
Channel Formation in Cry Toxins: An Alphafold-2 Perspective


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Cry Toxins
1.2. Structure of Cry Toxins
1.3. Interaction of Cry Toxins with Membranes
1.4. Proposed Pre-Pore Oligomer
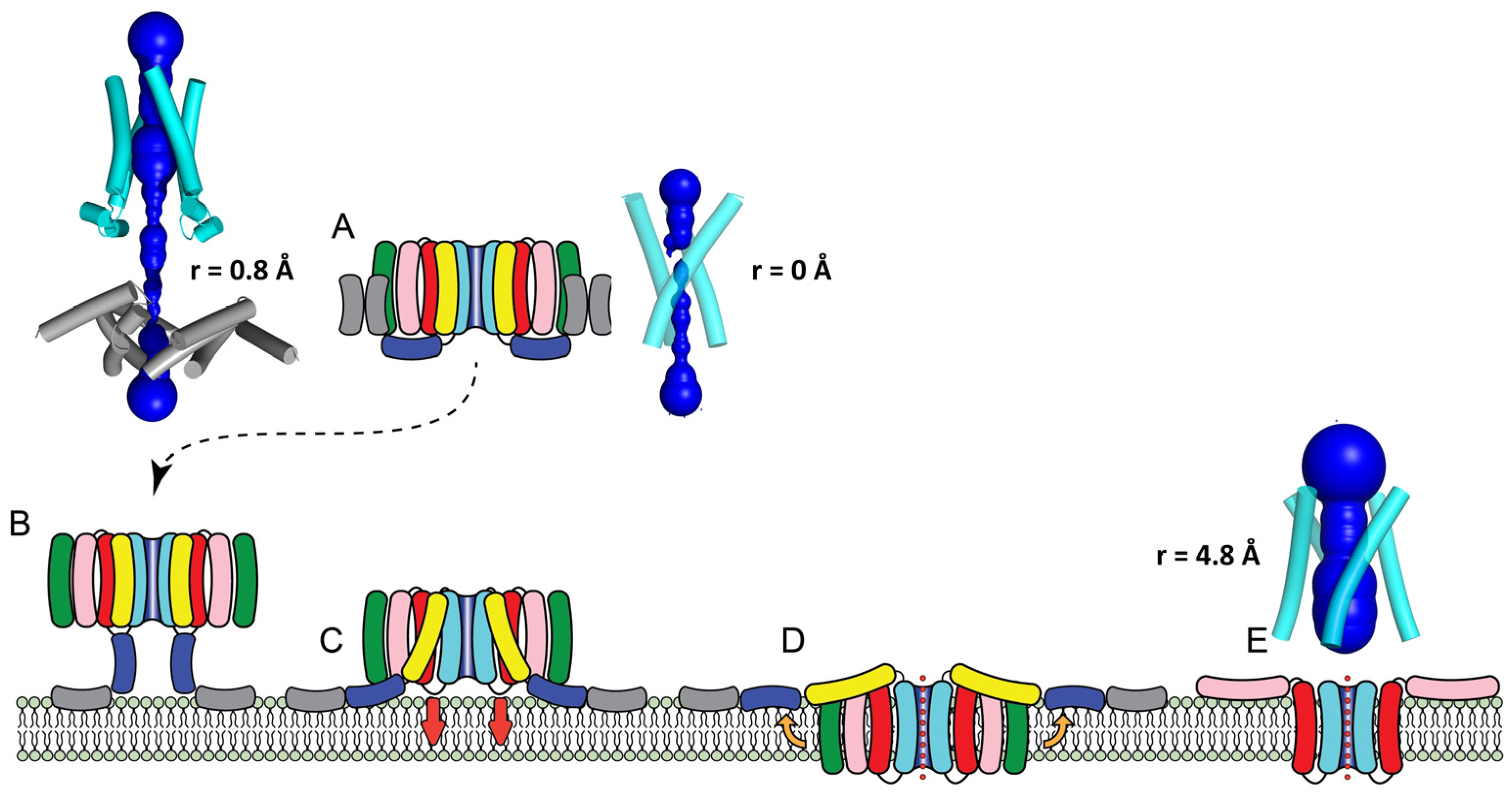
2. Results
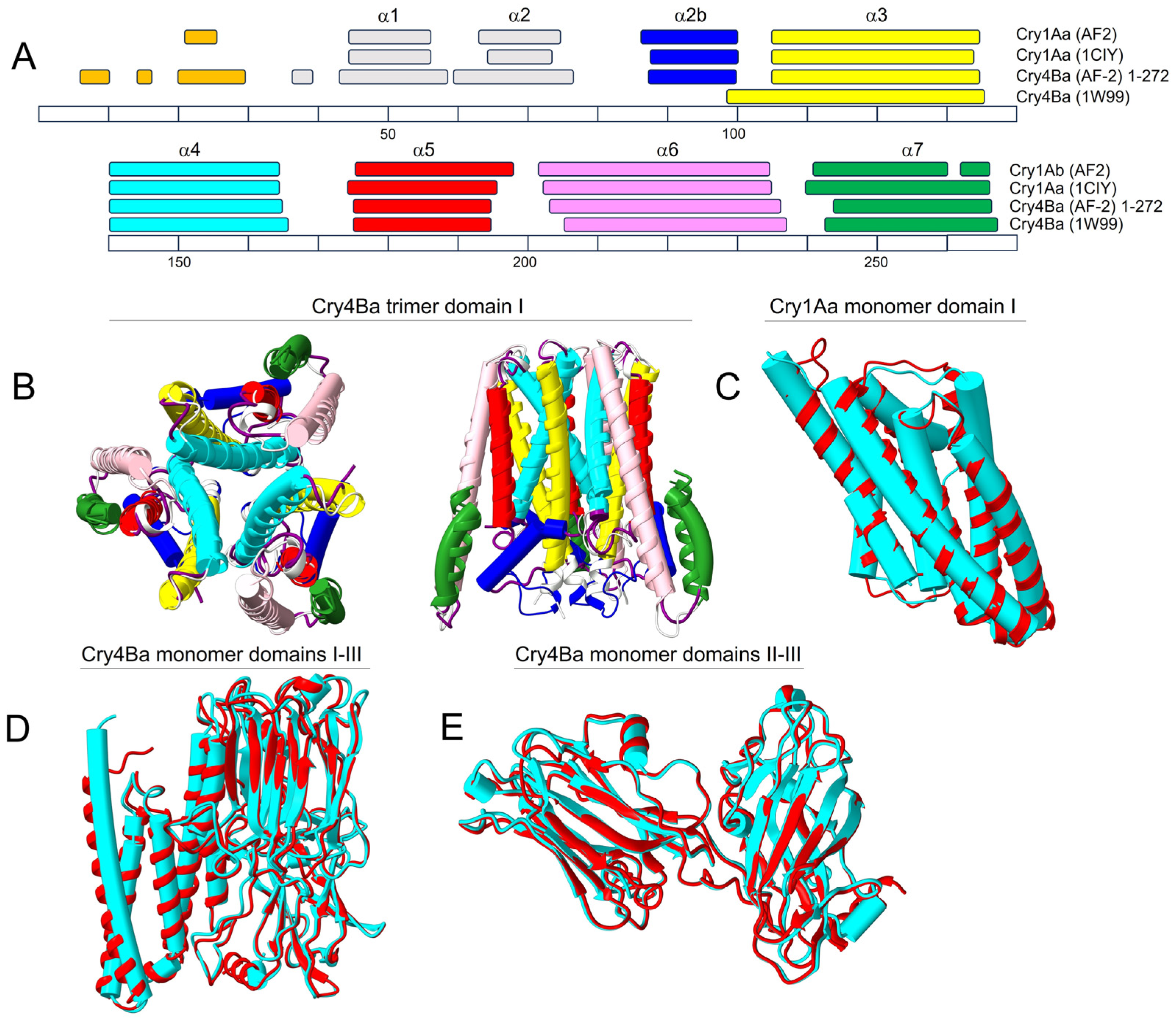
2.1. AF2 Prediction of Cry Toxin Structure


