puzianowska-Kuznicka, M. P53-dependent suppression of the human calcyclin gene (S100A6): The role of Sp1 and of NFkappaB. Acta Biochim. Pol. 2008, 55, 559–570. [Google Scholar] [CrossRef]
- Graczyk, A.; Leśniak, W. S100A6 expression in keratinocytes and its impact on epidermal differentiation. Int. J. Biochem. Cell Biol. 2014, 57, 135–141. [Google Scholar] [CrossRef]
- Kilańczyk, E.; Graczyk, A.; Ostrowska, H.; Kasacka, I.; Leśniak, W.; Filipek, A. S100A6 is transcriptionally regulated by β-catenin and interacts with a novel target, lamin A/C, in colorectal cancer cells. Cell Calcium 2012, 51, 470–477. [Google Scholar] [CrossRef]
- Dogra, S.; Das, D.; Maity, S.K.; Paul, A.; Rawat, P.; Daniel, P.V.; Das, K.; Mitra, S.; Chakrabarti, P.; Mondal, P. Liver-Derived S100A6 Propels β-Cell Dysfunction in NAFLD. Diabetes 2022, 71, 2284–2296. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Lv, X.; Zhang, Z.; Xie, S. S100A6/miR193a regulates the proliferation, invasion, migration and angiogenesis of lung cancer cells through the P53 acetylation. Am. J. Transl. Res. 2019, 11, 4634–4649. [Google Scholar] [PubMed]
- Liu, Q.; Luo, J.; Wang, H.; Zhang, L.; Jin, G. SNHG1 functions as an oncogenic lncRNA and promotes osteosarcoma progression by up-regulating S100A6 via miR-493-5p. Acta Biochim. Biophys. Sin. 2022, 54, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Xie, K. miR-202-3p overexpression attenuates endometriosis-like lesions by modulating YAP-dependent transcription of S100A6 in murine models. Life Sci. 2020, 265, 118757. [Google Scholar] [CrossRef]
- Abdi, J.; Rashedi, I.; Keating, A. Concise Review: TLR Pathway-miRNA Interplay in Mesenchymal Stromal Cells: Regulatory Roles and Therapeutic Directions. Stem Cells 2018, 36, 1655–1662. [Google Scholar] [CrossRef] [Green Version]
- Rehman, I.; Cross, S.S.; Catto, J.W.; Leiblich, A.; Mukherjee, A.; Azzouzi, A.R.; Leung, H.Y.; Hamdy, F.C. Promoter hyper-methylation of calcium binding proteins S100A6 and S100A2 in human prostate cancer. Prostate 2005, 65, 322–330. [Google Scholar] [CrossRef]
- Leśniak, W.; Słomnicki, Ł.P.; Kuźnicki, J. Epigenetic control of the S100A6 (calcyclin) gene expression. J. Investig. Dermatol. 2007, 127, 2307–2314. [Google Scholar] [CrossRef]
- Golitsina, N.L.; Kordowska, J.; Wang, C.L.; Lehrer, S.S. Ca2+-dependent binding of calcyclin to muscle tropomyosin. Biochem. Biophys. Res. Commun. 1996, 220, 360–365. [Google Scholar] [CrossRef]
- Mani, R.S.; McCubbin, W.D.; Kay, C.M. Calcium-dependent regulation of caldesmon by an 11-kDa smooth muscle calcium-binding protein, caltropin. Biochemistry 1992, 31, 11896–11901. [Google Scholar] [CrossRef]
- Mani, R.S.; Kay, C.M. Calcium-dependent regulation of the caldesmon-heavy meromyosin interaction by caltropin. Biochemistry 1993, 32, 11217–11223. [Google Scholar] [CrossRef]
- Mani, R.S.; Kay, C.M. Influence of caltropin on the caldesmon induced polymerization of G-actin. Biochem. Biophys. Res. Commun. 1995, 213, 349–355. [Google Scholar] [CrossRef]
- Mani, R.S.; Kay, C.M. Effect of caltropin on caldesmon-actin interaction. J. Biol. Chem. 1995, 270, 6658–6663. [Google Scholar] [CrossRef] [Green Version]
- Filipek, A.; Zasada, A.; Wojda, U.; Makuch, R.; Dabrowska, R. Characterization of chicken gizzard calcyclin and examination of its interaction with caldesmon. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1996, 113, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Wills, F.L.; McCubbin, W.D.; Kay, C.M. Smooth muscle calponin-caltropin interaction: Effect on biological activity and stability of calponin. Biochemistry 1994, 33, 5562–5569. [Google Scholar] [CrossRef] [PubMed]
- Filipek, A.; Wojda, U. Chicken gizzard calcyclin—Distribution and potential target proteins. Biochem. Biophys. Res. Commun. 1996, 225, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.A.; Ecsédi, P.; Kovács, G.M.; Póti, Á.L.; Reményi, A.; Kardos, J.; Gógl, G.; Nyitray, L. High-throughput competitive fluorescence polarization assay reveals functional redundancy in the S100 protein family. FEBS J. 2020, 287, 2834–2846. [Google Scholar] [CrossRef] [Green Version]
- Jurewicz, E.; Robaszkiewicz, K.; Moraczewska, J.; Filipek, A. Binding of S100A6 to actin and the actin-tropomyosin complex. Sci. Rep. 2020, 10, 12824. [Google Scholar] [CrossRef]
- Robaszkiewicz, K.; Jurewicz, E.; Moraczewska, J.; Filipek, A. Ca2+-dependent binding of S100A6 to cofilin-1 regulates actin filament polymerization-depolymerization dynamics. Cell Calcium 2021, 99, 102457. [Google Scholar] [CrossRef]
- Jurewicz, E.; Wyroba, E.; Filipek, A. Tubulin-dependent secretion of S100A6 and cellular signaling pathways activated by S100A6-integrin β1 interaction. Cell Signal. 2018, 42, 21–29. [Google Scholar] [CrossRef]
- Shimamoto, S.; Takata, M.; Tokuda, M.; Oohira, F.; Tokumitsu, H.; Kobayashi, R. Interactions of S100A2 and S100A6 with the tetratricopeptide repeat proteins, Hsp90/Hsp70-organizing protein and kinesin light chain. J. Biol. Chem. 2008, 283, 28246–28258. [Google Scholar] [CrossRef] [Green Version]
- Sakane, K.; Nishiguchi, M.; Denda, M.; Yamagchi, F.; Magari, M.; Kanayama, N.; Morishita, R.; Tokumitsu, H. Identification and characterization of a centrosomal protein, FOR20 as a novel S100A6 target. Biochem. Biophys. Res. Commun. 2017, 491, 980–985. [Google Scholar] [CrossRef] [PubMed]
- Doi, S.; Fujioka, N.; Ohtsuka, S.; Kondo, R.; Yamamoto, M.; Denda, M.; Magari, M.; Kanayama, N.; Hatano, N.; Morishita, R.; et al. Regulation of the tubulin polymerization-promoting protein by Ca2+/S100 proteins. Cell Calcium 2021, 96, 102404. [Google Scholar] [CrossRef] [PubMed]
