

Recognition of Differentially Expressed Molecular Signatures and Pathways Associated with COVID-19 Poor Prognosis in Glioblastoma Patients


Abstract
:1. Introduction
2. Results
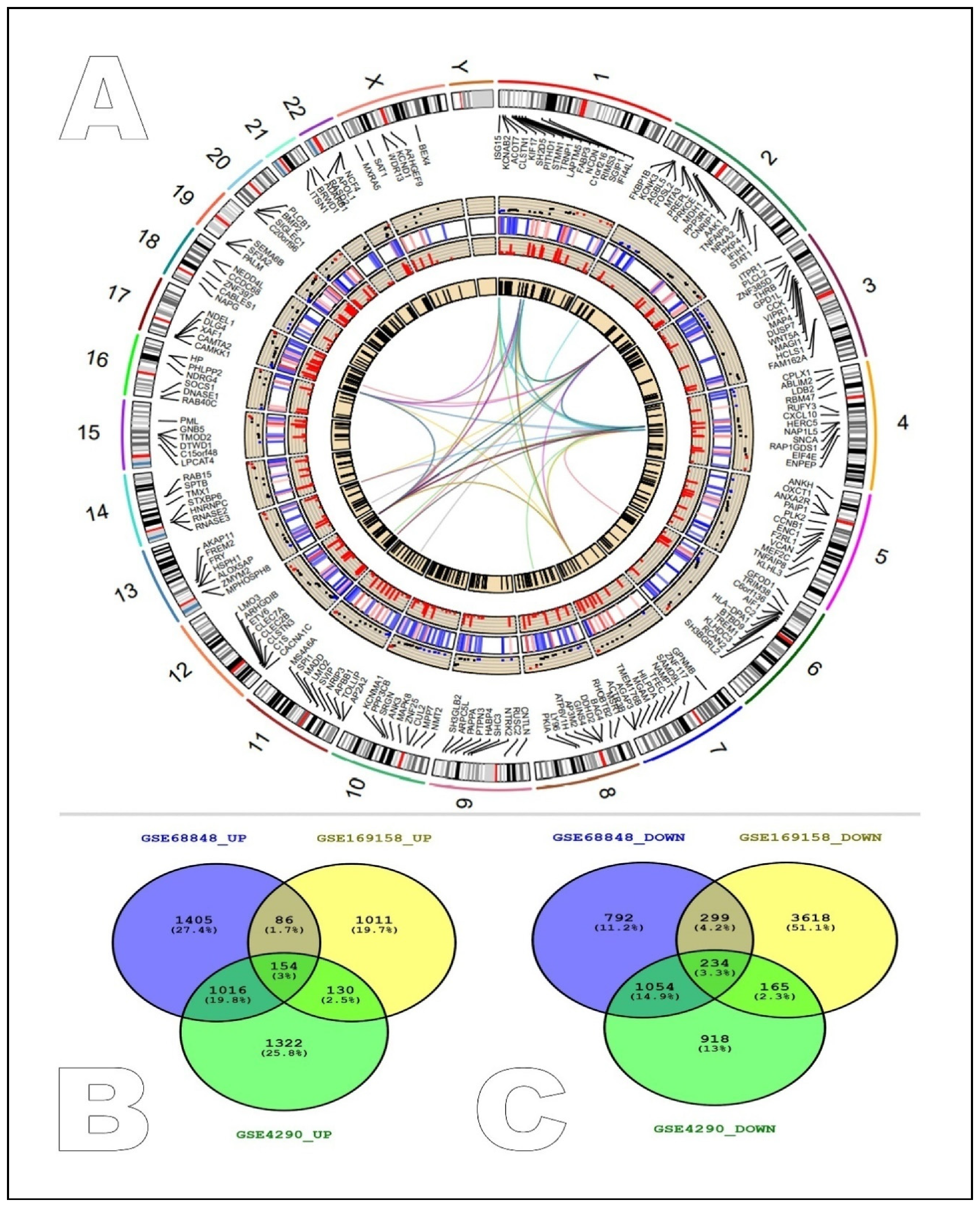
2.1. Identification of Common Differentially Expressed Genes between GEO Studies
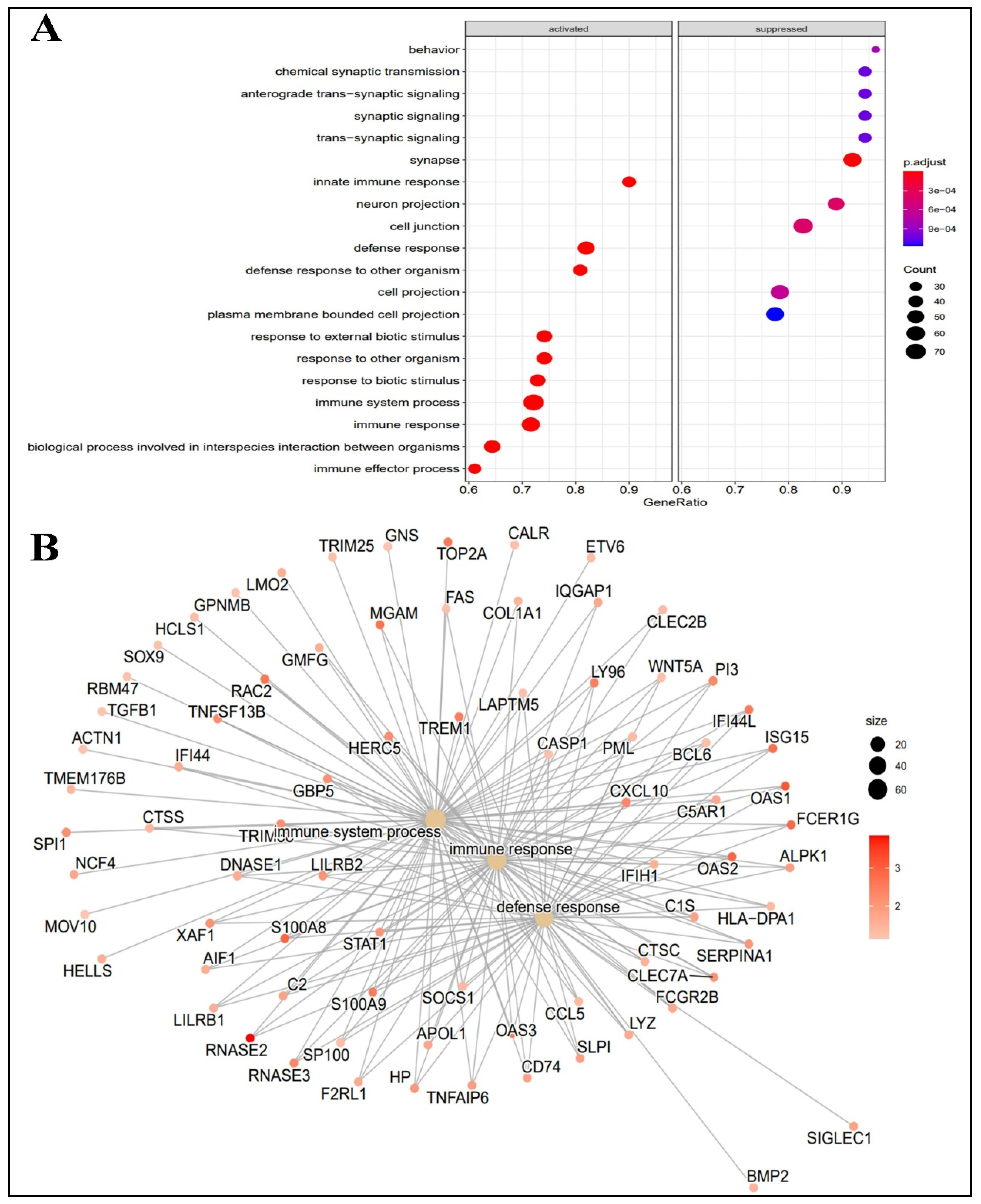
2.2. Functional Analysis of Differentially Expressed Genes
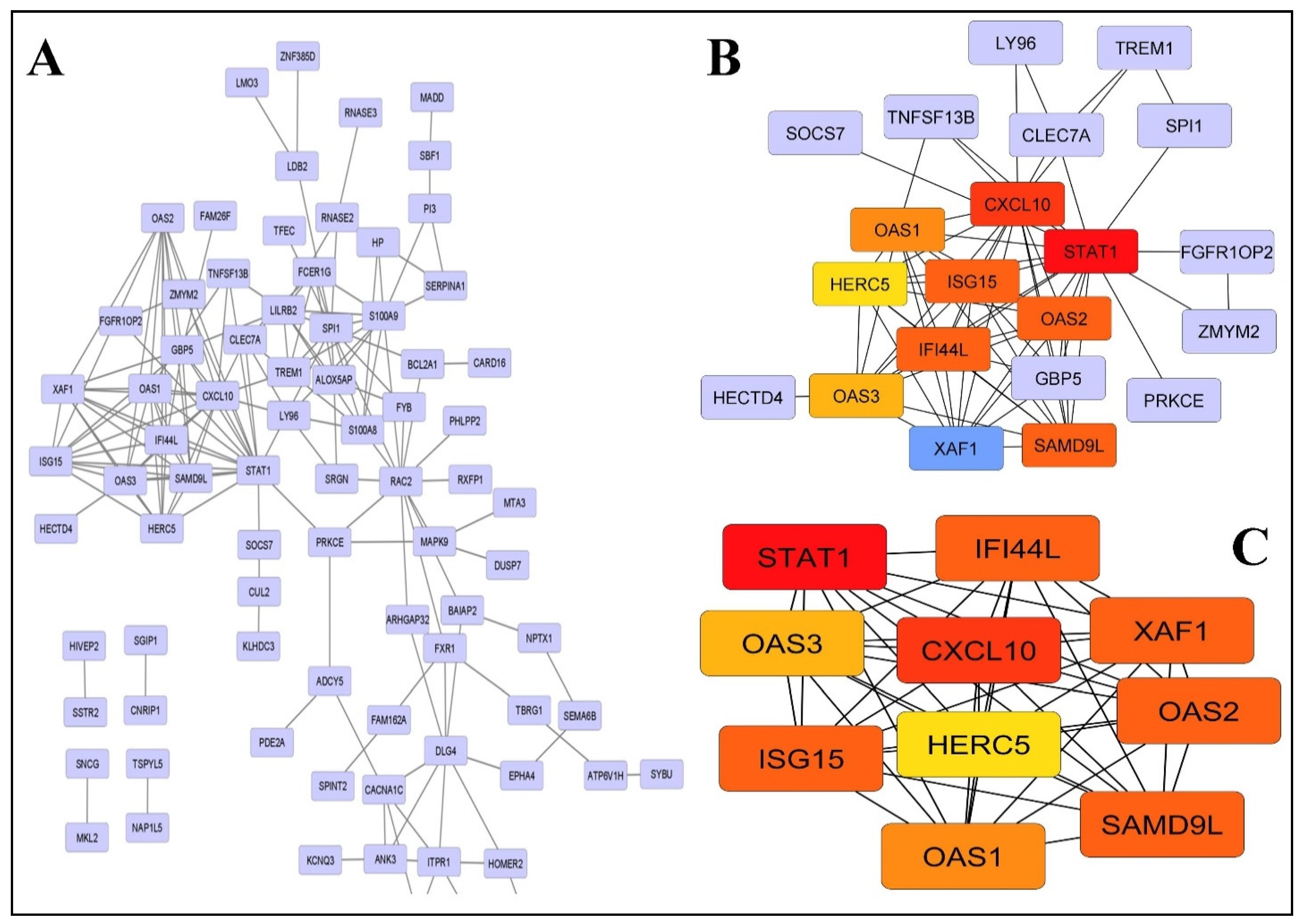
2.3. PPI Network Construction and Modules Selection
2.4. cMAP Analysis
2.5. Statistical Analysis
3. Discussion
4. Materials and Methods
4.1. Data Collection and Processing
4.2. Enrichment Analysis of Genes and Pathways of Differentially Expressed Genes
4.3. Construction of PPI Network and Selection of Hub Genes
4.4. Connectivity Map (cMAP)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| cMAP | connectivity map |
| overexpressed | up-regulated |
| low-expressed | down-regulated |
| DEGs | differentially expressed genes |
| GBM | glioblastoma |
| GEO | gene expression omnibus |
| GO | gene ontology |
| PPI | protein–protein interaction |
| KEGG | Kyoto encyclopedia of genes and genomes |
| Module | group of selected hub genes |
References
- Hanif, F.; Muzaffar, K.; Perveen, K.; Malhi, S.M.; Simjee Sh, U. Glioblastoma Multiforme: A Review of its Epidemiology and Pathogenesis through Clinical Presentation and Treatment. Asian Pac. J. Cancer Prev. APJCP 2017, 18, 3–9. [Google Scholar] [CrossRef]
- Mihelson, N.; McGavern, D.B. Viral Control of Glioblastoma. Viruses 2021, 13, 1264. [Google Scholar] [CrossRef]
- Wollmann, G.; Ozduman, K.; van den Pol, A.N. Oncolytic virus therapy for glioblastoma multiforme: Concepts and candidates. Cancer J. 2012, 18, 69–81. [Google Scholar] [CrossRef]