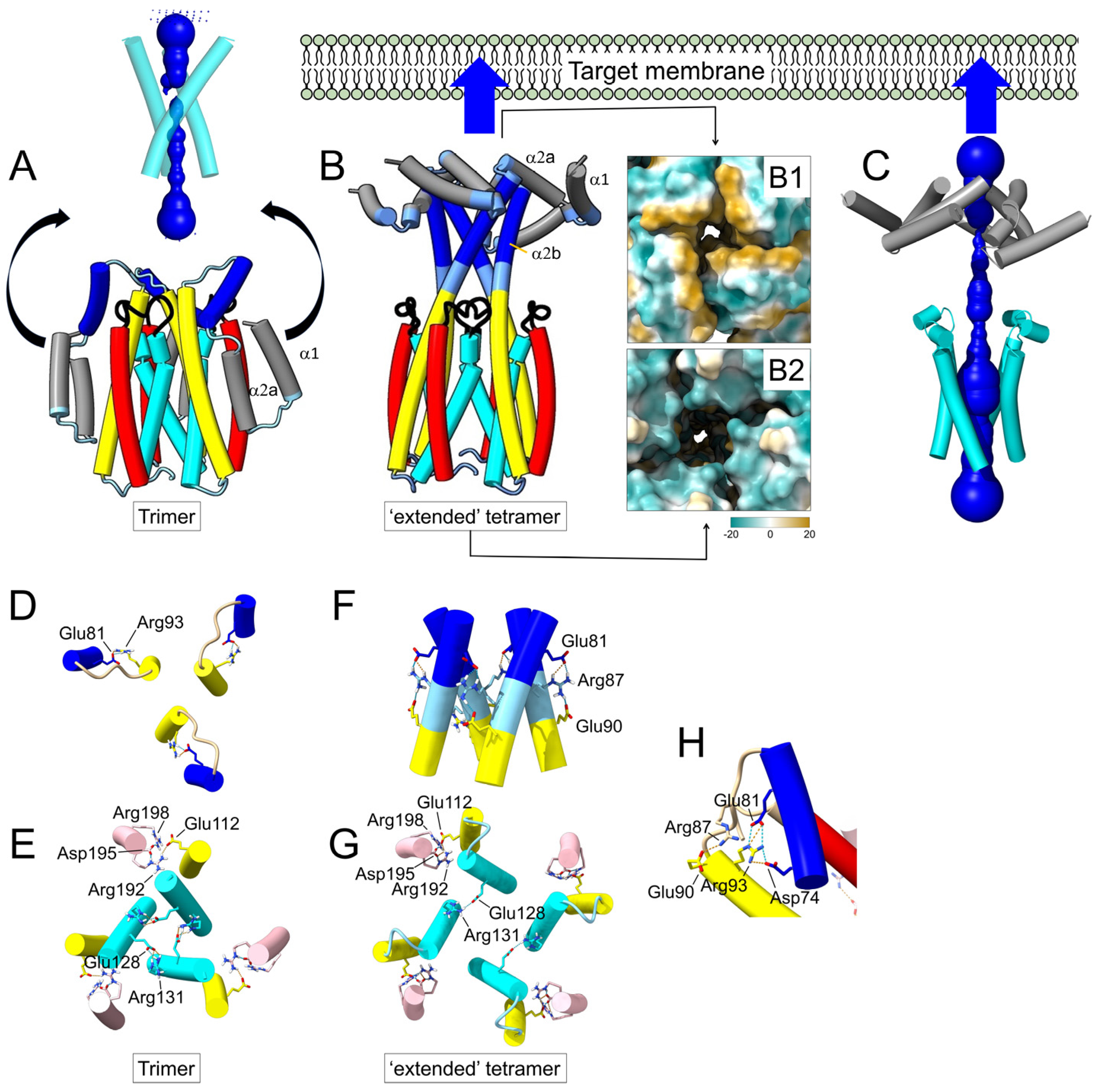
2.2. Prediction of a Tetrameric Oligomer of Domain I in Cry1Aa Toxin
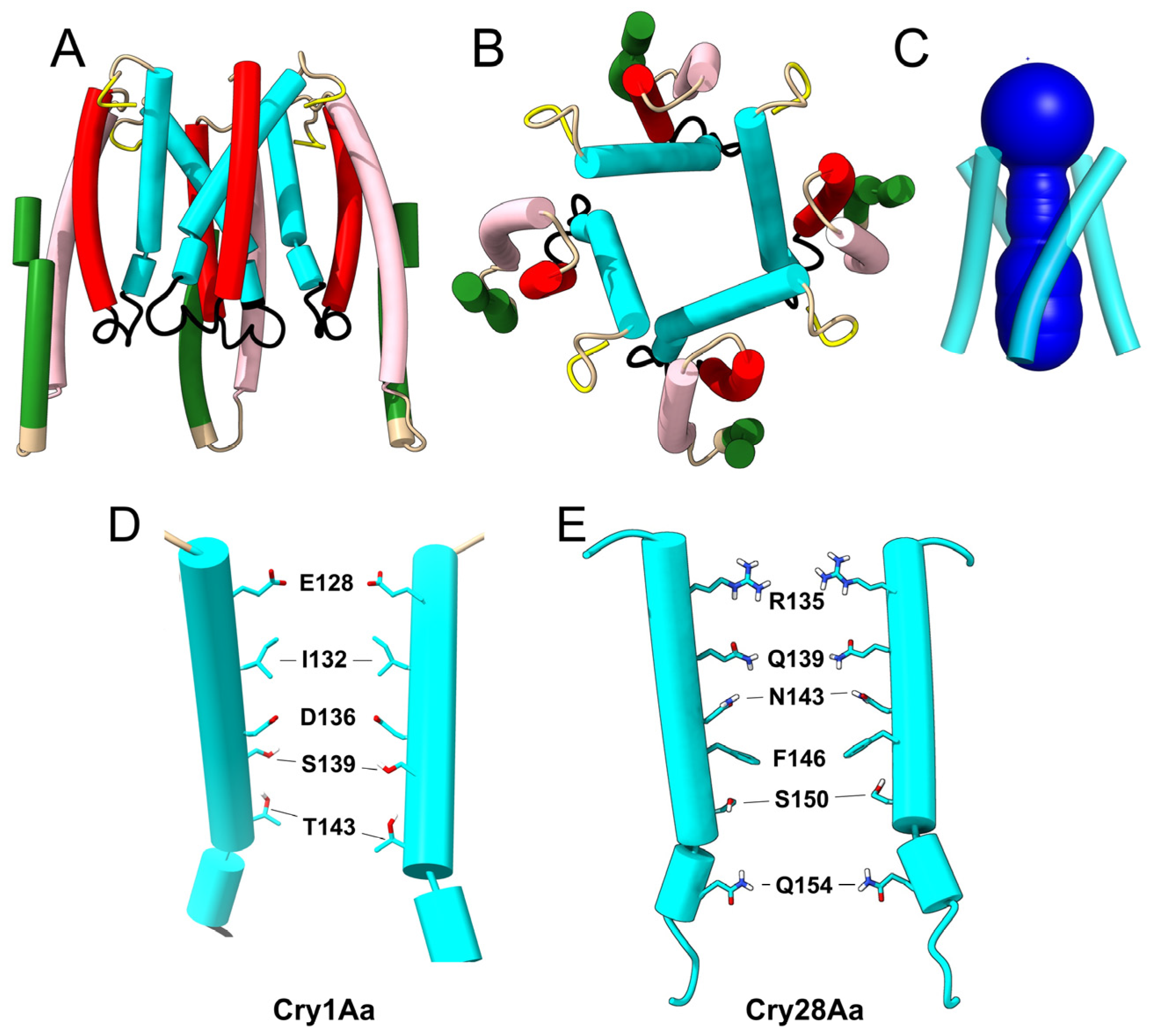
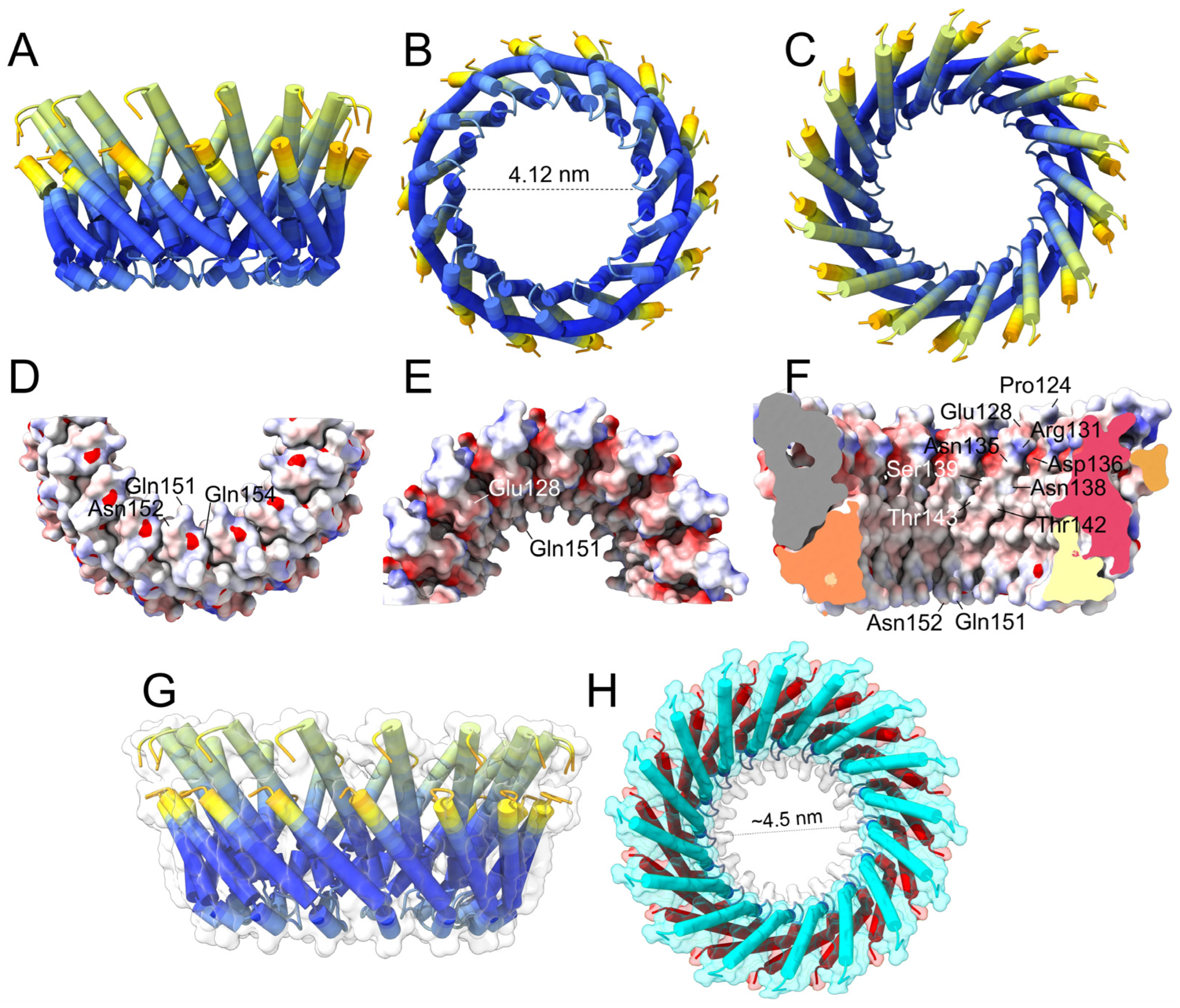
2.3. Prediction of Oligomers Formed by the α4–α5 Hairpin
3. Discussion
3.1. AF2 Can Accurately Predict the Crystal Structures of Cry1Aa and Cry4Ba
3.2. The Extended Cry1Aa Tetrameric Intermediate
3.3. The Cry1Aa Tetrameric Membrane-Inserted Channel
3.4. Pores Made of α4/α5 Hairpins
3.5. Testing the Validity of the Tetrameric Models for Cry1Aa
4. Materials and Methods
Sequences of Cry Toxin and Structure Prediction
5. Conclusions
- AF2 predicts the structures of three-domain Cry toxins with high accuracy.