- Nedjadi, T.; Kitteringham, N.; Campbell, F.; Jenkins, R.E.; Park, B.K.; Navarro, P.; Ashcroft, F.; Tepikin, A.; Neoptolemos, J.P.; Costello, E. S100A6 binds to annexin 2 in pancreatic cancer cells and promotes pancreatic cancer cell motility. Br. J. Cancer 2009, 101, 1145–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimi, T.; Pfleghaar, K.; Kojima, S.; Pack, C.G.; Solovei, I.; Goldman, A.E.; Adam, S.A.; Shumaker, D.K.; Kinjo, M.; Cremer, T.; et al. The A- and B-type nuclear lamin networks: Microdomains involved in chromatin organization and transcription. Genes Dev. 2008, 22, 3409–3421. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Kondo, R.; Hozumi, H.; Doi, S.; Denda, M.; Magari, M.; Kanayama, N.; Hatano, N.; Morishita, R.; Tokumitsu, H. Identification and Biochemical Characterization of High Mobility Group Protein 20A as a Novel Ca2+/S100A6 Target. Biomolecules 2021, 11, 510. [Google Scholar] [CrossRef]
- Takata, M.; Shimamoto, S.; Yamaguchi, F.; Tokuda, M.; Tokumitsu, H.; Kobayashi, R. Regulation of nuclear localization signal-importin α interaction by Ca2+/S100A6. FEBS Lett. 2010, 584, 4517–4523. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Chaturvedi, D.; Patel, T.B. Localization and retention of p90 ribosomal S6 kinase 1 in the nucleus: Implications for its function. Mol. Biol. Cell. 2012, 23, 503–515. [Google Scholar] [CrossRef]
- Schloer, S.; Pajonczyk, D.; Rescher, U. Annexins in Translational Research: Hidden Treasures to Be Found. Int. J. Mol. Sci. 2018, 19, 1781. [Google Scholar] [CrossRef] [Green Version]
- Filipek, A.; Gerke, V.; Weber, K.; Kuźnicki, J. Characterization of the cell-cycle-regulated protein calcyclin from Ehrlich ascites tumor cells. Identification of two binding proteins obtained by Ca2+-dependent affinity chromatography. Eur. J. Biochem. 1991, 195, 795–800. [Google Scholar] [CrossRef]
- Tokumitsu, H.; Mizutani, A.; Muramatsu, M.; Yokota, T.; Arai, K.; Hidaka, H. Molecular cloning of rabbit CAP-50, a calcyclin-associated annexin protein. Biochim. Biophys. Res. Commun. 1992, 186, 1227–1235. [Google Scholar] [CrossRef]
- Zeng, F.Y.; Gerke, V.; Gabius, H.J. Identification of annexin II, annexin VI and glyceraldehyde-3-phosphate dehydrogenase as calcyclin-binding proteins in bovine heart. Int. J. Biochem. 1993, 25, 1019–1027. [Google Scholar] [CrossRef] [PubMed]
- Streicher, W.W.; Lopez, M.M.; Makhatadze, G.I. Annexin I and annexin II N-terminal peptides binding to S100 protein family members: Specificity and thermodynamic characterization. Biochemistry 2009, 48, 2788–2798. [Google Scholar] [CrossRef] [Green Version]
- Nowotny, M.; Spiechowicz, M.; Jastrzebska, B.; Filipek, A.; Kitagawa, K.; Kuznick, J. Calcium-regulated interaction of Sgt1 with S100A6 (calcyclin) and other S100 proteins. J. Biol. Chem. 2003, 278, 26923–26928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiechowicz, M.; Zylicz, A.; Bieganowski, P.; Kuznicki, J.; Filipek, A. Hsp70 is a new target of Sgt1—An interaction modulated by S100A6. Biochem. Biophys. Res. Commun. 2007, 357, 1148–1153. [Google Scholar] [CrossRef] [PubMed]
- Filipek, A.; Kuźnicki, J. Molecular cloning and expression of a mouse brain cDNA encoding a novel protein target of calcyclin. J. Neurochem. 1998, 70, 1793–1798. [Google Scholar] [CrossRef]
- Filipek, A.; Michowski, W.; Kuznicki, J. Involvement of S100A6 (calcyclin) and its binding partners in intracellular signaling pathways. Adv. Enzym. Regul. 2008, 48, 225–239. [Google Scholar] [CrossRef]
- Shimamoto, S.; Kubota, Y.; Yamaguchi, F.; Tokumitsu, H.; Kobayashi, R. Ca2+/S100 proteins act as upstream regulators of the chaperone-associated ubiquitin ligase CHIP (C terminus of Hsc70-interacting protein). J. Biol. Chem. 2013, 288, 7158–7168. [Google Scholar] [CrossRef] [Green Version]
- Shimamoto, S.; Tsuchiya, M.; Yamaguchi, F.; Kubota, Y.; Tokumitsu, H.; Kobayashi, R. Ca2+/S100 proteins inhibit the interaction of FKBP38 with Bcl-2 and Hsp90. Biochem. J. 2014, 458, 141–152. [Google Scholar] [CrossRef]
- Shimamoto, S.; Kubota, Y.; Tokumitsu, H.; Kobayashi, R. S100 proteins regulate the interaction of Hsp90 with Cyclophilin 40 and FKBP52 through their tetratricopeptide repeats. FEBS Lett. 2010, 584, 1119–1125. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, F.; Umeda, Y.; Shimamoto, S.; Tsuchiya, M.; Tokumitsu, H.; Tokuda, M.; Kobayashi, R. S100 proteins modulate protein phosphatase 5 function: A link between CA2+ signal transduction and protein dephosphorylation. J. Biol. Chem. 2012, 287, 13787–13798. [Google Scholar] [CrossRef] [Green Version]
- Kilanczyk, E.; Filipek, S.; Jastrzebska, B.; Filipek, A. CacyBP/SIP binds ERK1/2 and affects transcriptional activity of Elk-1. Biochem. Biophys. Res. Commun. 2009, 380, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Wasik, U.; Kadziolka, B.; Kilanczyk, E.; Filipek, A. Influence of S100A6 on CacyBP/SIP Phosphorylation and Elk-1 Transcriptional Activity in Neuroblastoma NB2a Cells. J. Cell. Biochem. 2016, 117, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Słomnicki, Ł.P.; Nawrot, B.; Leśniak, W. S100A6 binds p53 and affects its activity. Int. J. Biochem. Cell Biol. 2009, 41, 784–790. [Google Scholar] [CrossRef] [PubMed]
- van Dieck, J.; Brandt, T.; Teufel, D.P.; Veprintsev, D.B.; Joerger, A.C.; Fersht, A.R. Molecular basis of S100 proteins interacting with the p53 homologs p63 and p73. Oncogene 2010, 29, 2024–2035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graczyk, A.; Słomnicki, L.P.; Leśniak, W. S100A6 competes with the TAZ2 domain of p300 for binding to p53 and attenuates p53 acetylation. J. Mol. Biol. 2013, 425, 3488–3494. [Google Scholar] [CrossRef]