- Pappula, A.L.; Rasheed, S.; Mirzaei, G.; Petreaca, R.C.; Bouley, R.A. A Genome-Wide Profiling of Glioma Patients with an IDH1 Mutation Using the Catalogue of Somatic Mutations in Cancer Database. Cancers 2021, 13, 4299. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.-L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Rong, L.; Li, N.; Zhang, Z. Emerging therapies for glioblastoma: Current state and future directions. J. Exp. Clin. Cancer Res. 2022, 41, 1–18. [Google Scholar] [CrossRef]
- Cai, Z.; Yang, S.; Li, X.; Chen, F.; Li, W. Viral infection and glioma: A meta-analysis of prognosis. BMC Cancer 2020, 20, 549. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Li, M.; Guo, Y.; Zhong, Y.; He, Z.; Xu, Y.; Zou, J. Immune response in glioma’s microenvironment. Innov. Surg. Sci. 2020, 5, 20190001. [Google Scholar] [CrossRef]
- Novellino, F.; Sacca, V.; Donato, A.; Zaffino, P.; Spadea, M.F.; Vismara, M.; Arcidiacono, B.; Malara, N.; Presta, I.; Donato, G. Innate Immunity: A Common Denominator between Neurodegenerative and Neuropsychiatric Diseases. Int. J. Mol. Sci. 2020, 21, 1115. [Google Scholar] [CrossRef]
- Sami, R.; Fathi, F.; Eskandari, N.; Ahmadi, M.; ArefNezhad, R.; Motedayyen, H. Characterizing the immune responses of those who survived or succumbed to COVID-19: Can immunological signatures predict outcome? Cytokine 2021, 140, 155439. [Google Scholar] [CrossRef]
- Peyneau, M.; Granger, V.; Wicky, P.H.; Khelifi-Touhami, D.; Timsit, J.F.; Lescure, F.X.; Yazdanpanah, Y.; Tran-Dinh, A.; Montravers, P.; Monteiro, R.C.; et al. Innate immune deficiencies are associated with severity and poor prognosis in patients with COVID-19. Sci. Rep. 2022, 12, 638. [Google Scholar] [CrossRef]
