
Involvement of Dynamic Adjustment of ABA, Proline and Sugar Levels in Rhizomes in Effective Acclimation of Solidago gigantea to Contrasting Weather and Soil Conditions in the Country of Invasion




Abstract
:1. Introduction
- (1)
- Alterations in soluble sugars, proline, and abscisic acid contents in subsequent seasons of the year are associated with the S. gigantea response to changing environmental conditions typical for the vegetation period irrespective of plant genetic distance;
- (2)
- The analyzed compounds reach their highest levels in late autumn, when the temperature drops, allowing plants to adapt to frost occurring in winter conditions;
- (3)
- Soil conditions (mostly shallowness) affect the aforementioned responses.
2. Results
2.1. Soil Conditions in Individual Locations
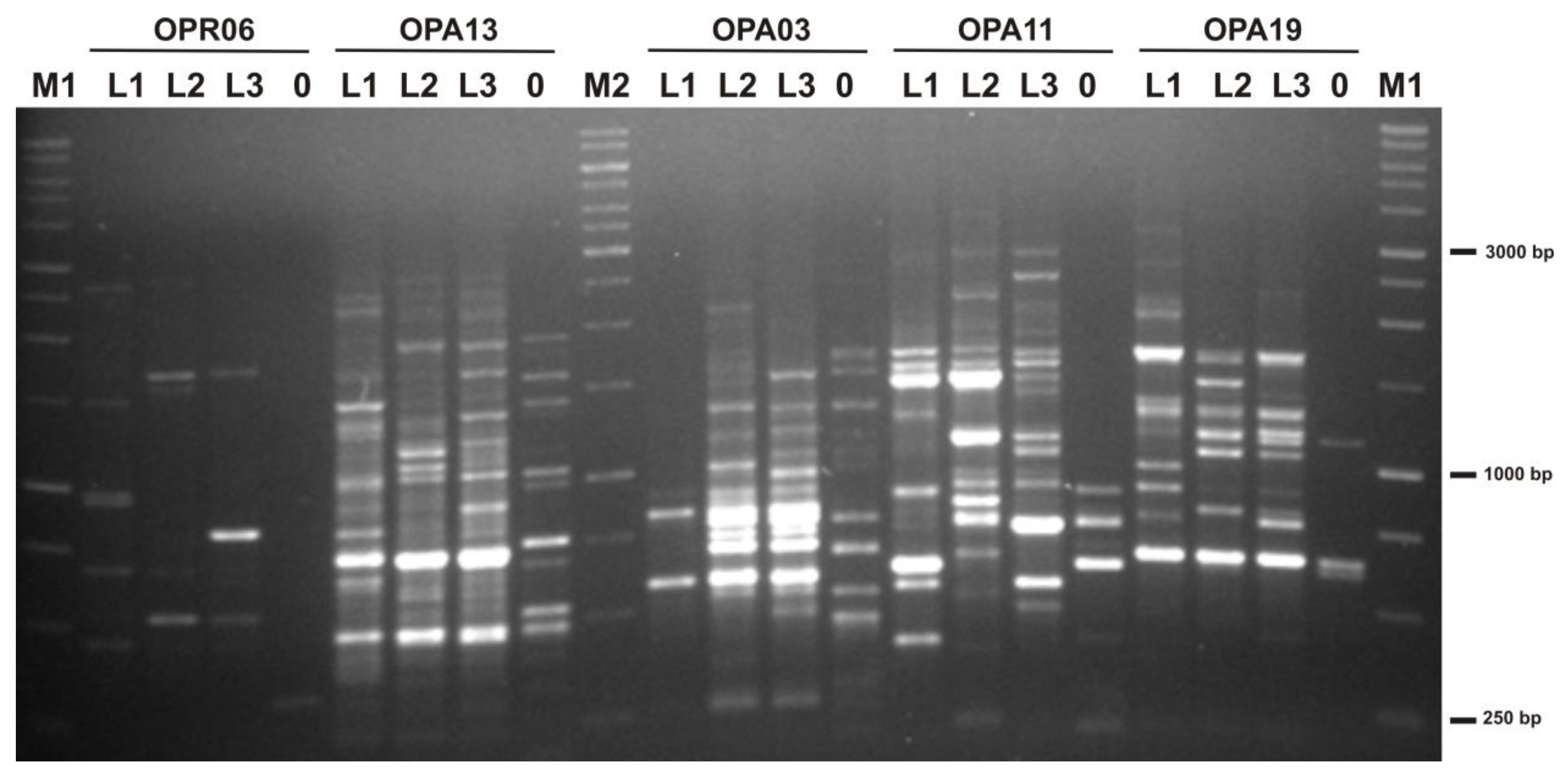