- van Dieck, J.; Lum, J.K.; Teufel, D.P.; Fersht, A.R. S100 proteins interact with the N-terminal domain of MDM2. FEBS Lett. 2010, 584, 3269–3274. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Fernandez, M.R.; Rutherford, T.J.; Fersht, A.R. Members of the S100 family bind p53 in two distinct ways. Protein Sci. 2008, 17, 1663–1670. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Méndez, J.D.; Méndez-Valenzuela, V.; Aguilar-Hernández, M.M. Cellular signalling of the receptor for advanced glycation end products (RAGE). Cell Signal. 2013, 25, 2185–2197. [Google Scholar] [CrossRef]
- Mohan, S.K.; Gupta, A.A.; Yu, C. Interaction of the S100A6 mutant (C3S) with the V domain of the receptor for advanced glycation end products (RAGE). Biochem. Biophys. Res. Commun. 2013, 434, 328–333. [Google Scholar] [CrossRef]
- Yatime, L.; Betzer, C.; Jensen, R.K.; Mortensen, S.; Jensen, P.H.; Andersen, G.R. The Structure of the RAGE: S100A6 Complex Reveals a Unique Mode of Homodimerization for S100 Proteins. Structure 2016, 24, 2043–2052. [Google Scholar] [CrossRef] [Green Version]
- Kazakov, A.S.; Sofin, A.D.; Avkhacheva, N.V.; Denesyuk, A.I.; Deryusheva, E.I.; Rastrygina, V.A.; Sokolov, A.S.; Permyakova, M.E.; Litus, E.A.; Uversky, V.N.; et al. Interferon Beta Activity Is Modulated via Binding of Specific S100 Proteins. Int. J. Mol. Sci. 2020, 21, 9473. [Google Scholar] [CrossRef] [PubMed]
- Kazakov, A.S.; Deryusheva, E.I.; Sokolov, A.S.; Permyakova, M.E.; Litus, E.A.; Rastrygin, V.A.; Uversky, V.N.; Permyakov, E.A.; Permyakov, S.E. Erythropoietin Interacts with Specific S100 Proteins. Biomolecules 2022, 12, 120. [Google Scholar] [CrossRef]
- Kazakov, A.; Sokolov, A.S.; Permyakova, M.E.; Litus, E.A.; Uversky, V.N.; Permyakov, E.A.; Permyakov, S.E. Specific cytokines of interleukin-6 family interact with S100 proteins. Cell Calcium 2022, 101, 102520. [Google Scholar] [CrossRef]
- Jurewicz, E.; Kasacka, I.; Bankowski, E.; Filipek, A. S100A6 and its extracellular targets in Wharton’s jelly of healthy and preeclamptic patients. Placenta 2014, 35, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Dowarha, D.; Katte, R.; Chou, R.H.; Filipek, A.; Yu, C. Lysozyme as the anti-proliferative agent to block the interaction between S100A6 and the RAGE V domain. PLoS ONE 2019, 14, e0216427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filipek, A.; Wojda, U.; Leśniak, W. Interaction of calcyclin and its cyanogen bromide fragments with annexin II and glyceraldehyde 3-phosphate dehydrogenase. Int. J. Biochem. Cell Biol. 1995, 27, 1123–1131. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Bunick, C.G.; Chazin, W.J. Target selectivity in EF-hand calcium binding proteins. Biochim. Biophys. Acta. 2004, 1742, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Sudo, T.; Hidaka, H. Characterization of the calcyclin (S100A6) binding site of annexin XI-A by site-directed mutagenesis. FEBS Lett. 1999, 444, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.T.; Dimitrova, Y.N.; Schneider, G.; Ridenour, W.B.; Bhattacharya, S.; Soss, S.E.; Caprioli, R.M.; Filipek, A.; Chazin, W.J. Structure of the S100A6 complex with a fragment from the C-terminal domain of Siah-1 interacting protein: A novel mode for S100 protein target recognition. Biochemistry 2008, 47, 10921–10932. [Google Scholar] [CrossRef] [Green Version]
- Nowotny, M.; Bhattacharya, S.; Filipek, A.; Krezel, A.M.; Chazin, W.; Kuznicki, J. Characterization of the interaction of calcyclin (S100A6) and calcyclin-binding protein. J. Biol. Chem. 2000, 275, 31178–31182. [Google Scholar] [CrossRef] [Green Version]
- Rezvanpour, A.; Shaw, G.S. Unique S100 target protein interactions. Gen. Physiol. Biophys. 2009, 28, F39–F46. [Google Scholar] [PubMed]
- Calabretta, B.; Battini, R.; Kaczmarek, L.; de Riel, J.K.; Baserga, R. Molecular cloning of the cDNA for a growth factor-inducible gene with strong homology to S-100, a calcium-binding protein. J. Biol. Chem. 1986, 261, 12628–12632. [Google Scholar] [CrossRef] [PubMed]
- Leśniak, W.; Słomnicki, Ł.P.; Filipek, A. S100A6—New facts and features. Biochem. Biophys. Res. Commun. 2009, 390, 1087–1092. [Google Scholar] [CrossRef]
- Breen, E.C.; Tang, K. Calcyclin (S100A6) regulates pulmonary fibroblast proliferation, morphology, and cytoskeletal organization in vitro. J. Cell Biochem. 2003, 88, 848–854. [Google Scholar] [CrossRef] [PubMed]
- Słomnicki, L.P.; Leśnia, W. S100A6 (calcyclin) deficiency induces senescence-like changes in cell cycle, morphology and functional characteristics of mouse NIH 3T3 fibroblasts. J. Cell Biochem. 2010, 109, 576–584. [Google Scholar] [PubMed]
- Hwang, R.; Lee, E.J.; Kim, M.H.; Li, S.Z.; Jin, Y.J.; Rhee, Y.; Kim, Y.M.; Lim, S.K. Calcyclin, a Ca2+ ion-binding protein, contributes to the anabolic effects of simvastatin on bone. J. Biol. Chem. 2004, 279, 21239–21247. [Google Scholar] [CrossRef] [Green Version]
- Ohuchida, K.; Mizumoto, K.; Ishikawa, N.; Fujii, K.; Konomi, H.; Nagai, E.; Yamaguch, K.; Tsuneyoshi, M.; Tanaka, M. The role of S100A6 in pancreatic cancer development and its clinical implication as a diagnostic marker and therapeutic target. Clin. Cancer Res. 2005, 11, 7785–7793. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Sharff, K.A.; Chen, J.; He, T.C.; Luu, H.H. S100A6 expression and function in human osteosarcoma. Clin. Orthop. Relat. Res. 2008, 466, 2060–2070. [Google Scholar] [CrossRef] [Green Version]