- Ansari, N.; Jahangiri, M.; Shirbandi, K.; Ebrahimi, M.; Rahim, F. The association between different predictive biomarkers and mortality of COVID-19. Bull. Natl. Res. Cent. 2022, 46, 158. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.J.A.; Ribeiro, L.R.; Lima, K.V.B.; Lima, L. Adaptive immunity to SARS-CoV-2 infection: A systematic review. Front. Immunol. 2022, 13, 1001198. [Google Scholar] [CrossRef] [PubMed]
- Kofman, A.; Marcinkiewicz, L.; Dupart, E.; Lyshchev, A.; Martynov, B.; Ryndin, A.; Kotelevskaya, E.; Brown, J.; Schiff, D.; Abounader, R. The roles of viruses in brain tumor initiation and oncomodulation. J. Neuro-Oncol. 2011, 105, 451–466. [Google Scholar] [CrossRef]
- Lee, H.Y.; Kim, H.S.; Park, J.W.; Baek, H.J.; Kim, S.J.; Choi, C.G. Atypical imaging features of Epstein-Barr virus-positive primary central nervous system lymphomas in patients without AIDS. AJNR. Am. J. Neuroradiol. 2013, 34, 1562–1567. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.F.; Zhang, Q.; Chen, Z.Y.; Wang, H.F.; Li, X.; Wang, H.X.; Li, H.X.; Kang, C.M.; Chu, S.; Li, K.F.; et al. NLRP3 in human glioma is correlated with increased WHO grade, and regulates cellular proliferation, apoptosis and metastasis via epithelial-mesenchymal transition and the PTEN/AKT signaling pathway. Int. J. Oncol. 2018, 53, 973–986. [Google Scholar] [CrossRef]
- Lim, J.; Kim, M.J.; Park, Y.; Ahn, J.W.; Hwang, S.J.; Moon, J.S.; Cho, K.G.; Kwack, K. Upregulation of the NLRC4 inflammasome contributes to poor prognosis in glioma patients. Sci. Rep. 2019, 9, 7895. [Google Scholar] [CrossRef]
- Iwadate, Y. Epithelial-mesenchymal transition in glioblastoma progression. Oncol. Lett. 2016, 11, 1615–1620. [Google Scholar] [CrossRef]
- Ribatti, D.; Tamma, R.; Annese, T. Epithelial-Mesenchymal Transition in Cancer: A Historical Overview. Transl. Oncol. 2020, 13, 100773. [Google Scholar] [CrossRef]