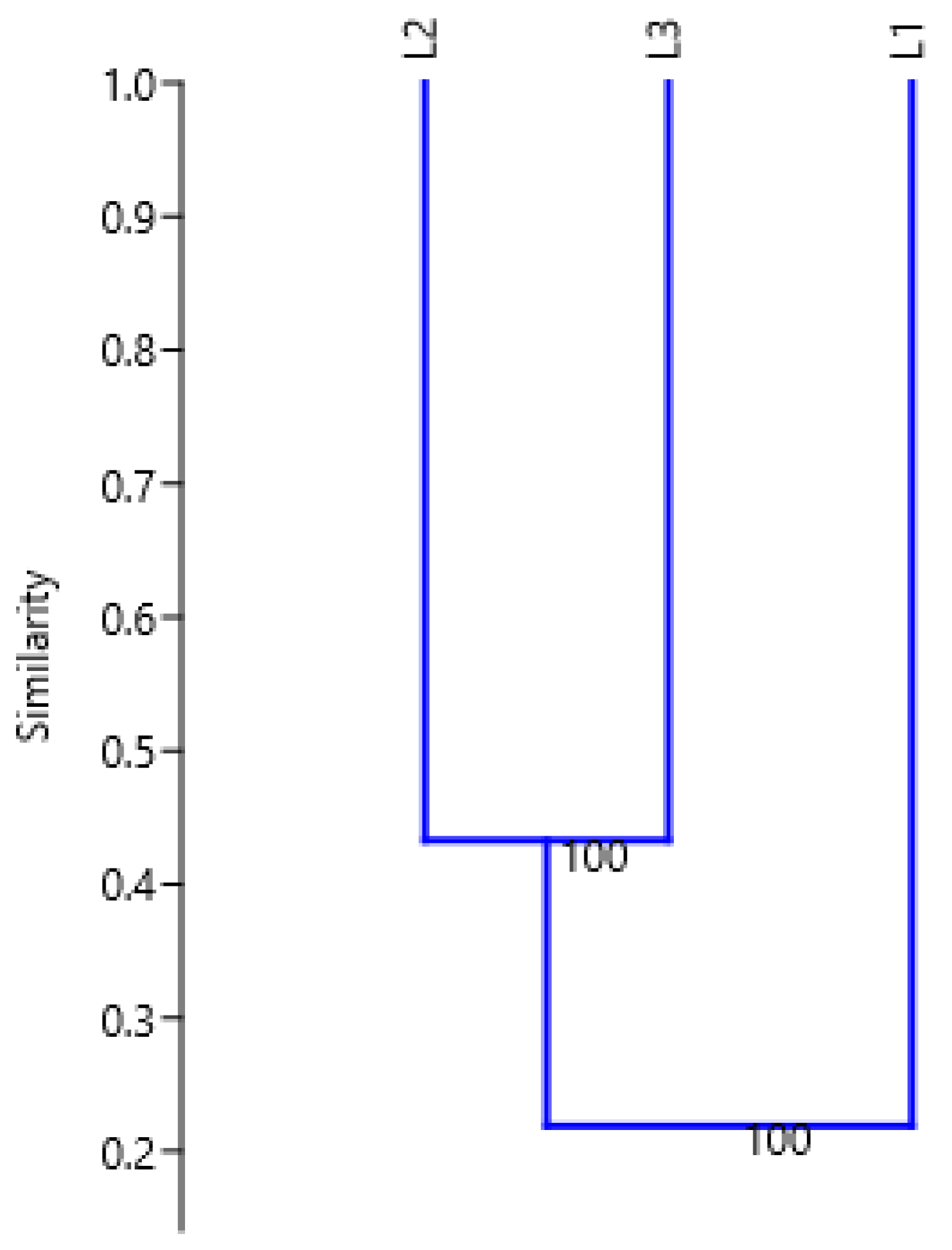
2.2. Genetic Variation between Plants from Three Locations
2.3. In Vivo Measurements in Individual Stands
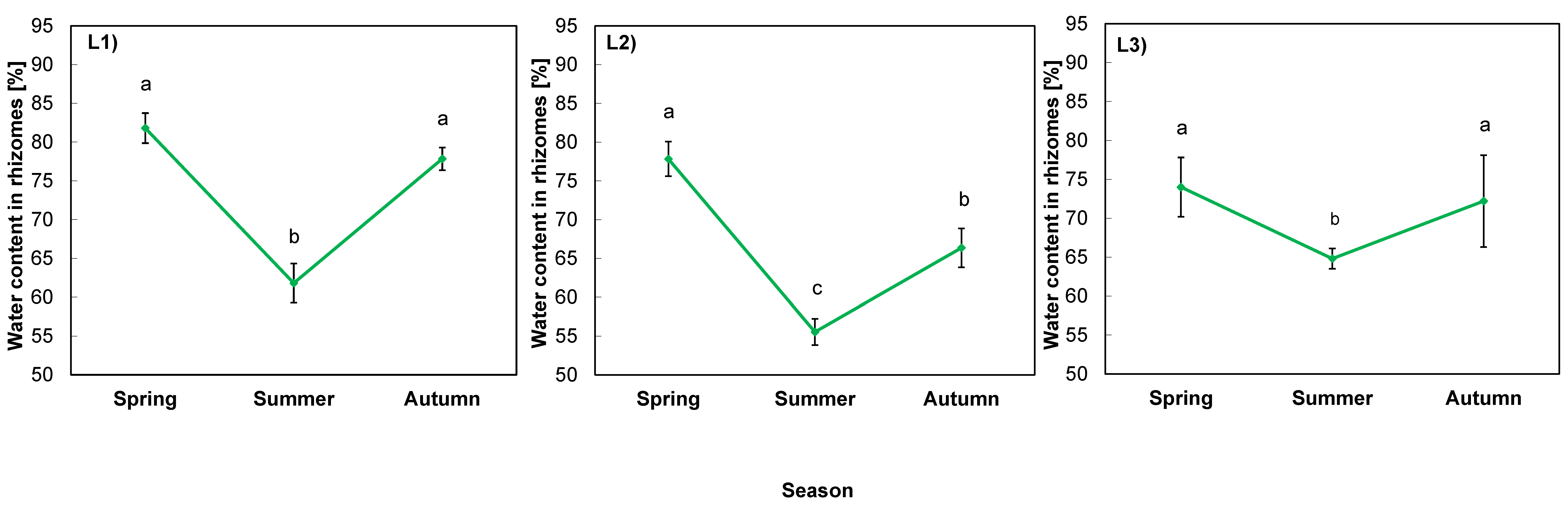
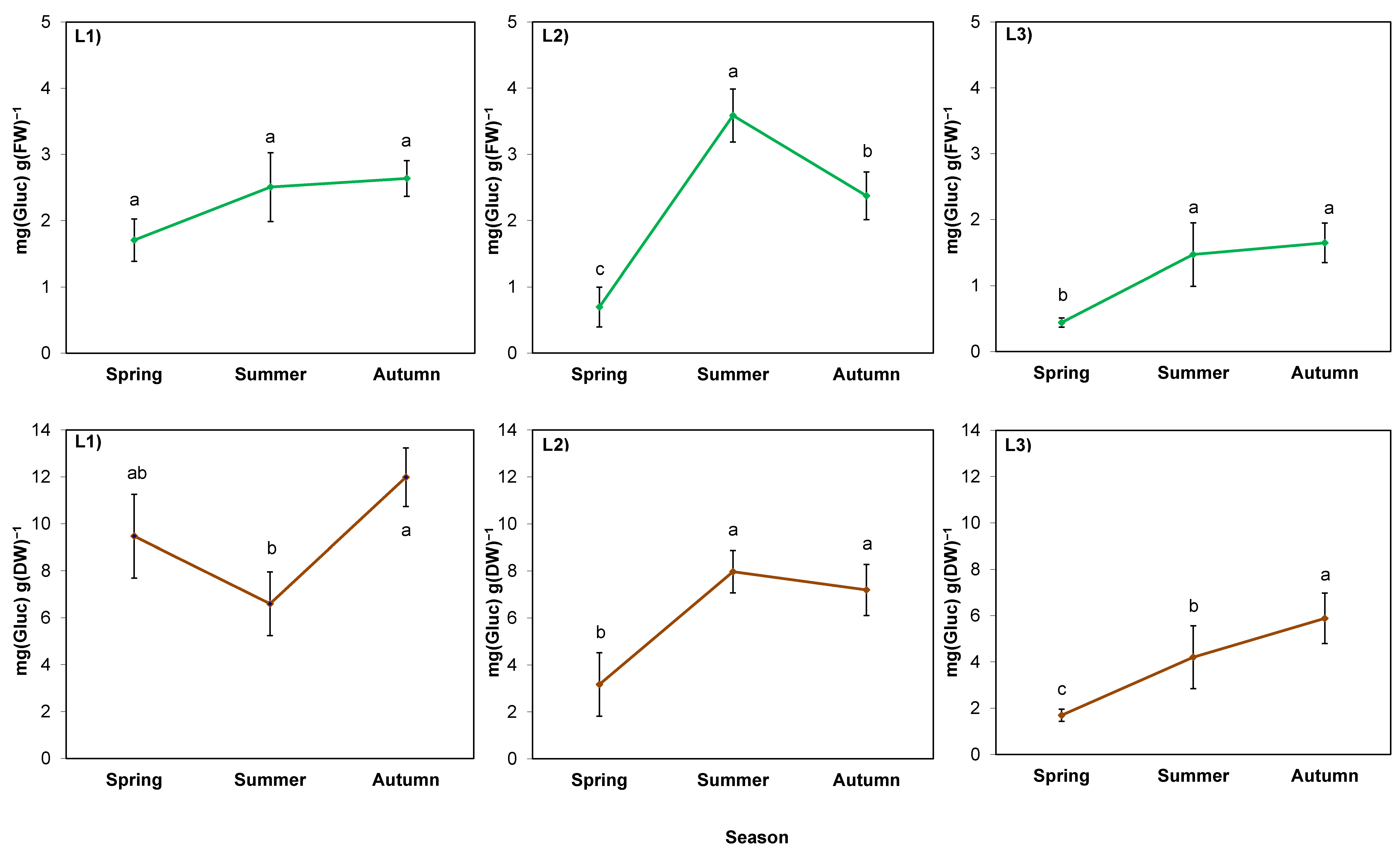
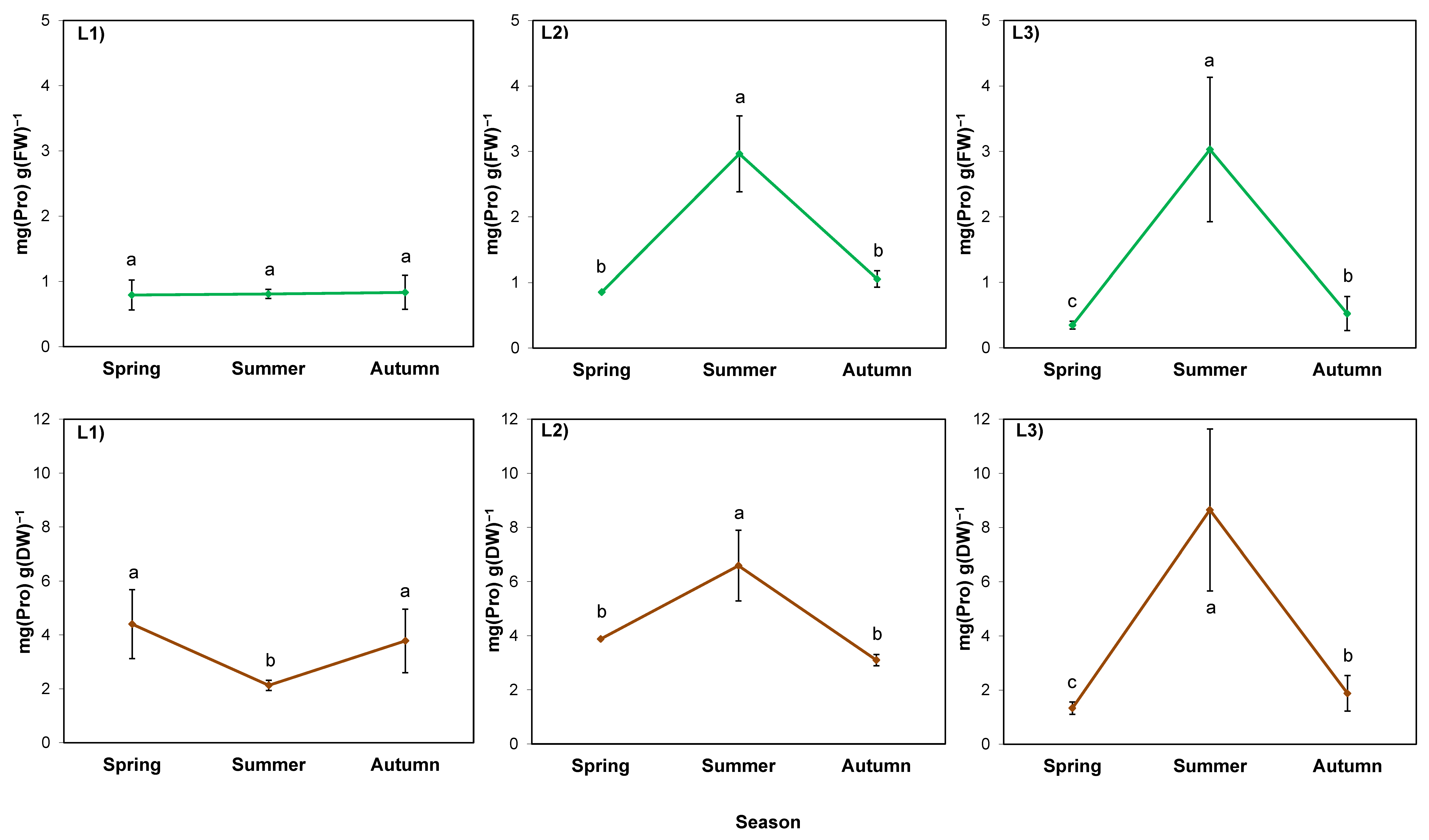
2.4. Physiological Indicators in Rhizomes
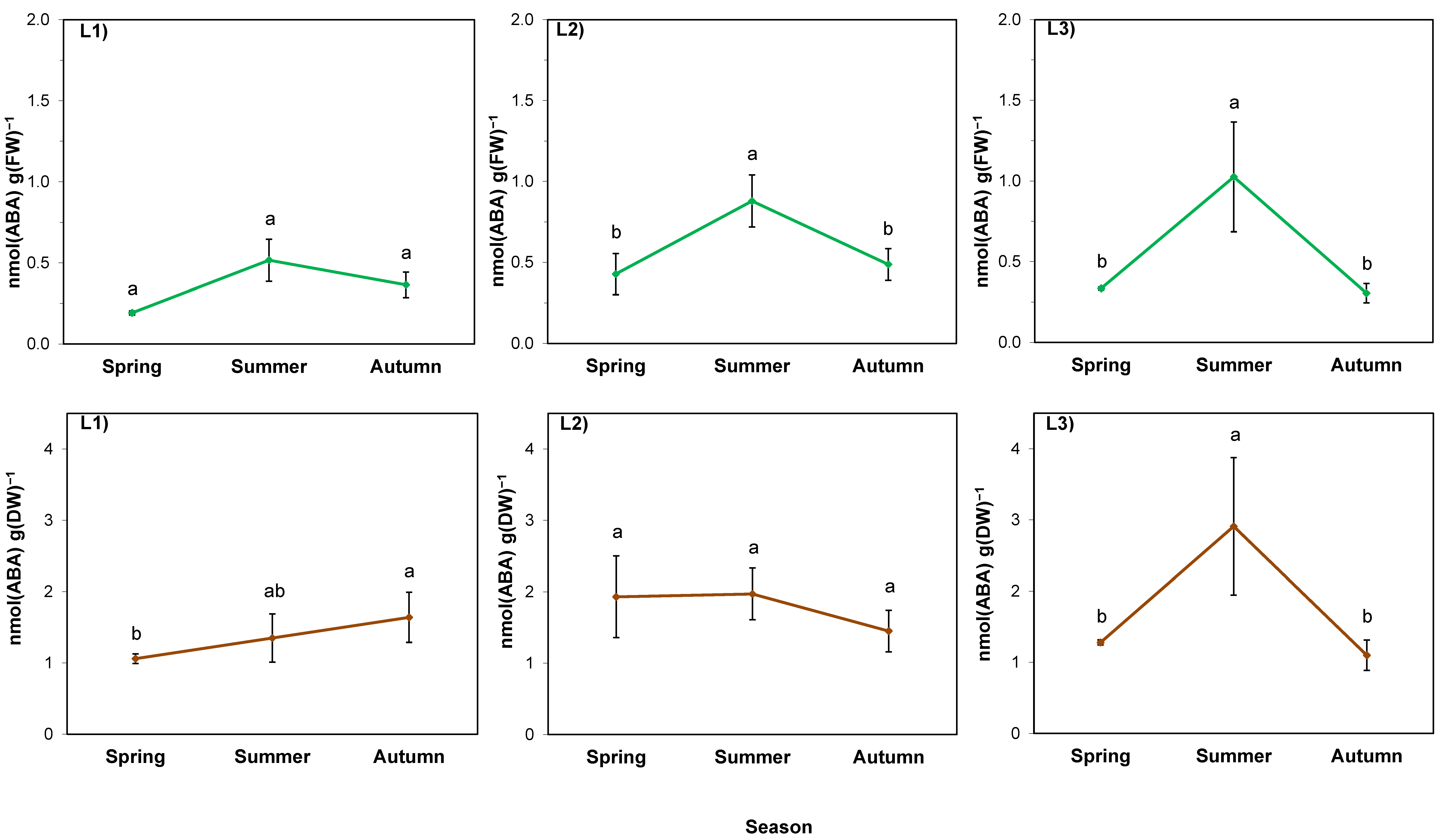
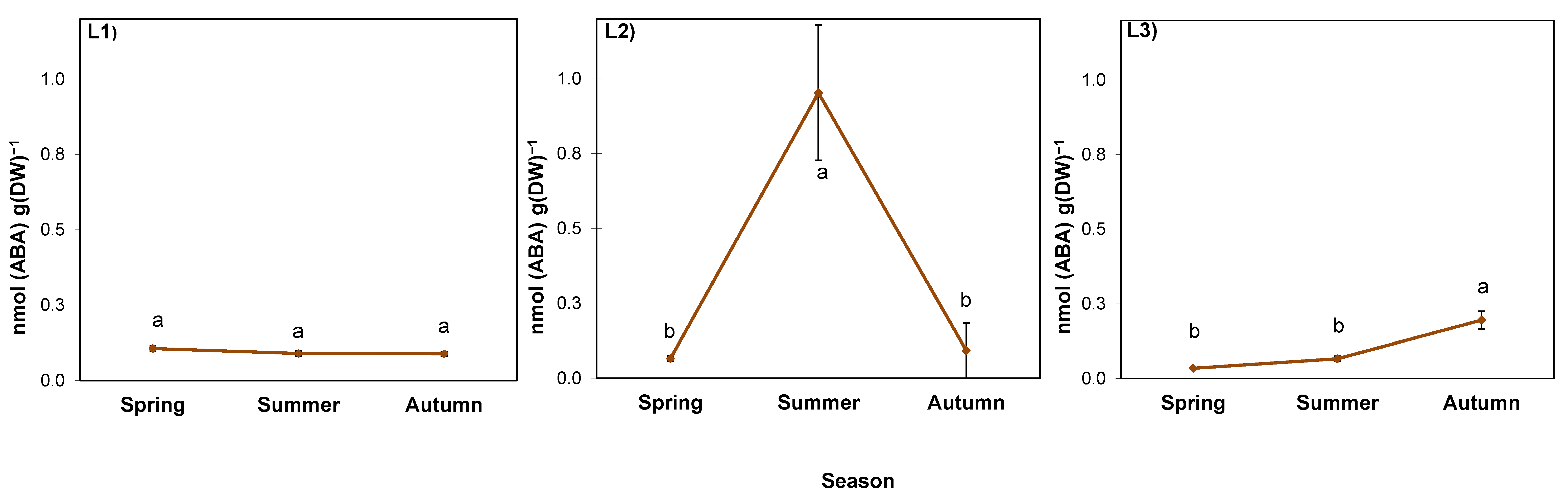
2.5. ABA in the Rhizosphere
3. Discussion
3.1. General Response of S. gigantea Plants in Studied Locations
3.2. The Response of the Aerial Parts of Plants
3.3. Alterations in Water Content in Rhizomes
3.4. Accumulation of Sugars in Rhizomes can Be a Part of the Process of Giant Goldenrod Adaptation to Dry, Hot Summer and Pre-Winter Conditions
3.5. Alterations of Proline and Abscisic Acid in Rhizomes Participate in Acclimation of S. gigantea to Local Drought and/or High-Temperature Stress, but Not to Pre-Winter Conditions