- Cervera, A.M.; Apostolova, N.; Crespo, F.L.; Mata, M.; McCreath, K.J. Cells silenced for SDHB expression display characteristic features of the tumor phenotype. Cancer Res. 2008, 68, 4058–4067. [Google Scholar] [CrossRef] [Green Version]
- Luu, H.H.; Zhou, L.; Haydon, R.C.; Deyrup, A.T.; Monta, A.G.; Huo, D.; Heck, R.; Heizmann, C.W.; Peabody, T.D.; Simon, M.A.; et al. Increased expression of S100A6 is associated with decreased metastasis and inhibition of cell migration and anchorage independent growth in human osteosarcoma. Cancer Lett. 2005, 229, 135–148. [Google Scholar] [CrossRef]
- Li, Z.; Tang, M.; Ling, B.; Liu, S.; Zheng, Y.; Nie, C.; Yuan, Z.; Zhou, L.; Guo, G.; Tong, A.; et al. Increased expression of S100A6 promotes cell proliferation and migration in human hepatocellular carcinoma. J. Mol. Med. 2014, 92, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, X.; Lang, H.; Zhang, S.; Luo, Y.; Zhang, J. S100 calcium-binding protein A6 promotes epithelial-mesenchymal transition through β-catenin in pancreatic cancer cell line. PLoS ONE 2015, 10, e0121319. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhang, X.; Chen, M.; Cao, Q.; Huang, D. Effect of S100A6 over-expression on β-catenin in endometriosis. J. Obstet. Gynaecol. Res. 2015, 41, 1457–1462. [Google Scholar] [CrossRef] [PubMed]
- Rijsewijk, F.; van Deemter, L.; Wagenaar, E.; Sonnenberg, A.; Nusse, R. Transfection of the int-1 mammary oncogene in cuboidal RAC mammary cell line results in morphological transformation and tumorigenicity. EMBO J. 1987, 6, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Wu, R.; Zou, Z.; Wang, H.; Ye, L.; Li, H.; Yuan, S.; Li, X.; Zha, H.; Sun, H.; et al. S100A6 stimulates proliferation and migration of colorectal carcinoma cells through activation of the MAPK pathways. Int. J. Oncol. 2014, 44, 781–790. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Shi, D.; Xu, B.; Wang, J.; Tang, Y.L.; Xiao, W.; Shen, G.; Deng, W.; Zhao, C. S100A6 promotes cell proliferation in human nasopharyngeal carcinoma via the p38/MAPK signaling pathway. Mol. Carcinog. 2017, 56, 972–984. [Google Scholar] [CrossRef]
- Li, A.; Gu, Y.; Li, X.; Sun, H.; Zha, H.; Xie, J.; Zhao, J.; Huang, M.; Chen, L.; Peng, Q.; et al. S100A6 promotes the proliferation and migration of cervical cancer cells via the PI3K/Akt signaling pathway. Oncol. Lett. 2018, 15, 5685–5693. [Google Scholar] [CrossRef] [Green Version]
- Graczyk-Jarzynka, A.; Sobiak, B.; Mlącki, M.; Wilanowski, T.; Leśniak, W. S100A6 activates EGFR and its downstream signaling in HaCaT keratinocytes. J. Cell Physiol. 2019, 234, 17561–17569. [Google Scholar] [CrossRef]
- Parsons, J.T. Focal adhesion kinase: The first ten years. J. Cell Sci. 2003, 116, 1409–1416. [Google Scholar] [CrossRef] [Green Version]
- Leclerc, E.; Fritz, G.; Weibel, M.; Heizmann, C.W.; Galichet, A. S100B and S100A6 differentially modulate cell survival by interacting with distinct RAGE (receptor for advanced glycation end products) immunoglobulin domains. J. Biol. Chem. 2007, 282, 31317–31331. [Google Scholar] [CrossRef] [Green Version]
- Lerchenmüller, C.; Heißenberg, J.; Damilano, F.; Bezzeridis, V.J.; Krämer, I.; Bochaton-Piallat, M.L.; Hirschberg, K.; Busch, M.; Katus, H.A.; Peppel, K.; et al. S100A6 Regulates Endothelial Cell Cycle Progression by Attenuating Antiproliferative Signal Transducers and Activators of Transcription 1 Signaling. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1854–1867. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wagner, E.R.; Yan, Z.; Wang, Z.; Luther, G.; Jiang, W.; Ye, J.; Wei, Q.; Wang, J.; Zhao, L.; et al. The Calcium-Binding Protein S100A6 Accelerates Human Osteosarcoma Growth by Promoting Cell Proliferation and Inhibiting Osteogenic Differentiation. Cell Physiol. Biochem. 2015, 37, 2375–2392. [Google Scholar] [CrossRef] [PubMed]
- Lewington, A.; Padanilam, B.J.; Hammerman, M.R. Induction of calcyclin after ischemic injury to rat kidney. Am. J. Physiol. 1997, 273, F380–F385. [Google Scholar] [CrossRef] [PubMed]
- Breen, E.C.; Fu, Z.; Normand, H. Calcyclin gene expression is increased by mechanical strain in fibroblasts and lung. Am. J. Respir. Cell Mol. Biol. 1999, 21, 746–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leśniak, W.; Szczepańska, A.; Kuźnicki, J. Calcyclin (S100A6) expression is stimulated by agents evoking oxidative stress via the antioxidant response element. Biochim. Biophys. Acta 2005, 1744, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasacka, I.; Piotrowska, Ż.; Filipek, A.; Majewski, M. Influence of doxazosin on biosynthesis of S100A6 and atrial natriuretic factor peptides in the heart of spontaneously hypertensive rats. Exp. Biol. Med. 2016, 241, 375–381. [Google Scholar] [CrossRef] [Green Version]
- Yamada, J.; Jinno, S. Upregulation of calcium binding protein, S100A6, in activated astrocytes is linked to glutamate toxicity. Neuroscience 2012, 226, 119–129. [Google Scholar] [CrossRef]
- Fulda, S.; Gorman, A.M.; Hori, O.; Samali, A. Cellular stress responses: Cell survival and cell death. Int. J. Cell Biol. 2010, 2010, 214074. [Google Scholar] [CrossRef] [Green Version]
- Joo, J.; Yoon, S.Y.; Kim, J.H.; Paik, S.G.; Min, S.R.; Lim, J.S.; Choe, I.S.; Choi, I.; Kim, J.W. S100A6 (calcyclin) enhances the sensitivity to apoptosis via the upregulation of caspase-3 activity in Hep3B cells. J. Cell. Biochem. 2008, 103, 1183–1197. [Google Scholar] [CrossRef]
- Lyu, X.; Li, H.Z.; Ma, X.; Li, X.T.; Gao, Y.; Ni, D.; Shen, D.L.; Gu, L.Y.; Wang, B.J.; Zhang, Y.; et al. Elevated S100A6 (Calcyclin) enhances tumorigenesis and suppresses CXCL14-induced apoptosis in clear cell renal cell carcinoma. Oncotarget 2015, 6, 6656–6669. [Google Scholar] [CrossRef] [Green Version]