- Lai, X.; Li, Q.; Wu, F.; Lin, J.; Chen, J.; Zheng, H.; Guo, L. Epithelial-Mesenchymal Transition and Metabolic Switching in Cancer: Lessons From Somatic Cell Reprogramming. Front. Cell Dev. Biol. 2020, 8, 760. [Google Scholar] [CrossRef]
- Pan, P.; Shen, M.; Yu, Z.; Ge, W.; Chen, K.; Tian, M.; Xiao, F.; Wang, Z.; Wang, J.; Jia, Y.; et al. Author Correction: SARS-CoV-2 N protein promotes NLRP3 inflammasome activation to induce hyperinflammation. Nat. Commun. 2021, 12, 5306. [Google Scholar] [CrossRef]
- Zhu, C.; Zou, C.; Guan, G.; Guo, Q.; Yan, Z.; Liu, T.; Shen, S.; Xu, X.; Chen, C.; Lin, Z.; et al. Development and validation of an interferon signature predicting prognosis and treatment response for glioblastoma. Oncoimmunology 2019, 8, e1621677. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, X.; Chen, X.; Zhang, Q.; Hong, J. Novel Immune-Related Gene Signature for Risk Stratification and Prognosis of Survival in Lower-Grade Glioma. Front. Genet. 2020, 11, 363. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Wu, S.; Chen, X.; Ye, Y.; Weng, Y.; Pan, Y.; Chen, Z.; Chen, L.; Qiu, X.; Qiu, S. Characterization of Hypoxia Signature to Evaluate the Tumor Immune Microenvironment and Predict Prognosis in Glioma Groups. Front. Oncol. 2020, 10, 796. [Google Scholar] [CrossRef] [PubMed]
- Vanderheiden, A.; Ralfs, P.; Chirkova, T.; Upadhyay, A.A.; Zimmerman, M.G.; Bedoya, S.; Aoued, H.; Tharp, G.M.; Pellegrini, K.L.; Manfredi, C.; et al. Type I and Type III Interferons Restrict SARS-CoV-2 Infection of Human Airway Epithelial Cultures. J. Virol. 2020, 94. [Google Scholar] [CrossRef]
- McGonagle, D.; Ramanan, A.V.; Bridgewood, C. Immune cartography of macrophage activation syndrome in the COVID-19 era. Nat. Rev. Rheumatol. 2021, 17, 145–157. [Google Scholar] [CrossRef]