- For Cry1Aa, AF2 predicted two tetrameric forms: (1) an ‘extended’ intermediate and (2) a membrane-inserted model, which would be responsible for channel activity.
- We provide a detailed description of the residues in α4 that line the lumen of the Cry1Aa tetrameric channel, which are in agreement with experimental data.
- AF2 predicts an intriguing large ensemble of α4–α5 hairpins that may be part of the toxicity mechanism. This large pore may contribute to osmotic lysis, or it may be able to facilitate the entry of toxins into the cell, leading to disruption of homeostasis and death.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pardo-López, L.; Soberón, M.; Bravo, A. Bacillus thuringiensis insecticidal three-domain Cry toxins: Mode of action, insect resistance and consequences for crop protection. FEMS Microbiol. Rev. 2013, 37, 3–22. [Google Scholar] [CrossRef]
- Sanahuja, G.; Banakar, R.; Twyman, R.M.; Capell, T.; Christou, P. Bacillus thuringiensis: A century of research, development and commercial applications. Plant Biotechnol. J. 2011, 9, 283–300. [Google Scholar] [CrossRef]
- Gould, E.; Solomon, T. Pathogenic flaviviruses. Lancet 2008, 371, 500–509. [Google Scholar] [CrossRef]
- Pierson, T.C.; Diamond, M.S. The continued threat of emerging flaviviruses. Nat. Microbiol. 2020, 5, 796–812. [Google Scholar] [CrossRef]
- Crickmore, N.; Berry, C.; Panneerselvam, S.; Mishra, R.; Connor, T.R.; Bonning, B.C. A structure-based nomenclature for Bacillus thuringiensis and other bacteria-derived pesticidal proteins. J. Invertebr. Pathol. 2021, 186, 107438. [Google Scholar] [CrossRef]
- Evdokimov, A.G.; Moshiri, F.; Sturman, E.J.; Rydel, T.J.; Zheng, M.; Seale, J.W.; Franklin, S. Structure of the full-length insecticidal protein Cry1Ac reveals intriguing details of toxin packaging into in vivo formed crystals. Protein Sci. 2014, 23, 1491–1497. [Google Scholar] [CrossRef]
- Peña-Cardeña, A.; Grande, R.; Sánchez, J.; Tabashnik, B.E.; Bravo, A.; Soberón, M.; Gómez, I. The C-terminal protoxin region of Bacillus thuringiensis Cry1Ab toxin has a functional role in binding to GPI-anchored receptors in the insect midgut. J. Biol. Chem. 2018, 293, 20263–20272. [Google Scholar] [CrossRef]
- Wu, Y. Detection and Mechanisms of Resistance Evolved in Insects to Cry Toxins from Bacillus thuringiensis. Adv. Insect Physiol. 2014, 47, 297–342. [Google Scholar]
- Heckel, D.G. How do toxins from Bacillus thuringiensis kill insects? An evolutionary perspective. Arch. Insect Biochem. Physiol. 2020, 104, e21673. [Google Scholar] [CrossRef]
- Pigott, C.R.; Ellar, D.J. Role of receptors in Bacillus thuringiensis crystal toxin activity. Microbiol. Mol. Biol. Rev. 2007, 71, 255–281. [Google Scholar] [CrossRef]
- Endo, H. Molecular and Kinetic Models for Pore Formation of Bacillus thuringiensis Cry Toxin. Toxins 2022, 14, 433. [Google Scholar] [CrossRef]
- Zhang, X.; Candas, M.; Griko, N.B.; Taussig, R.; Bulla, L.A., Jr. A mechanism of cell death involving an adenylyl cyclase/PKA signaling pathway is induced by the Cry1Ab toxin of Bacillus thuringiensis. Proc. Natl. Acad. Sci. USA 2006, 103, 9897–9902. [Google Scholar] [CrossRef]
- Li, J.; Carroll, J.; Ellar, D.J. Crystal structure of insecticidal δ-endotoxin from Bacillus thuringiensis at 2.5 Å resolution. Nature 1991, 353, 815–821. [Google Scholar] [CrossRef]
- Boonserm, P.; Davis, P.; Ellar, D.J.; Li, J. Crystal structure of the mosquito-larvicidal toxin Cry4Ba and its biological implications. J. Mol. Biol. 2005, 348, 363–382. [Google Scholar] [CrossRef]
- Thamwiriyasati, N.; Kanchanawarin, C.; Imtong, C.; Chen, C.J.; Li, H.C.; Angsuthanasombat, C. Complete structure elucidation of a functional form of the Bacillus thuringiensis Cry4Ba δ-endotoxin: Insights into toxin-induced transmembrane pore architecture. Biochem. Biophys. Res. Commun. 2022, 620, 158–164. [Google Scholar] [CrossRef]
- Boonserm, P.; Mo, M.; Angsuthanasombat, C.; Lescar, J. Structure of the functional form of the mosquito larvicidal Cry4Aa toxin from Bacillus thuringiensis at a 2.8-angstrom resolution. J. Bacteriol. 2006, 188, 3391–3401. [Google Scholar] [CrossRef]
- Grochulski, P.; Masson, L.; Borisova, S.; Pusztai-Carey, M.; Schwartz, J.-L.; Brousseau, R.; Cygler, M. Bacillus thuringiensis CryIA(a) insecticidal toxin: Crystal structure and channel formation. J. Mol. Biol. 1995, 254, 447–464. [Google Scholar] [CrossRef]
- Morse, R.J.; Yamamoto, T.; Stroud, R.M. Structure of Cry2Aa suggests an unexpected receptor binding epitope. Structure 2001, 9, 409–417. [Google Scholar] [CrossRef]
- Galitsky, N.; Cody, V.; Wojtczak, A.; Ghosh, D.; Luft, J.R.; Pangborn, W.; English, L. Structure of the insecticidal bacterial δ-endotoxin Cry3Bb1 of Bacillus thuringiensis. Acta Crystallogr. Sect. D Biol. Crystallogr. 2001, 57, 1101–1109. [Google Scholar] [CrossRef]
- Guo, S.; Ye, S.; Liu, Y.; Wei, L.; Xue, J.; Wu, H.; Song, F.; Zhang, J.; Wu, X.; Huang, D.; et al. Crystal structure of Bacillus thuringiensis Cry8Ea1: An insecticidal toxin toxic to underground pests, the larvae of Holotrichia parallela. J. Struct. Biol. 2009, 168, 259–266. [Google Scholar] [CrossRef]