- Wirths, O.; Breyhan, H.; Marcello, A.; Cotel, M.C.; Bruck, W.; Bayer, T.A. Inflammatory changes are tightly associated with neurodegeneration in the brain and spinal cord of the APP/PS1KI mouse model of Alzheimer’s disease. Neurobiol. Aging 2010, 31, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Tong, H.; Wang, L.; Zhang, K.; Shi, J.; Wu, Y.; Bao, Y.; Wang, C. S100A6 promotes inflammation and infiltration of mononuclear/macrophages via the p-P38 and p-JNK pathways in acute liver injury. Res. Sq. 2021. [CrossRef]
- Zhang, X.F.; Ma, J.X.; Wang, Y.L.; Ma, X.L. Calcyclin (S100A6) Attenuates Inflammatory Response and Mediates Apoptosis of Chondrocytes in Osteoarthritis via the PI3K/AKT Pathway. Orthop. Surg. 2021, 13, 1094–1101. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.Y.; Wang, C.Y.; Wang, T.; Li, Y.C.; Wang, Z.Y. Glial S100A6 Degrades β-amyloid Aggregation through Targeting Competition with Zinc Ions. Aging Dis. 2019, 10, 756–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagmeyer, S.; Romão, M.A.; Cristóvão, J.S.; Vilella, A.; Zoli, M.; Gomes, C.M.; Grabrucker, A.M. Distribution and Relative Abundance of S100 Proteins in the Brain of the APP23 Alzheimer’s Disease Model Mice. Front. Neurosci. 2019, 13, 640. [Google Scholar] [CrossRef]
- Cheng, C.W.; Rifai, A.; Ka, S.M.; Shui, H.A.; Lin, Y.F.; Lee, W.H.; Chen, A. Calcium-binding proteins annexin A2 and S100A6 are sensors of tubular injury and recovery in acute renal failure. Kidney Int. 2005, 68, 2694–2703. [Google Scholar] [CrossRef] [Green Version]
- Ding, H.; Yu, J.; Chang, W.; Liu, F.; He, Z. Searching for differentially expressed proteins in spinal cord injury based on the proteomics analysis. Life Sci. 2020, 242, 117235. [Google Scholar] [CrossRef]
- Ito, M.; Kizawa, K. Expression of calcium-binding S100 proteins A4 and A6 in regions of the epithelial sac associated with the onset of hair follicle regeneration. J. Investig. Dermatol. 2001, 116, 956–963. [Google Scholar] [CrossRef] [Green Version]
- Grahn, T.H.M.; Niroula, A.; Végvári, Á.; Oburoglu, L.; Pertesi, M.; Warsi, S.; Safi, F.; Miharada, N.; Garcia, S.C.; Siva, K.; et al. S100A6 is a critical regulator of hematopoietic stem cells. Leukemia 2020, 34, 3323–3337. [Google Scholar] [CrossRef]
- Bartkowska, K.; Swiatek, I.; Aniszewska, A.; Jurewicz, E.; Turlejski, K.; Filipek, A.; Djavadian, R.L. Stress-Dependent Changes in the CacyBP/SIP Interacting Protein S100A6 in the Mouse Brain. PLoS ONE 2017, 12, e0169760. [Google Scholar] [CrossRef] [Green Version]
- Fang, B.; Liang, M.; Yang, G.; Ye, Y.; Xu, H.; He, X.; Huang, J.H. Expression of S100A6 in Rat Hippocampus after Traumatic Brain Injury Due to Lateral Head Acceleration. Int. J. Mol. Sci. 2014, 15, 6378–6390. [Google Scholar] [CrossRef] [PubMed]
- Thordarson, G.; Southard, J.N.; Talamantes, F. Purification and characterization of mouse decidual calcyclin: A novel stimulator of mouse placental lactogen-II secretion. Endocrinology 1991, 129, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Jurewicz, E.; Góral, A.; Filipek, A. S100A6 is secreted from Wharton’s jelly mesenchymal stem cells and interacts with integrin β1. Int. J. Biochem. Cell Biol. 2014, 55, 298–303. [Google Scholar] [CrossRef]
- Stewart, V.D.; Cadieux, J.; Thulasiram, M.R.; Douglas, T.C.; Drewnik, D.A.; Selamat, S.; Lao, Y.; Spicer, V.; Hannila, S.S. Myelin-associated glycoprotein alters the neuronal secretome and stimulates the release of TGFβ and proteins that affect neural plasticity. FEBS Lett. 2022, 596, 2952–2973. [Google Scholar] [CrossRef]
- Celis, J.E.; Gesser, B.; Rasmussen, H.H.; Madsen, P.; Leffers, H.; Dejgaard, K.; Honore, B.; Olsen, E.; Ratz, G.; Lauridsen, J.B.; et al. Comprehensive two-dimensional gel protein databases offer a global approach to the analysis of human cells: The transformed amnion cells (AMA) master database and its link to genome DNA sequence data. Electrophoresis 1990, 11, 989–1071. [Google Scholar] [CrossRef] [PubMed]
- Ohuchida, K.; Mizumoto, K.; Yu, J.; Yamaguchi, H.; Konomi, H.; Nagai, E.; Yamaguchi, K.; Tsuneyoshi, M.; Tanaka, M. S100A6 is increased in a stepwise manner during pancreatic carcinogenesis: Clinical value of expression analysis in 98 pancreatic juice samples. Cancer Epidemiol. Biomark. Prev. 2007, 16, 649–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnier, J.L.; Fall, N.; Thornton, S.; Witte, D.; Bennett, M.R.; Appenzeller, S.; Klein-Gitelman, M.S.; Grom, A.A.; Brunner, H.I. Urine S100 proteins as potential biomarkers of lupus nephritis activity. Arthritis Res. Ther. 2017, 19, 242. [Google Scholar] [CrossRef] [Green Version]
- Dubois, J.; Rueger, J.; Haubold, B.; Far, R.K.; Sczakiel, G. Transcriptome analyses of urine RNA reveal tumor markers for human bladder cancer: Validated amplicons for RT-qPCR-based detection. Oncotarget 2021, 12, 1011–1023. [Google Scholar] [CrossRef]
- Recalde, J.I.; Duran, J.A.; Rodriguez-Agirretxe, I.; Soria, J.; Sanchez-Tena, M.A.; Pereiro, X.; Suarez, T.; Acera, A. Changes in tear biomarker levels in keratoconus after corneal collagen crosslinking. Mol. Vis. 2019, 25, 12–21. [Google Scholar]
- Assenhöj, M.; Ward, L.J.; Ghafouri, B.; Graff, P.; Ljunggren, S.A. Metal exposure from additive manufacturing and its effect on the nasal lavage fluid proteome—A pilot study. PLoS ONE 2021, 16, e0256746. [Google Scholar] [CrossRef]