4. Materials and Methods
4.1. Location and Plant Selection
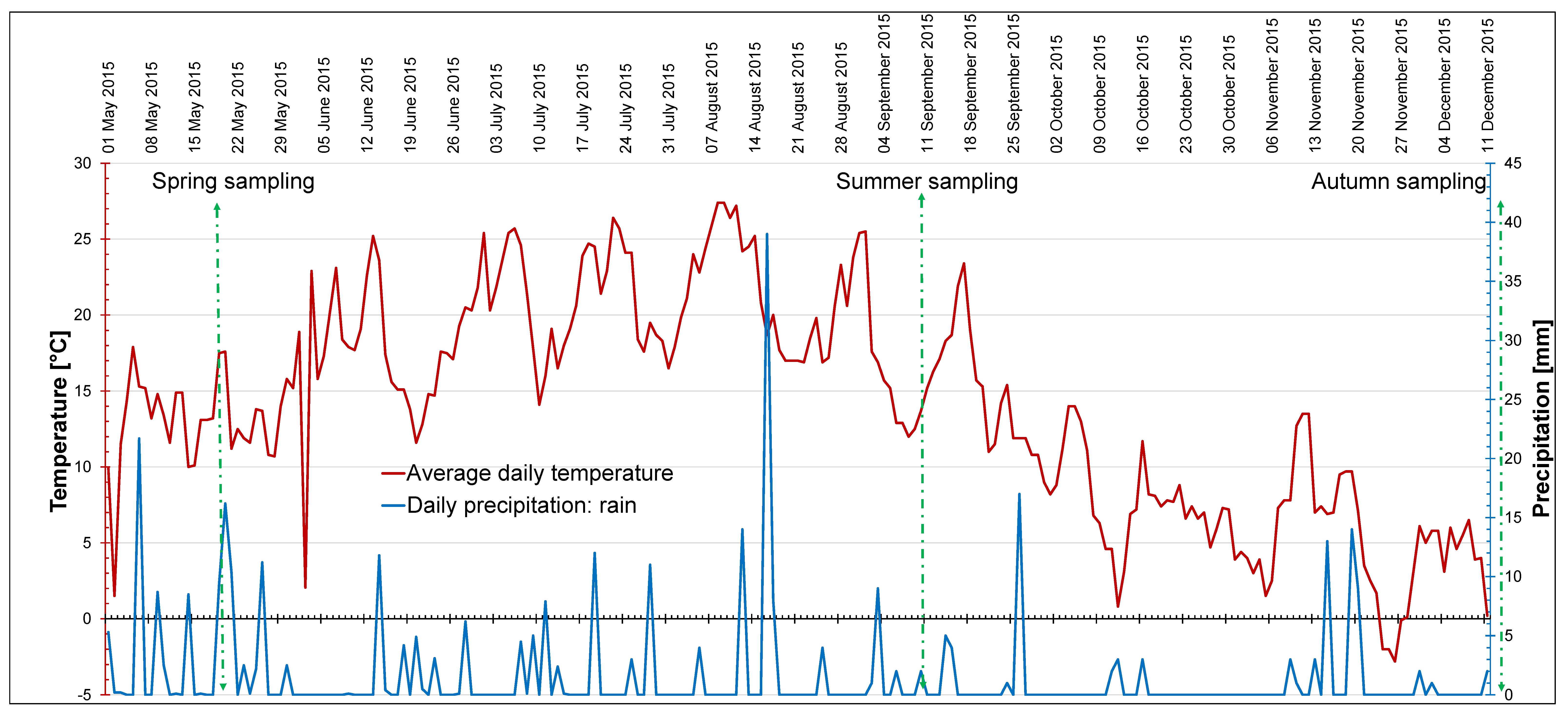
4.2. Meteorological Data and Light Conditions In Situ
4.3. Plant Analyses In Situ and Plant Material Collection
4.4. Soil Collection and Analyses
4.5. Random Amplified Polymorphic DNA (RAPD) Analysis on Leaves
4.6. Measurement of Rhizome Water Content
4.7. Measurement of Rhizome Sugar Content
4.8. Measurement of Rhizome Proline Content
4.9. Measurement of Abscisic Acid Content in the Rhizomes and Rhizosphere
4.10. Statistical Analysis
5. Conclusions
- The response of S. gigantea plants depends on changing environmental conditions in the specific habitat (location), not on genetic distance, because soluble sugar, proline, and abscisic acid in rhizomes during the growing season differ between all locations, as well as between L2 and L3, which are genetically similar.
- In all locations, the soluble sugar accumulation in goldenrod rhizomes is high in pre-winter conditions, which is definitely a part of winter hardening.
- Soil shallowness triggers intensive metabolic changes in goldenrod plants towards the accumulation of sugars and ABA in L2 rhizomes.
- Giant goldenrod plants grown in shaded locations invest their resources into shoot growth in summer to obtain more light, and during the vegetative season the changes in soluble sugar, proline, and abscisic acid in their rhizomes are moderate. Meanwhile, plants growing in the two unshaded locations, thus being more exposed to drought and high-temperature stress, inhibit anabolic processes in summer, which is reflected in the low chlorophyll content and shorter the aerial plant part.