- Hui, F.; Scheib, U.; Hu, Y.; Sommer, R.J.; Aroian, R.V.; Ghosh, P. Structure and glycolipid binding properties of the nematicidal protein Cry5B. Biochemistry 2012, 51, 9911–9921. [Google Scholar] [CrossRef]
- Li, J.; Wang, L.; Kotaka, M.; Lee, M.M.; Chan, M.K. Insights from the Structure of an Active Form of Bacillus thuringiensis Cry5B. Toxins 2022, 14, 823. [Google Scholar] [CrossRef]
- Jing, X.; Yuan, Y.; Wu, Y.; Wu, D.; Gong, P.; Gao, M. Crystal structure of Bacillus thuringiensis Cry7Ca1 toxin active against Locusta migratoria manilensis. Protein Sci. 2019, 28, 609–619. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.; Fu, X.; Nageotte, J.R.; Silverman, J.; Bretsnyder, E.C.; Chen, D.; Rydel, T.J.; Bean, G.J.; Li, K.S.; et al. Bacillus thuringiensis Cry1Da_7 and Cry1B.868 Protein Interactions with Novel Receptors Allow Control of Resistant Fall Armyworms, Spodoptera frugiperda (J.E. Smith). Appl Environ. Microbiol 2019, 85, e00579-19. [Google Scholar] [CrossRef]
- Tetreau, G.; Sawaya, M.R.; De Zitter, E.; Andreeva, E.A.; Banneville, A.-S.; Schibrowsky, N.A.; Coquelle, N.; Brewster, A.S.; Grünbein, M.L.; Kovacs, G.N.; et al. De novo determination of mosquitocidal Cry11Aa and Cry11Ba structures from naturally-occurring nanocrystals. Nat. Commun. 2022, 13, 4376. [Google Scholar] [CrossRef]
- De Maagd, R.A.; Kwa, M.S.G.; Van Klei, H.D.; Yamamoto, T.; Schipper, B.; Vlak, J.M.; Stiekema, W.J.; Bosch, D. Domain III substitution in Bacillus thuringiensis delta-endotoxin CryIA(b) results in superior toxicity for Spodoptera exigua and altered membrane protein recognition. Appl. Environ. Microbiol. 1996, 62, 1537–1543. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, S.; Gómez, I.; Peláez-Aguilar, A.E.; Verduzco-Rosas, L.A.; García-Suárez, R.; do Nascimento, N.A.; Rivera-Nájera, L.Y.; Cantón, P.E.; Soberón, M.; Bravo, A. Structural changes upon membrane insertion of the insecticidal pore-forming toxins produced by Bacillus thuringiensis. Front. Insect Sci. 2023, 3, 1188891. [Google Scholar] [CrossRef]
- Knowles, B.H. Mechanism of Action of Bacillus thuringiensis Insecticidal δ-Endotoxins. Adv. Insect Physiol. 1994, 24, 275–308. [Google Scholar]
- Gazit, E.; La Rocca, P.; Sansom, M.S.P.; Shai, Y. The structure and organization within the membrane of the helices composing the pore-forming domain of Bacillus thuringiensis delta-endotoxin are consistent with an “umbrella-like” structure of the pore. Proc. Natl. Acad. Sci. USA 1998, 95, 12289–12294. [Google Scholar] [CrossRef] [PubMed]
- Gerber, D.; Shai, Y. Insertion and organization within membranes of the delta-endotoxin pore-forming domain, helix 4-loop-helix 5, and inhibition of its activity by a mutant helix 4 peptide. J. Biol. Chem. 2000, 275, 23602–23607. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, J.L.; Juteau, M.; Grochulski, P.; Cygler, M.; Prefontaine, G.; Brousseau, R.; Masson, L. Restriction of intramolecular movements within the Cry1Aa toxin molecule of Bacillus thuringiensis through disulfide bond engineering. FEBS Lett. 1997, 410, 397–402. [Google Scholar] [CrossRef]
- Masson, L.; Tabashnik, B.E.; Yong-Biao, L.; Brousseau, R.; Schwartz, J.-L. Helix 4 of the Bacillus thuringiensis Cry1Aa toxin lines the lumen of the ion channel. J. Biol. Chem. 1999, 274, 31996–32000. [Google Scholar] [CrossRef] [PubMed]
- Vachon, V.; Prefontaine, G.; Rang, C.; Coux, F.; Juteau, M.; Schwartz, J.L.; Brousseau, R.; Frutos, R.; Laprade, R.; Masson, L. Helix 4 mutants of the Bacillus thuringiensis insecticidal toxin Cry1Aa display altered pore-forming abilities. Appl. Environ. Microbiol. 2004, 70, 6123–6130. [Google Scholar] [CrossRef] [PubMed]
- Schnepf, E.; Crickmore, N.; Van Rie, J.; Lereclus, D.; Baum, J.; Feitelson, J.; Zeigler, D.R.; Dean, D.H. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 775–806. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Aronson, A.I. Localized Mutagenesis Defines Regions of the Bacillus-Thuringiensis Delta-Endotoxin Involved in Toxicity and Specificity. J. Biol. Chem. 1992, 267, 2311–2317. [Google Scholar] [CrossRef]
- Aronson, A.I.; Wu, D.; Zhang, C. Mutagenesis of specificity and toxicity regions of a Bacillus thuringiensis protoxin gene. J. Bacteriol. 1995, 177, 4059–4065. [Google Scholar] [CrossRef]
- Aronson, A.I.; Geng, C.X.; Wu, L. Aggregation of Bacillus thuringiensis Cry1A toxins upon binding to target insect larval midgut vesicles. Appl. Environ. Microbiol. 1999, 65, 2503–2507. [Google Scholar] [CrossRef]
- Kumar, A.S.M.; Aronson, A.I. Analysis of mutations in the pore-forming region essential for insecticidal activity of a Bacillus thuringiensis delta-endotoxin. J. Bacteriol. 1999, 181, 6103–6107. [Google Scholar] [CrossRef]
- Alzate, O.; You, T.; Claybon, M.; Osorio, C.; Curtiss, A.; Dean, D.H. Effects of disulfide bridges in domain I of Bacillus thuringiensis Cry1Aa δ-endotoxin on ion-channel formation in biological membranes. Biochemistry 2006, 45, 13597–13605. [Google Scholar] [CrossRef]