- Biji, A.; Khatun, O.; Swaraj, S.; Narayan, R.; Rajmani, R.S.; Sardar, R.; Satish, D.; Mehta, S.; Bindhu, H.; Jeevan, M.; et al. Identification of COVID-19 prognostic markers and therapeutic targets through meta-analysis and validation of Omics data from nasopharyngeal samples. EBioMedicine 2021, 70, 103525. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, K.; Niki, I.; Iino, S.; Kobayashi, S.; Hidaka, H. A role of calcyclin, a Ca(2+)-binding protein, on the Ca(2+)-dependent insulin release from the pancreatic beta cell. J. Biol. Chem. 1994, 269, 6149–6152. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Kuzumaki, N.; Ogoma, Y.; Kondo, Y. Effects of calcium-binding proteins on histamine release from permeabilized rat peritoneal mast cells. Biol. Pharm. Bull. 1994, 17, 581–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, B.R.; Hoover, S.B.; Ross, M.M.; Zhou, W.; Meani, F.; Edwards, J.B.; Spehalski, E.I.; Risinger, J.I.; Alvord, W.G.; Quiñones, O.A.; et al. Serum S100A6 concentration predicts peritoneal tumor burden in mice with epithelial ovarian cancer and is associated with advanced stage in patients. PLoS ONE 2009, 4, e7670. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, K.; Jiang, X.; Zhang, J. S100A6 as a potential serum prognostic biomarker and therapeutic target in gastric cancer. Dig. Dis. Sci. 2014, 59, 2136–2144. [Google Scholar] [CrossRef]
- Nishi, M.; Matsumoto, K.; Kobayashi, M.; Yanagita, K.; Matsumoto, T.; Nagashio, R.; Ishii, D.; Fujita, T.; Sato, Y.; Iwamura, M. Serum expression of S100A6 is a potential detection marker in patients with urothelial carcinoma in the urinary bladder. Biomed. Res. 2014, 35, 351–356. [Google Scholar] [CrossRef] [Green Version]
- Ismail, M.F.; El Boghdady, N.A.; Shabayek, M.I.; Awida, H.A.; Abozeed, H. Evaluation and screening of mRNA S100A genes as serological biomarkers in different stages of bladder cancer in Egypt. Tumour Biol. 2016, 37, 4621–4631. [Google Scholar] [CrossRef]
- Wang, T.; Liang, Y.; Thakur, A.; Zhang, S.; Yang, T.; Chen, T.; Gao, L.; Chen, M.; Ren, H. Diagnostic significance of S100A2 and S100A6 levels in sera of patients with non-small cell lung cancer. Tumour Biol. 2016, 37, 2299–2304. [Google Scholar] [CrossRef]
- Bai, Y.; Li, L.D.; Li, J.; Lu, X. Prognostic values of S100 family members in ovarian cancer patients. BMC Cancer 2018, 18, 1256. [Google Scholar] [CrossRef]
- Cai, X.Y.; Lu, L.; Wang, Y.N.; Jin, C.; Zhang, R.Y.; Zhang, Q.; Chen, Q.J.; Shen, W.F. Association of increased S100B, S100A6 and S100P in serum levels with acute coronary syndrome and also with the severity of myocardial infarction in cardiac tissue of rat models with ischemia-reperfusion injury. Atherosclerosis 2011, 217, 536–542. [Google Scholar] [CrossRef]
- Dong, X.H.; Dai, D.; Yang, Z.D.; Yu, X.O.; Li, H.; Kang, H. S100 calcium binding protein A6 and associated long noncoding ribonucleic acids as biomarkers in the diagnosis and staging of primary biliary cholangitis. World J. Gastroenterol. 2021, 27, 1973–1992. [Google Scholar] [CrossRef]
- Güzel, C.; van den Berg, C.B.; Duvekot, J.J.; Stingl, C.; van den Bosch, T.P.P.; van der Weiden, M.; Steegers, E.A.P.; Steegers-Theunisse, R.P.M.; Luider, T.M. Quantification of Calcyclin and Heat Shock Protein 90 in Sera from Women with and without Preeclampsia by Mass Spectrometry. Proteom. Clin. Appl. 2019, 13, e1800181. [Google Scholar] [CrossRef] [PubMed]
- Landi, C.; Bargagli, E.; Bianchi, L.; Gagliardi, A.; Carleo, A.; Bennett, D.; Perari, M.G.; Armini, A.; Prasse, A.; Rottoli, P.; et al. Towards a functional proteomics approach to the comprehension of idiopathic pulmonary fibrosis, sarcoidosis, systemic sclerosis and pulmonary Langerhans cell histiocytosis. J. Proteom. 2013, 83, 60–75. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Chen, P.W.; Tsai, P.J.; Su, S.H.; Liao, P.C. Proteomics analysis revealed changes in rat bronchoalveolar lavage fluid proteins associated with oil mist exposure. Proteomics 2006, 6, 2236–2250. [Google Scholar] [CrossRef] [PubMed]
- Benitez-Del-Castillo, J.M.; Soria, J.; Acera, A.; Muñoz, A.M.; Rodríguez, S.; Suárez, T. Quantification of a panel for dry-eye protein biomarkers in tears: A comparative pilot study using standard ELISA and customized microarrays. Mol. Vis. 2021, 27, 243–261. [Google Scholar] [PubMed]
- Valencia, E.; García, M.; Fernández-Vega, B.; Pereiro, R.; Lobo, L.; González-Iglesias, H. Targeted Analysis of Tears Revealed Specific Altered Metal Homeostasis in Age-Related Macular Degeneration. Investig. Ophthalmol. Vis. Sci. 2022, 63, 10. [Google Scholar] [CrossRef]
- Klein, L.L.; Freitag, B.C.; Gibbs, R.S.; Reddy, A.P.; Nagalla, S.R.; Gravett, M.G. Detection of intra-amniotic infection in a rabbit model by proteomics-based amniotic fluid analysis. Am. J. Obstet. Gynecol. 2005, 193, 1302–1306. [Google Scholar] [CrossRef]
- Patgaonkar, M.; Aranha, C.; Bhonde, G.; Reddy, K.V. Identification and characterization of anti-microbial peptides from rabbit vaginal fluid. Vet. Immunol. Immunopathol. 2011, 139, 176–186. [Google Scholar] [CrossRef]
- Zhang, S.P.; Wu, Z.Z.; Wu, Y.W.; Su, S.B.; Tong, J. Mechanism study of adaptive response in high background radiation area of Yangjiang in China. Zhonghua Yu Fang Yi Xue Za Zhi 2010, 44, 815–819. (In Chinese) [Google Scholar]
- Henderson, N.C.; Rieder, F.; Wynn, T.A. Fibrosis: From mechanisms to medicines. Nature 2020, 587, 555–566. [Google Scholar] [CrossRef]
- Krenkel, O.; Hundertmark, J.; Ritz, T.P.; Weiskirchen, R.; Tacke, F. Single Cell RNA Sequencing Identifies Subsets of Hepatic Stellate Cells and Myofibroblasts in Liver Fibrosis. Cells 2019, 8, 503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landi, C.; Bargagli, E.; Carleo, A.; Refini, R.M.; Bennett, D.; Bianchi, L.; Cillis, G.; Prasse, A.; Bini, L.; Rottoli, P. Bronchoalveolar lavage proteomic analysis in pulmonary fibrosis associated with systemic sclerosis: S100A6 and 14-3-3ε as potential biomarkers. Rheumatology 2019, 58, 165–178. [Google Scholar] [PubMed]