- During the growing season, changes in the content of sugars and proline in rhizomes are greater than the changes in ABA level, which indicates their significant participation in plant metabolism and response to meteorological and soil factors. At the same time, in sun-exposed locations, ABA content in rhizomes and the rhizosphere is high, which suggests its role in the survival of giant goldenrod rhizomes in sites prone to continuous solar irradiation combined with high temperature, water scarcity, toxic metals, and P deficiency.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
References
- Weber, E.; Jakobs, G. Biological flora of central Europe: Solidago gigantea Aiton. Flora 2005, 200, 109–118. [Google Scholar] [CrossRef]
- Bochenek, A.; Synowiec, A.; Kondrat, B.; Szymczak, M.; Lahuta, L.B.; Gołaszewski, J. Do the seeds of Solidago gigantea Aiton have physiological determinants of invasiveness? Acta Physiol. Plant 2016, 38, 159. [Google Scholar] [CrossRef]
- Botta-Dukát, Z.; Balogh, L. The Most Important Invasive Plants in Hungary; HAS Institute of Ecology and Botany: Vácrátót, Hungary, 2008; p. 255. [Google Scholar]
- Guo, S.; Fang, F. Physiological adaptation of the invasive plant Solidago canadensis to environments. Acta Phytoecol. Sin. 2003, 27, 47–52. [Google Scholar]
- Tokarska-Guzik, B.; Bzdęga, K.; Nowak, T.; Urbisz, A.; Węgrzynek, B.; Dajdok, Z. Propozycja Listy Roślin Gatunków Obcych, Które Mogą Stanowić Zagrożenie dla Przyrody Polski i Unii Europejskiej; Wydawnictwo Uniwersytetu Śląskiego: Katowice, Poland, 2015. [Google Scholar]
- Yuan, Y.; Wang, B.; Zhang, S.; Tang, J.; Tu, C.; Hu, S.; Yong, J.W.H.; Chen, X. Enhanced allelopathy and competitive ability of invasive plant Solidago canadensis in its introduced range. J. Plant Ecol. 2013, 6, 253–263. [Google Scholar] [CrossRef]
- Lenda, M.; Skórka, P.; Kuszewska, K.; Moroń, D.; Bełcik, M.; Bączek-Kwinta, R.; Janowiak, F.; Duncan, D.H.; Vesk, P.A.; Possingham, H.P.; et al. Misinformation, internet honey trading and beekeepers drive a plant invasion. Ecol. Lett. 2021, 24, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Meyer, G.A.; Hull-Sanders, H.M. Altered patterns of growth, physiology and reproduction in invasive genotypes of Solidago gigantea (Asteraceae). Biol. Invasions 2008, 10, 303–317. [Google Scholar] [CrossRef]
- Szymura, M.; Szymura, T.H. Growth, phenology, and biomass allocation of alien Solidago species in central Europe. Plant Species Biol. 2015, 30, 245–256. [Google Scholar] [CrossRef]
- Liao, H.; Gurgel, P.C.S.; Pal, R.W.; Hooper, D.; Callaway, R.M. Solidago gigantea plants from nonnative ranges compensate more in response to damage than plants from the native range. Ecology 2016, 97, 2355–2363. [Google Scholar] [CrossRef]
- Majewska, M.L.; Rola, K.; Zubek, S. The growth and phosphorus acquisition of invasive plants Rudbeckia laciniata and Solidago gigantea are enhanced by arbuscular mycorrhizal fungi. Mycorrhiza 2017, 27, 83–94. [Google Scholar] [CrossRef]
- Stefanowicz, A.M.; Stanek, M.; Nobis, M.; Zubek, S. Few effects of invasive plants Reynoutria japonica, Rudbeckia laciniata and Solidago gigantea on soil physical and chemical properties. Sci. Total Environ. 2017, 574, 938–946. [Google Scholar] [CrossRef]
- Hanson, J.; Smeekens, S. Sugar perception and signaling—An update. Curr. Opin. Plant Biol. 2009, 12, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Hey, S.J.; Byrne, E.; Halford, N.G. The interface between metabolic and stress signalling. Ann. Bot. 2010, 105, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Lendl, A.; Reznicek, G. Two new saponins from Solidago gigantea. Sci. Pharm. 2007, 75, 111–120. [Google Scholar] [CrossRef]
- Ratiu, I.A.; Al-Suod, H.; Ligor, M.; Ligor, T.; Railean-Plugaru, V.; Buszewski, B. Complex investigation of extraction techniques applied for cyclitols and sugars isolation from different species of Solidago genus. Electrophoresis 2018, 39, 1966–1974. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Verslues, P.E.; Sharma, S. Proline metabolism and its implications for plant-environment interaction. Arab. Book 2010, 8, e0140. [Google Scholar] [CrossRef] [PubMed]
- Du, L.S.; Liu, H.Y.; Yan, M.; Li, J.M.; Li, J.S. Individual plasticity of the shade response of the invasive Solidago canadensis in China. PLoS ONE 2017, 12, e0170049. [Google Scholar] [CrossRef] [PubMed]
- Guan, C.; Cui, X.; Liu, H.Y.; Li, X.; Li, M.Q.; Zhang, Y.W. Proline biosynthesis enzyme genes confer salt tolerance to switchgrass (Panicum virgatum L.) in cooperation with polyamines metabolism. Front. Plant Sci. 2020, 11, 46. [Google Scholar] [CrossRef]
- Xie, E.; Wei, X.J.; Ding, A.Z.; Zheng, L.; Wu, X.N.; Anderson, B. Short-term effects of salt stress on the amino acids of Phragmites australis root exudates in constructed wetlands. Water 2020, 12, 569. [Google Scholar] [CrossRef]
- Xiong, J.; Zhang, L.; Fu, G.; Yang, Y.; Zhu, C.; Tao, L. Drought-induced proline accumulation is uninvolved with increased nitric oxide, which alleviates drought stress by decreasing transpiration in rice. J. Plant Res. 2012, 125, 155–164. [Google Scholar] [CrossRef]
- Behdad, A.; Mohsenzadeh, S.; Azizi, M.; Moshtaghi, N. Salinity effects on physiological and phytochemical characteristics and gene expression of two Glycyrrhiza glabra L. populations. Phytochemistry 2020, 171, 112236. [Google Scholar] [CrossRef] [PubMed]
- Biancucci, M.; Mattioli, R.; Moubayidin, L.; Sabatini, S.; Costantino, P.; Trovato, M. Proline affects the size of the root meristematic zone in Arabidopsis. BMC Plant Biol. 2015, 15, 263. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.; Funck, D.; Szabados, L.; Rentsch, D. Proline metabolism and transport in plant development. Amino Acids 2010, 39, 949–962. [Google Scholar] [CrossRef]
- Mattioli, R.; Costantino, P.; Trovato, M. Proline accumulation in plants. Plant Signal. Behav. 2009, 4, 1016–1018. [Google Scholar] [CrossRef] [PubMed]
- Janowiak, F.; Maas, B.; Dörffling, K. Importance of abscisic acid for chilling tolerance of maize seedlings. J. Plant Physiol. 2002, 159, 635–643. [Google Scholar] [CrossRef]
- Vishwakarma, K.; Upadhyay, N.; Kumar, N.; Yadav, G.; Singh, J.; Mishra, R.K.; Kumar, V.; Verma, R.; Upadhyay, R.G.; Pandey, M.; et al. Abscisic acid signaling and abiotic stress tolerance in plants: A review on current knowledge and future prospects. Front. Plant Sci. 2017, 8, 161. [Google Scholar] [CrossRef]