- Nair, M.S.; Dean, D.H. All domains of Cry1A toxins insert into insect brush border membranes. J. Biol. Chem. 2008, 283, 26324–26331. [Google Scholar] [CrossRef]
- Loseva, O.I.; Tiktopulo, E.I.; Vasiliev, V.D.; Nikulin, A.D.; Dobritsa, A.P.; Potekhin, S.A. Structure of Cry3A δ-endotoxin within phospholipid membranes. Biochemistry 2001, 40, 14143–14151. [Google Scholar] [CrossRef] [PubMed]
- Groulx, N.; Juteau, M.; Blunck, R. Rapid topology probing using fluorescence spectroscopy in planar lipid bilayer: The pore-forming mechanism of the toxin Cry1Aa of Bacillus thuringiensis. J. Gen. Physiol. 2010, 136, 497–513. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, S.; Quiliche, J.P.J.; Gómez, I.; Sánchez, J.; Soberón, M.; Bravo, A. Rearrangement of N-terminal α-helices of bacillus thuringiensis Cry1Ab toxin essential for oligomer assembly and toxicity. Toxins 2020, 12, 647. [Google Scholar] [CrossRef]
- Pacheco, S.; Gómez, I.; Sánchez, J.; García-Gómez, B.I.; Czajkowsky, D.M.; Zhang, J.; Soberón, M.; Bravo, A. Helix α-3 inter-molecular salt bridges and conformational changes are essential for toxicity of Bacillus thuringiensis 3D-Cry toxin family. Sci. Rep. 2018, 8, 10331. [Google Scholar] [CrossRef]
- Pacheco, S.; Gómez, I.; Soberón, M.; Bravo, A. A major conformational change of N-terminal helices of Bacillus thuringiensis Cry1Ab insecticidal protein is necessary for membrane insertion and toxicity. FEBS J. 2023, 290, 2692–2705. [Google Scholar] [CrossRef] [PubMed]
- Gómez, I.; Sánchez, J.; Muñoz-Garay, C.; Matus, V.; Gill, S.S.; Soberón, M.; Bravo, A. Bacillus thuringiensis Cry1A toxins are versatile proteins with multiple modes of action: Two distinct pre-pores are involved in toxicity. Biochem. J. 2014, 459, 383–396. [Google Scholar] [CrossRef]
- Khorramnejad, A.; Domínguez-Arrizabalaga, M.; Caballero, P.; Escriche, B.; Bel, Y. Study of the Bacillus thuringiensis Cry1Ia Protein Oligomerization Promoted by Midgut Brush Border Membrane Vesicles of Lepidopteran and Coleopteran Insects, or Cultured Insect Cells. Toxins 2020, 12, 133. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, S.; Gomez, I.; Gill, S.S.; Bravo, A.; Soberon, M. Enhancement of insecticidal activity of Bacillus thuringiensis Cry1A toxins by fragments of a toxin-binding cadherin correlates with oligomer formation. Peptides 2009, 30, 583–588. [Google Scholar] [CrossRef]
- Arenas, I.; Bravo, A.; Soberón, M.; Gómez, I. Role of alkaline phosphatase from Manduca sexta in the mechanism of action of Bacillus thuringiensis Cry1Ab toxin. J. Biol. Chem. 2010, 285, 12497–12503. [Google Scholar] [CrossRef]
- Obata, F.; Kitami, M.; Inoue, Y.; Atsumi, S.; Yoshizawa, Y.; Sato, R. Analysis of the region for receptor binding and triggering of oligomerization on Bacillus thuringiensis Cry1Aa toxin. FEBS J. 2009, 276, 5949–5959. [Google Scholar] [CrossRef]
- Vie, V.; Van Mau, N.; Pomarede, P.; Dance, C.; Schwartz, J.L.; Laprade, R.; Frutos, R.; Rang, C.; Masson, L.; Heitz, F.; et al. Lipid-induced pore formation of the Bacillus thuringiensis Cry1Aa insecticidal toxin. J. Membr. Biol. 2001, 180, 195–203. [Google Scholar] [CrossRef]
- Sriwimol, W.; Aroonkesorn, A.; Sakdee, S.; Kanchanawarin, C.; Uchihashi, T.; Ando, T.; Angsuthanasombat, C. Potential prepore trimer formation by the Bacillus thuringiensis mosquito-specific toxin: Molecular insights into a critical prerequisite of membrane-bound monomers. J. Biol. Chem. 2015, 290, 20793–20803. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Curtiss, A.; Alcantara, E.; Dean, D.H. Mutations in Domain I of Bacillus thuringiensis δ-Endotoxin CryIAb Reduce the Irreversible Binding of Toxin to Manduca sexta Brush Border Membrane Vesicles. J. Biol. Chem. 1995, 270, 6412–6419. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.R.; Aronson, A.I.; Dean, D.H. Substitution of residues on the proximal side of Cry1A Bacillus thuringiensis delta-endotoxins affects irreversible binding to Manduca sexta midgut membrane. Biochem. Biophys. Res. Commun. 1996, 226, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Groulx, N.; McGuire, H.; Laprade, R.; Schwartz, J.L.; Blunck, R. Single Molecule Fluorescence Study of the Bacillus thuringiensis Toxin Cry1Aa Reveals Tetramerization. J. Biol. Chem. 2011, 286, 42274–42282. [Google Scholar] [CrossRef] [PubMed]
- Tigue, N.J.; Jacoby, J.; Ellar, D.J. The alpha-helix 4 residue, Asn135, is involved in the oligomerization of Cry1Ac1 and Cry1Ab5 Bacillus thuringiensis toxins. Appl. Environ. Microbiol. 2001, 67, 5715–5720. [Google Scholar] [CrossRef]
- Likitvivatanavong, S.; Katzenmeier, G.; Angsuthanasombat, C. Asn183 in a5 is essential for oligomerisation and toxicity of the Bacillus thuringiensis Cry4Ba toxin. Arch. Biochem. Biophys. 2006, 445, 46–55. [Google Scholar] [CrossRef]
- Knowles, B.H.; Ellar, D.J. Colloid-osmotic lysis is a general feature of the mechanism of action of Bacillus thuringiensis d-endotoxins with different insect specificity. Biochim. Biophys. Acta-Gen. Subj. 1987, 924, 509–518. [Google Scholar] [CrossRef]