- Ju, W.; Eichinger, F.; Bitzer, M.; Oh, J.; McWeeney, S.; Berthier, C.C.; Shedden, K.; Cohen, C.D.; Henger, A.; Krick, S.; et al. Renal gene and protein expression signatures for prediction of kidney disease progression. Am. J. Pathol. 2009, 174, 2073–2085. [Google Scholar] [CrossRef]
- Stanton, L.W.; Garrard, L.J.; Damm, D.; Garrick, B.L.; Lam, A.; Kapoun, A.M.; Zheng, Q.; Protter, A.A.; Schreiner, G.F.; White, R.T. Altered patterns of gene expression in response to myocardial infarction. Circ. Res. 2000, 86, 939–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, P.; He, H.; Kristine, M.S.; Guan, W.; Gao, J.; Wang, Z.; Hu, J.; Han, L.; Li, J.; Han, W.; et al. Therapeutic effects of recombinant human S100A6 and soluble receptor for advanced glycation end products(sRAGE) on CCl4-induced liver fibrosis in mice. Eur. J. Pharmacol. 2018, 833, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Boom, A.; Pochet, R.; Authelet, M.; Pradier, L.; Borghgraef, P.; Van Leuven, F.; Heizmann, C.W.; Brion, J.P. Astrocytic calcium/zinc binding protein S100A6 over expression in Alzheimer’s disease and in PS1/APP transgenic mice models. Biochim. Biophys. Acta 2004, 1742, 161–168. [Google Scholar] [CrossRef]
- Astillero-Lopez, V.; Gonzalez-Rodriguez, M.; Villar-Conde, S.; Flores-Cuadrado, A.; Martinez-Marcos, A.; Ubeda-Banon, I.; Saiz-Sanchez, D. Neurodegeneration and astrogliosis in the entorhinal cortex in Alzheimer’s disease: Stereological layer-specific assessment and proteomic analysis. Alzheimers Dement. 2022, 18, 2468–2480. [Google Scholar] [CrossRef]
- Hoyaux, D.; Alao, J.; Fuchs, J.; Kiss, R.; Keller, B.; Heizmann, C.W.; Pochet, R.; Frermann, D. S100A6, a calcium- and zinc-binding protein, is overexpressed in SOD1 mutant mice, a model for amyotrophic lateral sclerosis. Biochim. Biophys. Acta 2000, 1498, 264–272. [Google Scholar] [CrossRef] [Green Version]
- Hoyaux, D.; Boom, A.; Van Den Bosch, L.; Belot, N.; Martin, J.-J.; Heizmann, C.W.; Kiss, R.; Pochet, R. S100A6 Overexpression within Astrocytes Associated with Impaired Axons from Both ALS Mouse Model and Human Patients. J. Neuropath. Exp. Neurol. 2002, 61, 736–744. [Google Scholar] [CrossRef] [Green Version]
- Iridoy, M.O.; Zubiri, I.; Zelaya, M.V.; Martinez, L.; Ausín, K.; Lachen-Montes, M.; Santamaría, E.; Fernandez-Irigoyen, J.; Jericó, I. Neuroanatomical Quantitative Proteomics Reveals Common Pathogenic Biological Routes between Amyotrophic Lateral Sclerosis (ALS) and Frontotemporal Dementia (FTD). Int. J. Mol. Sci. 2019, 20, 4. [Google Scholar] [CrossRef] [Green Version]
- Filipek, A.; Leśniak, W. S100A6 and Its Brain Ligands in Neurodegenerative Disorders. Int. J. Mol. Sci. 2020, 21, 3979. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, L.; Li, Z.; Song, Y.; Yuan, Y.; Liu, T.; Hong, J.; Wang, Q.; Chang, H.; Kuang, Z.; et al. Potential Serum Biomarkers for Postoperative Neurocognitive Disorders Based on Proteomic Analysis of Cognitive-Related Brain Regions. Front. Aging Neurosci. 2021, 13, 741263. [Google Scholar] [CrossRef] [PubMed]
- Jurewicz, E.; Filipek, A. Ca2+- binding proteins of the S100 family in preeclampsia. Placenta 2022, 127, 43–51. [Google Scholar] [CrossRef]
- Peng, Y.; Lin, J.; Ma, J.; Lin, K.; Xu, K.; Lin, J. Upregulation of S100A6 in patients with endometriosis and its role in ectopic endometrial stromal cells. Gynecol. Endocrinol. 2018, 34, 815–820. [Google Scholar] [CrossRef] [PubMed]
- Allgöwer, C.; Kretz, A.L.; von Karstedt, S.; Wittau, M.; Henne-Bruns, D.; Lemke, J. Friend or Foe: S100 Proteins in Cancer. Cancers 2020, 12, 2037. [Google Scholar] [CrossRef]
- Weterman, M.A.; Stoopen, G.M.; van Muijen, G.N.; Kuznicki, J.; Ruiter, D.J.; Bloemers, H.P. Expression of calcyclin in human melanoma cell lines correlates with metastatic behavior in nude mice. Cancer Res. 1992, 52, 1291–1296. [Google Scholar]
- Komatsu, K.; Andoh, A.; Ishiguro, S.; Suzuki, N.; Hunai, H.; Kobune-Fujiwara, Y.; Kameyama, M.; Miyoshi, J.; Akedo, H.; Nakamura, H. Increased expression of S100A6 (Calcyclin), a calcium-binding protein of the S100 family, in human colorectal adenocarcinomas. Clin. Cancer Res. 2000, 6, 172–177. [Google Scholar]
- Yang, Y.Q.; Zhang, L.J.; Dong, H.; Jiang, C.L.; Zhu, Z.G.; Wu, J.X.; Wu, Y.L.; Han, J.S.; Xiao, H.S.; Gao, H.J.; et al. Upregulated expression of S100A6 in human gastric cancer. J. Dig. Dis. 2007, 8, 186–193. [Google Scholar] [CrossRef]
- Kim, J.; Kim, J.; Yoon, S.; Joo, J.; Lee, Y.; Lee, K.; Chung, J.; Choe, I. S100A6 protein as a marker for differential diagnosis of cholangiocarcinoma from hepatocellular carcinoma. Hepatol. Res. 2002, 23, 274. [Google Scholar] [CrossRef]
- Onsurathum, S.; Haonon, O.; Pinlaor, P.; Pairojkul, C.; Khuntikeo, N.; Thanan, R.; Roytrakul, S.; Pinlaor, S. Proteomics detection of S100A6 in tumor tissue interstitial fluid and evaluation of its potential as a biomarker of cholangiocarcinoma. Tumour Biol. 2018, 40, 1010428318767195. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Yao, R.; Chen, J.; Zou, Q.; Zeng, L. S100 family members: Potential therapeutic target in patients with hepatocellular carcinoma: A STROBE study. Medicine 2021, 100, e24135. [Google Scholar] [CrossRef] [PubMed]