- Scharwies, J.D.; Dinneny, J.R. Water transport, perception, and response in plants. J. Plant Res. 2019, 132, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Trejo, C.L.; Clephan, A.L.; Davies, W.J. How do stomata read abscisic acid signals? Plant Physiol. 1995, 109, 803–811. [Google Scholar] [CrossRef]
- Płażek, A.; Dubert, F.; Janowiak, F.; Krępski, T.; Tatrzańska, M. Plant age and in vitro or in vivo propagation considerably affect cold tolerance of Miscanthus × giganteus. Eur. J. Agron. 2011, 34, 163–171. [Google Scholar] [CrossRef]
- Parent, B.; Hachez, C.; Redondo, E.; Simonneau, T.; Chaumont, F.; Tardieu, F. Drought and abscisic acid effects on aquaporin content translate into changes in hydraulic conductivity and leaf growth rate: A trans-scale approach. Plant Physiol. 2009, 149, 2000–2012. [Google Scholar] [CrossRef]
- Finkelstein, R. Abscisic acid synthesis and response. Arab. Book 2013, 2013, e0166. [Google Scholar] [CrossRef] [PubMed]
- Antoni, R.; Gonzalez-Guzman, M.; Rodriguez, L.; Peirats-Llobet, M.; Pizzio, G.A.; Fernandez, M.A.; De Winne, N.; De Jaeger, G.; Dietrich, D.; Bennett, M.J.; et al. PYRABACTIN RESISTANCE1-LIKE8 plays an important role for the regulation of abscisic acid signaling in root. Plant Physiol. 2013, 161, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Moriwaki, T.; Miyazawa, Y.; Kobayashi, A.; Takahashi, H. Molecular mechanisms of hydrotropism in seedling roots of Arabidopsis thaliana (Brassicaceae). Am. J. Bot. 2013, 100, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.G.K.; Kubelik, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphisms amplified by arbitrary primers are useful as genetic-markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef]
- Mucciarelli, M.; Ferrazzini, D.; Belletti, P. Genetic variability and population divergence in the rare Fritillaria tubiformis subsp. moggridgei Rix (Liliaceae) as revealed by RAPD analysis. PLoS ONE 2014, 9, e101967. [Google Scholar] [CrossRef]
- Krebs, C.; Mahy, G.; Matthies, D.; Schaffner, U.; Tiébré, M.-S.; Bizoux, J.-P. Taxa distribution and RAPD markers indicate different origin and regional differentiation of hybrids in the invasive Fallopia complex in central-western Europe. Plant Biol. 2010, 12, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Guo, S. Analysis of population genetic differences of the invasive plant (Solidago canadensis). Zhiwu Yanjiu 2005, 25, 197–204. [Google Scholar]
- Czortek, P.; Królak, E.; Borkowska, L.; Bielecka, A. Impacts of soil properties and functional diversity on the performance of invasive plant species Solidago canadensis L. on post-agricultural wastelands. Sci. Total Environ. 2020, 729, 139077. [Google Scholar] [CrossRef]
- Eckert, S.; Herden, J.; Stift, M.; Durka, W.; van Kleunen, M.; Joshi, J. Traces of genetic but not epigenetic adaptation in the invasive goldenrod Solidago canadensis despite the absence of population structure. Front. Ecol. Evol. 2022, 10, 856453. [Google Scholar] [CrossRef]
- Herrera, C.M.; Medrano, M.; Bazaga, P. Comparative spatial genetics and epigenetics of plant populations: Heuristic value and a proof of concept. Mol. Ecol. 2016, 25, 1653–1664. [Google Scholar] [CrossRef]
- Eamus, D.; Huete, A.; Yu, Q. Vegetation Dynamics: A Synthesis of Plant Ecophysiology, Remote Sensing and Modelling; Cambridge University Press: Cambridge, UK, 2016. [Google Scholar] [CrossRef]
- Gupta, N.; Ram, H.; Kumar, B. Mechanism of Zinc absorption in plants: Uptake, transport, translocation and accumulation. Rev. Environ. Sci. Biotechnol. 2016, 15, 89–109. [Google Scholar] [CrossRef]
- Collin, S.; Baskar, A.; Geevarghese, D.M.; Ali, M.N.V.S.; Bahubali, P.; Choudhary, R.; Lvov, V.; Tovar, G.I.; Senatov, F.; Koppala, S.; et al. Bioaccumulation of lead (Pb) and its effects in plants: A review. J. Hazard. Mater. 2022, 3, 100064. [Google Scholar] [CrossRef]
- Bączek-Kwinta, R.; Antonkiewicz, J. Differential physiological response and potential toxicological risk of white cabbage grown in zinc-spiked soil. Agronomy 2022, 12, 2186. [Google Scholar] [CrossRef]
- Dambiec, M.; Klink, A.; Polechońska, L. Concentration and translocation of trace metals in Solidago gigantea in urban areas: A potential bioindicator. Int. J. Environ. Sci. Technol. 2022, 19, 11729–11740. [Google Scholar] [CrossRef]
- Jakobs, G.; Weber, E.; Edwards, P.J. Introduced plants of the invasive Solidago gigantea (Asteraceae) are larger and grow denser than conspecifics in the native range. Divers. Distrib. 2004, 10, 11–19. [Google Scholar] [CrossRef]
- Popova, E.; Kim, H.-H. Cryobiotechnology of Korean orchid biodiversity: A case study using Cymbidium kanran. In Orchid Biology: Recent Trends & Challenges; Khasim, S.M., Hegde, S.N., González-Arnao, M.T., Thammasiri, K., Eds.; Springer: Singapore, 2020; pp. 119–135. [Google Scholar] [CrossRef]
- Pagter, M.; Lefevre, I.; Arora, R.; Hausman, J.-F. Quantitative and qualitative changes in carbohydrates associated with spring deacclimation in contrasting Hydrangea species. Environ. Exp. Bot. 2011, 72, 358–367. [Google Scholar] [CrossRef]
- Borah, A.; Hazarika, K.; Khayer, S.M. Drying kinetics of whole and sliced turmeric rhizomes (Curcuma longa L.) in a solar conduction dryer. Inf. Process. Agric. 2015, 2, 85–92. [Google Scholar] [CrossRef]
- Gupta, A.K.; Kaur, N. Sugar signalling and gene expression in relation to carbohydrate metabolism under abiotic stresses in plants. J. Biosci. 2005, 30, 761–776. [Google Scholar] [CrossRef]
- Rapacz, M.; Tokarz, K.; Janowiak, F. The initiation of elongation growth during long-term low-temperature stay of spring-type oilseed rape may trigger loss of frost resistance and changes in photosynthetic apparatus. Plant Sci. 2001, 161, 221–230. [Google Scholar] [CrossRef]
- Xu, W.; Jia, L.; Shi, W.; Liang, J.; Zhou, F.; Li, Q.; Zhang, J. Abscisic acid accumulation modulates auxin transport in the root tip to enhance proton secretion for maintaining root growth under moderate water stress. New Phytol. 2013, 197, 139–150. [Google Scholar] [CrossRef]