- Peyronnet, O.; Nieman, B.; Genereux, F.; Vachon, V.; Laprade, R.; Schwartz, J.L. Estimation of the radius of the pores formed by the Bacillus thuringiensis Cry1C delta-endotoxin in planar lipid bilayers. Biochim. Biophys. Acta 2002, 1567, 113–122. [Google Scholar] [CrossRef]
- Ounjai, P.; Unger, V.M.; Sigworth, F.J.; Angsuthanasombat, C. Two conformational states of the membrane-associated Bacillus thuringiensis Cry4Ba delta-endotoxin complex revealed by electron crystallography: Implications for toxin-pore formation. Biochem. Biophys. Res. Commun. 2007, 361, 890–895. [Google Scholar] [CrossRef]
- Mirdita, M.; Schutze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making protein folding accessible to all. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, S.; Gómez, I.; Sánchez, J.; García-Gómez, B.I.; Soberón, M.; Bravo, A. An intramolecular salt bridge in Bacillus thuringiensis Cry4Ba toxin is involved in the stability of helix α-3, which is needed for oligomerization and insecticidal activity. Appl. Environ. Microbiol. 2017, 83, e01515-17. [Google Scholar] [CrossRef] [PubMed]
- Puntheeranurak, T.; Stroh, C.; Zhu, R.; Angsuthanasombat, C.; Hinterdorfer, P. Structure and distribution of the Bacillus thuringiensis Cry4Ba toxin in lipid membranes. Ultramicroscopy 2005, 105, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Mariani, V.; Biasini, M.; Barbato, A.; Schwede, T. lDDT: A local superposition-free score for comparing protein structures and models using distance difference tests. Bioinformatics 2013, 29, 2722–2728. [Google Scholar] [CrossRef] [PubMed]
- Tunyasuvunakool, K.; Adler, J.; Wu, Z.; Green, T.; Zielinski, M.; Zidek, A.; Bridgland, A.; Cowie, A.; Meyer, C.; Laydon, A.; et al. Highly accurate protein structure prediction for the human proteome. Nature 2021, 596, 590–596. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef]
- Rausell, C.; Pardo-Lopez, L.; Sanchez, J.; Munoz-Garay, C.; Morera, C.; Soberon, M.; Bravo, A. Unfolding events in the water-soluble monomeric Cry1Ab toxin during transition to oligomeric pre-pore and membrane-inserted pore channel. J. Biol. Chem. 2004, 279, 55168–55175. [Google Scholar] [CrossRef]
- Alcaraz, A.; Ramírez, P.; García-Giménez, E.; López, M.L.; Andrio, A.; Aguilella, V.M. A pH-Tunable Nanofluidic Diode: Electrochemical Rectification in a Reconstituted Single Ion Channel. J. Phys. Chem. B 2006, 110, 21205–21209. [Google Scholar] [CrossRef]
- Alcaraz, A.; Nestorovich, E.M.; Aguilella-Arzo, M.; Aguilella, V.M.; Bezrukov, S.M. Salting Out the Ionic Selectivity of a Wide Channel: The Asymmetry of OmpF. Biophys. J. 2004, 87, 943–957. [Google Scholar] [CrossRef]
- Nestorovich, E.M.; Rostovtseva, T.K.; Bezrukov, S.M. Residue Ionization and Ion Transport through OmpF Channels. Biophys. J. 2003, 85, 3718–3729. [Google Scholar] [CrossRef] [PubMed]
- McBride, J.M.; Polev, K.; Abdirasulov, A.; Reinharz, V.; Grzybowski, B.A.; Tlusty, T. AlphaFold2 Can Predict Single-Mutation Effects. Phys. Rev. Lett. 2023, 131, 218401. [Google Scholar] [CrossRef]
- Bryant, P.; Pozzati, G.; Elofsson, A. Improved prediction of protein-protein interactions using AlphaFold2. Nat. Commun. 2022, 13, 1265. [Google Scholar] [CrossRef] [PubMed]
- Hegedus, T.; Geisler, M.; Lukacs, G.L.; Farkas, B. Ins and outs of AlphaFold2 transmembrane protein structure predictions. Cell Mol. Life Sci. 2022, 79, 73. [Google Scholar] [CrossRef]
- Varadi, M.; Bordin, N.; Orengo, C.; Velankar, S. The opportunities and challenges posed by the new generation of deep learning-based protein structure predictors. Curr. Opin. Struct. Biol. 2023, 79, 102543. [Google Scholar] [CrossRef]
- Girard, F.; Vachon, V.; Préfontaine, G.; Marceau, L.; Su, Y.; Larouche, G.; Vincent, C.; Schwartz, J.L.; Masson, L.; Laprade, R. Cysteine scanning mutagenesis of α4, a putative pore-lining helix of the Bacillus thunngiensis insecticidal toxin Cry1Aa. Appl. Environ. Microbiol. 2008, 74, 2565–2572. [Google Scholar] [CrossRef]
- Vachon, V.; Préfontaine, G.; Coux, F.; Rang, C.; Marceau, L.; Masson, L.; Brousseau, R.; Frutos, R.; Schwartz, J.L.; Laprade, R. Role of helix 3 in pore formation by the Bacillus thuringiensis insecticidal toxin Cry1Aa. Biochemistry 2002, 41, 6178–6184. [Google Scholar] [CrossRef]
- Jimenez-Juarez, N.; Munoz-Garay, C.; Gomez, I.; Saab-Rincon, G.; Damian-Almazo, J.Y.; Gill, S.S.; Soberon, M.; Bravo, A. Bacillus thuringiensis Cry1Ab mutants affecting oligomer formation are non-toxic to Manduca sexta larvae. J. Biol. Chem. 2007, 282, 21222–21229. [Google Scholar] [CrossRef]
- Lin, X.; Parthasarathy, K.; Surya, W.; Zhang, T.; Mu, Y.; Torres, J. A conserved tetrameric interaction of cry toxin helix α3 suggests a functional role for toxin oligomerization. Biochim. Biophys. Acta 2014, 1838, 1777–1784. [Google Scholar] [CrossRef] [PubMed]
- Kirouac, M.; Vachon, V.; Noel, J.F.; Girard, F.; Schwartz, J.L.; Laprade, R. Amino acid and divalent ion permeability of the pores formed by the Bacillus thuringiensis toxins Cry1Aa and Cry1Ac in insect midgut brush border membrane vesicles. Biochim. Biophys. Acta 2002, 1561, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Soberón, M.; Pardo-López, L.; López, I.; Gómez, I.; Tabashnik, B.E.; Bravo, A. Engineering modified Bt toxins to counter insect resistance. Science 2007, 318, 1640–1642. [Google Scholar] [CrossRef]