- Melle, C.; Ernst, G.; Schimmel, B.; Bleul, A.; von Eggeling, F. Colon-derived liver metastasis, colorectal carcinoma, and hepatocellular carcinoma can be discriminated by the Ca(2+)-binding proteins S100A6 and S100A11. PLoS ONE 2008, 3, e3767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterova, E.; Bures, J.; Moravkova, P.; Kohoutova, D. Tissue mRNA for S100A4, S100A6, S100A8, S100A9, S100A11 and S100P Proteins in Colorectal Neoplasia: A Pilot Study. Molecules 2021, 26, 402. [Google Scholar] [CrossRef] [PubMed]
- Jia, R.; Sui, Z.; Zhang, H.; Yu, Z. Identification and Validation of Immune-Related Gene Signature for Predicting Lymph Node Metastasis and Prognosis in Lung Adenocarcinoma. Front. Mol. Biosci. 2021, 8, 679031. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, X.; Xie, J.; Zhu, Q.; Yuan, Y. A Novel Immune-Related Prognostic Signature Predicting Survival in Patients with Pancreatic Adenocarcinoma. J. Oncol. 2022, 2022, 8909631. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Ligand | Target Region | Direct Effect | Proposed Function | Reference |
|---|---|---|---|---|
| Actin and tropomyosin | ND | ND | Regulation of actin filament dynamics | [48,57] |
| Caldesmon | C-terminus | Decrease in caldesmon affinity to actin and heavy meromyosin (HMM) | Regulation of caldesmon function in smooth muscle contraction | [49,50,51,52,53] |
| Calponin | ND | Stabilization of calponin | Regulation of calponin function in smooth muscle contraction | [54,55] |
| Coffilin | ND | Decrease in actin depolymerization | Regulation of actin polymerization | [58] |
| Non-muscle myosin IIA | ND | ND | Modulation of cell motility | [56] |
| Tubulin α and β | ND | ND | Regulation of microtubule organization | [59] |
| Kinesin light chain (KLC) | TPR domain | dissociation of the KLC-JIP1 complex | Modulation of KLC–cargo interaction | [60] |
| FOR20, FOP and OFD1 | N-terminus (aa 1–30) | ND | Modulation of cilia formation | [61] |
| TPPP (tubulin polymerization-promoting protein) | C-terminal region (aa 110–160) | Inhibition of TPPP dimerization | Modulation of microtubule organization | [62] |
| Lamin B1 and A/C | ND | ND | Regulation of chromatin organization | [40,63] |
| Importin α | Armadillo motif; NLS-cargo-binding domain | Inhibition of importin–NPM1 binding | Inhibition of nuclear transport | [66] |
| HMG20A | C-terminus (aa 311–347) | ND | Regulation of neuronal differentiation | [65] |
| RSK1 | ND | ND | Regulation of cell survival and proliferation | [56] |
| Annexins: Anx1, Anx2, Anx6 and Anx11 | N-terminus | ND | Modulation of membrane dynamics | [70,71,72,96] |
| Sgt1 | SGS domain | Inhibition of Sgt1-Hsp90 binding | Modulation of chaperone complexes | [74] |
| CacyBP/SIP | SGS domain | Inhibition of CacyBP/SIP phosphorylation by CKII | Regulation of CacyBP/SIP phosphatase activity and ERK1/2-Elk-1 signaling pathway | [81,82,100] |
| Hop and TOM70 | TPR domain | Dissociation of Hop and TOM70 complexes with Hsp90 or Hsp70 | Modulation of chaperone complexes | [60] |
| Melusin | C-terminus | ND | Regulation of cardioprotective pathway | [76] |
| CHIP | TPR and U-box domains | Moderate inhibition of CHIP interaction with Hsp90 and Hsp70; suppression of mutant p53 degradation | Modulation of chaperone complexes and p53 degradation | [77] |
| Cyp-40 and FKBP52 | TPR domain | Inhibition of Cyp-40 and FKBP52 binding with Hsp90 | Modulation of chaperone complexes | [79] |
| FKBP38 | TPR domain | Dissociation of the FKBP38-Hsp90 and FKBP38-Bcl2 complexes; suppression of Bcl2 stability | Modulation of chaperone complexes | [78] |
| PP5 | TPR domain | Dissociation of the PP5-Hsp90 complex; stimulation of PP5 activity | Modulation of chaperone complexes | [80] |
| P53, p63 and p73 | P53: C-terminal tetramerization domain (aa 293–393); N-terminal transactivation domain (aa 1–57) | Inhibition of p53 tetramerization | Regulation of p53, p63 and p73 oligomerization and activity | [83,84,87] |
| Mdm2 | N-terminus (aa 2–125) | ND | Moderate inhibition of p53 ubiquitination | [86] |
| RAGE | V, C1 and C2 domains | Induction of RAGE dimerization/clustering | Modulation of signal transduction through RAGE | [89,90,120] |
| Integrin β1 | Extracellular domain | Increase in FAK and PAK phosphorylation | Modulation of integrin- dependent signaling | [59] |
| NCX1 and TRPM4 | ND | ND | Modulation of ion transport | [56] |
| Erythropoietin | N- and C- terminus | ND | Regulation of erythropoietin secretion and/or activity | [92] |
| Cytokines, e.g., IFN-β, IL-11 and CNTF | ND | No effect on IFN-β-induced cytotoxicity | Modulation of cytokine activity/signaling | [91,93] |
| Lysozyme | N-terminus | Inhibition of S100A6-RAGE interaction | Modulation of RAGE-dependent signaling | [55,95] |
| Lumican, PRELP and IGFBP-1 | ND | Competition of the IGF-1–IGFBP-1 interaction | Remodeling of extracellular matrix | [94] |
| GAPDH | ND | ND | Regulation of metabolic processes | [96] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leśniak, W.; Filipek, A. S100A6 Protein—Expression and Function in Norm and Pathology. Int. J. Mol. Sci. 2023, 24, 1341. https://doi.org/10.3390/ijms24021341
Leśniak W, Filipek A. S100A6 Protein—Expression and Function in Norm and Pathology. International Journal of Molecular Sciences. 2023; 24(2):1341. https://doi.org/10.3390/ijms24021341
Chicago/Turabian StyleLeśniak, Wiesława, and Anna Filipek. 2023. "S100A6 Protein—Expression and Function in Norm and Pathology" International Journal of Molecular Sciences 24, no. 2: 1341. https://doi.org/10.3390/ijms24021341
APA StyleLeśniak, W., & Filipek, A. (2023). S100A6 Protein—Expression and Function in Norm and Pathology. International Journal of Molecular Sciences, 24(2), 1341. https://doi.org/10.3390/ijms24021341