- Dong, J.; Jones, R.H.; Mou, P. Relationships between nutrient heterogeneity, root growth, and hormones: Evidence for interspecific variation. Plants 2018, 7, 15. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Zou, H.-F.; Wei, W.; Hao, Y.-J.; Tian, A.-G.; Huang, J.; Liu, Y.-F.; Zhang, J.-S.; Chen, S.-Y. Soybean GmbZIP44, GmbZIP62 and GmbZIP78 genes function as negative regulator of ABA signaling and confer salt and freezing tolerance in transgenic Arabidopsis. Planta 2008, 228, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Pagter, M.; Jensen, C.R.; Petersen, K.K.; Liu, F.; Arora, R. Changes in carbohydrates, ABA and bark proteins during seasonal cold acclimation and deacclimation in Hydrangea species differing in cold hardiness. Physiol. Plant. 2008, 134, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Martinez, O.; Pernas, M.; Carol, R.J.; Dolan, L. Ethylene modulates stem cell division in the Arabidopsis thaliana root. Science 2007, 317, 507–510. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.K.; Luijten, M.; Miyashima, S.; Lenhard, M.; Hashimoto, T.; Nakajima, K.; Scheres, B.; Heidstra, R.; Laux, T. Conserved factors regulate signalling in Arabidopsis thaliana shoot and root stem cell organizers. Nature 2007, 446, 811–814. [Google Scholar] [CrossRef] [PubMed]
- Charpentier, M.; Sun, J.H.; Wen, J.Q.; Mysore, K.S.; Oldroyd, G.E.D. Abscisic acid promotion of arbuscular mycorrhizal colonization requires a component of the PROTEIN PHOSPHATASE 2A Complex(1 W OPEN). Plant Physiol. 2014, 166, 2077-U1340. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Medina, M.J.; Steinkellner, S.; Vierheilig, H.; Bote, J.A.O.; Garrido, J.M.G. Abscisic acid determines arbuscule development and functionality in the tomato arbuscular mycorrhiza. New Phytol. 2007, 175, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Abrahamson, W.G.; Gadgil, M. Growth form and reproductive effort in goldenrods (Solidago, Compositae). Am. Nat. 1973, 107, 651–661. [Google Scholar] [CrossRef]
- Conti, L.; Galbiati, M.; Tonelli, C. ABA and the floralt transition. In Abscisic Acid: Metabolism, Transport and Signaling; Zhang, D.-P., Ed.; Springer: Dordrecht, The Netherlands, 2014; pp. 365–384. [Google Scholar] [CrossRef]
- Nambara, E.; Okamoto, M.; Tatematsu, K.; Yano, R.; Seo, M.; Kamiya, Y. Abscisic acid and the control of seed dormancy and germination. Seed Sci. Res. 2010, 20, 55–67. [Google Scholar] [CrossRef]
- Brookbank, B.P.; Patel, J.; Gazzarrini, S.; Nambara, E. Role of basal ABA in plant growth and development. Genes 2021, 12, 1936. [Google Scholar] [CrossRef]
- Zhao, H.; Peng, S.; Chen, Z.; Wu, Z.; Zhou, G.; Wang, X.; Qiu, Z. Abscisic Acid in Soil Facilitates Community Succession in Three Forests in China. J. Chem. Ecol. 2011, 37, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, F.; Ding, Y.; Du, H.; Zhang, Q.; Dang, X.; Cao, Y.; Dodd, I.C.; Xu, W. Abscisic acid mediates barley rhizosheath formation under mild soil drying by promoting root hair growth and auxin response. Plant Cell Environ. 2021, 44, 1935–1945. [Google Scholar] [CrossRef] [PubMed]
- Basirat, M.; Mousavi, S.M.; Abbaszadeh, S.; Ebrahimi, M.; Zarebanadkouki, M. The rhizosheath: A potential root trait helping plants to tolerate drought stress. Plant Soil 2019, 445, 565–575. [Google Scholar] [CrossRef]
- Aslam, M.M.; Karanja, J.K.; Dodd, I.C.; Waseem, M.; Weifeng, X. Rhizosheath: An adaptive root trait to improve plant tolerance to phosphorus and water deficits? Plant Cell Environ. 2022, 45, 2861–2874. [Google Scholar] [CrossRef] [PubMed]
- Pellicone, G.; Caloiero, T.; Guagliardi, I. The De Martonne aridity index in Calabria (Southern Italy). J. Maps 2019, 15, 788–796. [Google Scholar] [CrossRef]
- Ellenberg, H. Vegetation Ecology of Central Europe, 4th ed.; Cambridge University Press: Cambridge, UK, 1988; p. 731. [Google Scholar] [CrossRef]
- Stojanovic, D.B.; Krzic, A.; Matovic, B.; Orlovic, S.; Duputie, A.; Djurdjevic, V.; Galic, Z.; Stojnic, S. Prediction of the European beech (Fagus sylvatica L.) xeric limit using a regional climate model: An example from southeast Europe. Agric. For. Meteorol. 2013, 176, 94–103. [Google Scholar] [CrossRef]
- Jarosińska, E.; Bodziony, M. Czasowo-przestrzenna zmienność deszczu na zurbanizowanym obszarze Krakowa. Acta Sci. Pol. Form. Circumiectus 2019, 18, 43–55. [Google Scholar] [CrossRef]
- Cassol, D.; De Silva, F.S.P.; Falqueto, A.R.; Bacarin, M.A. An evaluation of non-destructive methods to estimate total chlorophyll content. Photosynthetica 2008, 46, 634–636. [Google Scholar] [CrossRef]
- Bączek-Kwinta, R.; Juzoń, K.; Borek, M.; Antonkiewicz, J. Photosynthetic response of cabbage in cadmium-spiked soil. Photosynthetica 2019, 57, 731–739. [Google Scholar] [CrossRef]
- Borek, M.; Bączek-Kwinta, R.; Rapacz, M. Photosynthetic activity of variegated leaves of Coleus x hybridus hort. cultivars characterised by chlorophyll fluorescence techniques. Photosynthetica 2016, 54, 331–339. [Google Scholar] [CrossRef]
- Jones, J.B., Jr.; Case, V.W. Sampling, handling, and analyzing plant tissue samples. In Soil Testing and Plant Analysis; Westerman, R.L., Ed.; Soil Science Society of America, Inc.: Madison, WI, USA, 1990; pp. 389–427. [Google Scholar] [CrossRef]
- Ostrowska, A.; Gawliński, S.; Szczubiałka, Z. Methods of Analysis and Assessment of Soil and Plant Properties; A Catalgoue; Institute of Environmental Protection–National Research Institute: Warsaw, Poland, 1991; Volume 334. [Google Scholar]
- van de Wiel, H.J. Determination of Elements by ICP-AES and ICP-MS; National Institute of Public Health and the Environment (RIVM); Horizontal: Bilthoven, The Netherlands, 2003. [Google Scholar]
- Wedepohl, K.H. The composition of the continental-crust. Geochim. Cosmochim. Acta 1995, 59, 1217–1232. [Google Scholar] [CrossRef]
- Gope, M.; Masto, R.E.; George, J.; Hoque, R.R.; Balachandran, S. Bioavailability and health risk of some potentially toxic elements (Cd, Cu, Pb and Zn) in street dust of Asansol, India. Ecotoxicol. Environ. Saf. 2017, 138, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Kabir, M.H.; Rashid, M.H.; Wang, Q.Y.; Wang, W.Q.; Lu, S.L.; Yonemochi, S. Determination of Heavy Metal Contamination and Pollution Indices of Roadside Dust in Dhaka City, Bangladesh. Processes 2021, 9, 1732. [Google Scholar] [CrossRef]
- Simlat, M.; Ptak, A.; Kula, A.; Orzel, A. Assessment of genetic variability among raspberry accessions using molecular markers. Acta Sci. Pol.-Hortorum Cultus 2018, 17, 61–72. [Google Scholar] [CrossRef]
- Ashwell, G. Colorimetric analysis of sugars. Methods Enzymol. 1957, 3, 73–105. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Carillo, P.; Mastrolonardo, G.; Nacca, F.; Parisi, D.; Verlotta, A.; Fuggi, A. Nitrogen metabolism in durum wheat under salinity: Accumulation of proline and glycine betaine. Funct. Plant Biol. 2008, 35, 412–426. [Google Scholar] [CrossRef] [PubMed]
- Hummel, I.; Pantin, F.; Sulpice, R.; Piques, M.; Rolland, G.; Dauzat, M.; Christophe, A.; Pervent, M.; Bouteille, M.; Stitt, M.; et al. Arabidopsis plants acclimate to water deficit at low cost through changes of carbon usage: An integrated perspective using growth, metabolite, enzyme, and gene expression analysis. Plant Physiol. 2010, 154, 357–372. [Google Scholar] [CrossRef]