- Muñóz-Garay, C.; Portugal, L.; Pardo-López, L.; Jiménez-Juárez, N.; Arenas, I.; Gómez, I.; Sánchez-López, R.; Arroyo, R.; Holzenburg, A.; Savva, C.G.; et al. Characterization of the mechanism of action of the genetically modified Cry1AbMod toxin that is active against Cry1Ab-resistant insects. Biochim. Biophys. Acta-Biomembr. 2009, 1788, 2229–2237. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Garay, C.; Rodríguez-Almazán, C.; Aguilar, J.N.; Portugal, L.; Gómez, I.; Saab-Rincon, G.; Soberón, M.; Bravo, A. Oligomerization of Cry11Aa from Bacillus thuringiensis has an important role in toxicity against Aedes aegypti. Appl. Environ. Microbiol. 2009, 75, 7548–7550. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.; Curtiss, A.; Dean, D.H.; Alzate, O. The role of a proline-induced broken-helix motif in α-helix 2 of Bacillus thuringiensis δ-endotoxins. FEBS Lett. 2001, 490, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Ramírez, R.; Huesa, J.; Bel, Y.; Ferré, J.; Casino, P.; Arias-Palomo, E. Molecular architecture and activation of the insecticidal protein Vip3Aa from Bacillus thuringiensis. Nat. Commun. 2020, 11, 3974. [Google Scholar] [CrossRef] [PubMed]
- Byrne, M.J.; Iadanza, M.G.; Perez, M.A.; Maskell, D.P.; George, R.M.; Hesketh, E.L.; Beales, P.A.; Zack, M.D.; Berry, C.; Thompson, R.F. Cryo-EM structures of an insecticidal Bt toxin reveal its mechanism of action on the membrane. Nat. Commun. 2021, 12, 2791. [Google Scholar] [CrossRef] [PubMed]
- Mueller, M.; Grauschopf, U.; Maier, T.; Glockshuber, R.; Ban, N. The structure of a cytolytic α-helical toxin pore reveals its assembly mechanism. Nature 2009, 459, 726–730. [Google Scholar] [CrossRef]
- Gatsogiannis, C.; Merino, F.; Prumbaum, D.; Roderer, D.; Leidreiter, F.; Meusch, D.; Raunser, S. Membrane insertion of a Tc toxin in near-atomic detail. Nat. Struct. Mol. Biol. 2016, 23, 884–890. [Google Scholar] [CrossRef]
- Tomimoto, K.; Hayakawa, T.; Hori, H. Pronase digestion of brush border membrane-bound Cry1Aa shows that almost the whole activated Cry1Aa molecule penetrates into the membrane. Comp. Biochem. Physiol.-B Biochem. Mol. Biol. 2006, 144, 413–422. [Google Scholar] [CrossRef]
- Zavala, L.E.; Pardo-López, L.; Cantón, P.E.; Gómez, I.; Soberón, M.; Bravo, A. Domains II and III of Bacillus thuringiensis Cry1Ab Toxin remain exposed to the solvent after insertion of part of domain I into the membrane. J. Biol. Chem. 2011, 286, 19109–19117. [Google Scholar] [CrossRef]
- Nunez-Valdez, M.E.; Sanchez, J.; Lina, L.; Guereca, L.; Bravo, A. Structural and functional studies of alpha-helix 5 region from Bacillus thuringiensis Cry1Ab delta-endotoxin. Biochim. Biophys. Acta-Protein Struct. Mol. Enzymol. 2001, 1546, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, J.-L.; Garneau, L.; Savaria, D.; Masson, L.; Brousseau, R.; Rousseau, E. Lepidopteran-specific crystal toxins from Bacillus thuringiensis form cation- and anion-selective channels in planar lipid bilayers. J. Membr. Biol. 1993, 132, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Tetreau, G.; Banneville, A.S.; Andreeva, E.A.; Brewster, A.S.; Hunter, M.S.; Sierra, R.G.; Teulon, J.M.; Young, I.D.; Burke, N.; Grünewald, T.A.; et al. Serial femtosecond crystallography on in vivo-grown crystals drives elucidation of mosquitocidal Cyt1Aa bioactivation cascade. Nat. Commun. 2020, 11, 1153. [Google Scholar] [CrossRef] [PubMed]
- Evans, R.; O’Neill, M.; Pritzel, A.; Antropova, N.; Senior, A.; Green, T.; Žídek, A.; Bates, R.; Blackwell, S.; Yim, J.; et al. Protein complex prediction with AlphaFold-Multimer. bioRxiv 2022. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, Y. Improving the physical realism and structural accuracy of protein models by a two-step atomic-level energy minimization. Biophys. J. 2011, 101, 2525–2534. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Smart, O.S.; Neduvelil, J.G.; Wang, X.; Wallace, B.A.; Sansom, M.S. HOLE: A program for the analysis of the pore dimensions of ion channel structural models. J. Mol. Graph. 1996, 14, 354–360, 376. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres, J.; Surya, W.; Boonserm, P. Channel Formation in Cry Toxins: An Alphafold-2 Perspective. Int. J. Mol. Sci. 2023, 24, 16809. https://doi.org/10.3390/ijms242316809
Torres J, Surya W, Boonserm P. Channel Formation in Cry Toxins: An Alphafold-2 Perspective. International Journal of Molecular Sciences. 2023; 24(23):16809. https://doi.org/10.3390/ijms242316809
Chicago/Turabian StyleTorres, Jaume, Wahyu Surya, and Panadda Boonserm. 2023. "Channel Formation in Cry Toxins: An Alphafold-2 Perspective" International Journal of Molecular Sciences 24, no. 23: 16809. https://doi.org/10.3390/ijms242316809
APA StyleTorres, J., Surya, W., & Boonserm, P. (2023). Channel Formation in Cry Toxins: An Alphafold-2 Perspective. International Journal of Molecular Sciences, 24(23), 16809. https://doi.org/10.3390/ijms242316809