- Walker-Simmons, M.K.; Abrams, S.R. Use of ABA immunoassays. In Abscisic Acid, Physiology and Biochemistry; Davies, W.J., Jones, H.G., Eds.; Bios Scientific Publishers: Oxford, UK, 1991; pp. 53–63. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location Number | Description | PPFD (µmol (Quantum) m−2 s−1) | ||
|---|---|---|---|---|
| Spring | Summer | Autumn | ||
| L1 | Permanent shade caused by the proximity of tall trees | 42–406 | 1134–652 | 32–212 |
| L2 | No shade; plants lit with natural sunlight for the most part of the day | 803–1198 | 830–1299 | 153–469 |
| L3 | No shade; plants lit with natural sunlight for the most part of the day | 807–1122 | 817–1301 | 149–484 |
| Parameter | Unit | L1 | L2 | L3 |
|---|---|---|---|---|
| Thickness | cm | 100–204 a | 5–32 c | 53–107 b |
| pH(H2O) | - | 5.82 ± 0.003 a | 5.71 ± 0.006 a | 5.73 ± 0.003 a |
| pH(KCl) | - | 5.66 ± 0.013 a | 5.32 ± 0.001 a | 5.35 ± 0.024 a |
| Macronutrients | ||||
| C organic | % | 1.948 ± 0.003 b | 2.355 ± 0.013 a | 1.885 ± 0.016 c |
| N total | % | 0.204 ± 0.001 a | 0.209 ± 0.001 a | 0.186 ± 0.002 b |
| P | mg ∙ kg−1 DW | 118.7 ± 1.4 a | 92.4 ± 1.5 b | 53.3 ± 0.1 c |
| Ca | mg ∙ kg−1 DW | 2395.6 ± 23.7 b | 3015.9 ± 22.3 a | 2348.7 ± 24.9 b |
| Na | mg ∙ kg−1 DW | 4.52 ± 0.38 b | 10.48 ± 0.87 a | 9.09 ± 1.04 a |
| S | mg ∙ kg−1 DW | 25.7 ± 0.7 b | 38.7 ± 0.6 a | 26.5 ± 0.4 b |
| Metals | ||||
| Zn | mg ∙ kg−1 DW | 43.1 ± 1.4 b | 212.3 ± 2.8 a | 44.2 ± 0.4 b |
| Ni | mg ∙ kg−1 DW | 1.11 ± 0.03 a | 0.86 ± 0.01 b | 1.21 ± 0.01 a |
| Cd | mg ∙ kg−1 DW | 0.60 ± 0.01 a | 0.33 ± 0.00 b | 0.35 ± 0.00 b |
| Pb | mg ∙ kg−1 DW | 10.8 ± 0.0 c | 23.3 ± 0.1 a | 15.1 ± 0.2 b |
| Al | mg ∙ kg−1 DW | 530.7 ± 21.7 b | 619.4 ± 4.6 a | 530.8 ± 5.6 b |
| Metal | Location | ||
|---|---|---|---|
| L1 | L2 | L3 | |
| Zn | 0.83 | 4.08 | 0.85 |
| Ni | 0.06 | 0.05 | 0.07 |
| Cd | 5.88 | 3.24 | 3.43 |
| Pb | 0.64 | 1.37 | 0.89 |
| Al | 0.01 | 0.01 | 0.01 |
| Primer | Primer Sequence (5′→3′) | Total Number of Amplified Bands | The Number of Polymorphic Bands | % of Polymorphic Bands |
|---|---|---|---|---|
| OPA-01 | CAGGCCCTTC | 20 | 16 | 80.0 |
| OPA-02 | TGCCGAGCTG | 13 | 9 | 69.2 |
| OPA-03 | AGTCAGCCAC | 18 | 14 | 77.8 |
| OPA-10 | GTGATCGCAG | 13 | 13 | 100.0 |
| OPA-11 | CAATCGCCGT | 17 | 15 | 88.2 |
| OPA-13 | CAGCACCCAC | 20 | 16 | 80.0 |
| OPA-19 | CAAACGTCGG | 15 | 15 | 100.0 |
| OPB-07 | GGTGACGCAG | 30 | 28 | 93.3 |
| OPB-11 | GTAGACCCGT | 16 | 16 | 100.0 |
| OPR-01 | TGCGGGTCCT | 15 | 9 | 60.0 |
| OPR-02 | CACAGCTGCC | 23 | 19 | 82.6 |
| OPR-03 | ACACAGAGGG | 14 | 14 | 100.0 |
| OPR-04 | CCCGTAGCAC | 20 | 20 | 100.0 |
| OPR-05 | GACCTAGTGG | 7 | 7 | 100.0 |
| OPR-06 | GTCTACGGCA | 17 | 15 | 88.2 |
| Total | 258 | 226 | ||
| Average/primer | 17.20 | 15.13 | 87.95 |
| L1 | L2 | L3 | |
|---|---|---|---|
| L1 | 1.0000 | ||
| L2 | 0.2099 | 1.0000 | |
| L3 | 0.2278 | 0.4326 | 1.0000 |
| Location | Greenness Index (CL-01, Arbitrary Units) | Plant Height (cm) | ||||
|---|---|---|---|---|---|---|
| Season | Season | |||||
| Spring | Summer | Autumn | Spring | Summer | Autumn | |
| L1 | 8.68 b,B 100% | 13.1 a,A 151% | Leaves dried | 37.3 c,B 100% | 157.3 b,B 421% | 209.7 a,A 562% |
| L2 | 10.9 a,A 100% | 8.59 b,B 79% | Leaves dried | 53.2 b,A 100% | 166.7 a,AB 314% | 171.3 a,B 322% |
| L3 | 7.90 b,B 100% | 13.5 a,A 171% | Leaves dried | 52.7 c,A 100% | 179.0 b,A 340% | 206.3 a,A 391% |
| Factor | Water Content | Sugars | Proline | ABA | ||||
|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | |
| Season (S) | 53.10 | 0.000 *** | 16.36 | 0.000 *** | 13.26 | 0.008 ** | 8.51 | 0.002 ** |
| Location (L) | 8.79 | 0.002 ** | 8.92 | 0.002 ** | 2.82 | 0.086 | 33.83 | 0.000 *** |
| S × L | 3.55 | 0.026 * | 2.69 | 0.065 | 3.46 | 0.029 * | 3.39 | 0.027 * |
| Parameters Tested for Correlation | Correlation Coefficient in Specific Location | ||
|---|---|---|---|
| L1 | L2 | L3 | |
| Water content/water-soluble sugar content | −0.4386 ns | −0.8103 ** | −0.5368 ns |
| Water content/proline content | 0.0191 ns | −0.3183 ns | −0.5447 ns |
| Water content/ABA content | −0.7489 ** | −0.7580 ** | −0.7156 ** |
| Water-soluble sugar content/proline content | −0.2260 ns | 0.5357 ns | 0.0764 ns |
| Water-soluble sugar content/ABA content | 0.6286 ns | 0.5750 ns | 0.3024 ns |
| ABA content/proline content | 0.0931 ns | 0.1757 ns | 0.5335 ns |
| Factor | ABA | |
|---|---|---|
| Test Value F | p | |
| Season (S) | 326.9 | 0.002 ** |
| Location (L) | 294.3 | 0.000 *** |
| S × L | 341.5 | 0.000 *** |
| Parameter | Description/Formula and Unit | Value |
|---|---|---|
| P * | Total precipitation [mm] | 345.1 |
| Pm * | Mean precipitation [mm] | 1.53 |
| Tm * | Mean air temperature [°C] | 14.0 |
| Tmax * | Maximal air temperature [°C] | 27.4 |
| De Martonne aridity index (DMI) * | P/(Tm + 10) [mm/°C] | 14.4 |
| Ellenberg climate quotient (EQ) * | (Tmax * × 1000)/P | 31.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bączek-Kwinta, R.; Janowiak, F.; Simlat, M.; Antonkiewicz, J. Involvement of Dynamic Adjustment of ABA, Proline and Sugar Levels in Rhizomes in Effective Acclimation of Solidago gigantea to Contrasting Weather and Soil Conditions in the Country of Invasion. Int. J. Mol. Sci. 2023, 24, 15368. https://doi.org/10.3390/ijms242015368
Bączek-Kwinta R, Janowiak F, Simlat M, Antonkiewicz J. Involvement of Dynamic Adjustment of ABA, Proline and Sugar Levels in Rhizomes in Effective Acclimation of Solidago gigantea to Contrasting Weather and Soil Conditions in the Country of Invasion. International Journal of Molecular Sciences. 2023; 24(20):15368. https://doi.org/10.3390/ijms242015368
Chicago/Turabian StyleBączek-Kwinta, Renata, Franciszek Janowiak, Magdalena Simlat, and Jacek Antonkiewicz. 2023. "Involvement of Dynamic Adjustment of ABA, Proline and Sugar Levels in Rhizomes in Effective Acclimation of Solidago gigantea to Contrasting Weather and Soil Conditions in the Country of Invasion" International Journal of Molecular Sciences 24, no. 20: 15368. https://doi.org/10.3390/ijms242015368
APA StyleBączek-Kwinta, R., Janowiak, F., Simlat, M., & Antonkiewicz, J. (2023). Involvement of Dynamic Adjustment of ABA, Proline and Sugar Levels in Rhizomes in Effective Acclimation of Solidago gigantea to Contrasting Weather and Soil Conditions in the Country of Invasion. International Journal of Molecular Sciences, 24(20), 15368. https://doi.org/10.3390/ijms242015368