- Perlman, S. COVID-19 poses a riddle for the immune system. Nature 2020, 584, 345–346. [Google Scholar] [CrossRef]
- Salman, A.A.; Waheed, M.H.; Ali-Abdulsahib, A.A.; Atwan, Z.W. Low type I interferon response in COVID-19 patients: Interferon response may be a potential treatment for COVID-19. Biomed. Rep. 2021, 14, 43. [Google Scholar] [CrossRef]
- Zhu, H.; Li, J.T.; Zheng, F.; Martin, E.; Kots, A.Y.; Krumenacker, J.S.; Choi, B.K.; McCutcheon, I.E.; Weisbrodt, N.; Bögler, O.; et al. Restoring soluble guanylyl cyclase expression and function blocks the aggressive course of glioma. Mol. Pharmacol. 2011, 80, 1076–1084. [Google Scholar] [CrossRef]
- Paulus, W.J.; Tschöpe, C. A novel paradigm for heart failure with preserved ejection fraction: Comorbidities drive myocardial dysfunction and remodeling through coronary microvascular endothelial inflammation. J. Am. Coll. Cardiol. 2013, 62, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Portela, M.; Venkataramani, V.; Fahey-Lozano, N.; Seco, E.; Losada-Perez, M.; Winkler, F.; Casas-Tintó, S. Glioblastoma cells vampirize WNT from neurons and trigger a JNK/MMP signaling loop that enhances glioblastoma progression and neurodegeneration. PLoS Biol. 2019, 17, e3000545. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liao, Y.; Chen, B.; Chen, Y.; Yu, Z.; Wei, H.; Zhang, L.; Huang, S.; Rothman, P.B.; Gao, G.F.; et al. Critical role of Syk-dependent STAT1 activation in innate antiviral immunity. Cell Rep. 2021, 34, 108627. [Google Scholar] [CrossRef]
- Salazar-Mather, T.P.; Hamilton, T.A.; Biron, C.A. A chemokine-to-cytokine-to-chemokine cascade critical in antiviral defense. J. Clin. Investig. 2000, 105, 985–993. [Google Scholar] [CrossRef]
- Lee, M.G.; Han, J.; Jeong, S.I.; Her, N.G.; Lee, J.H.; Ha, T.K.; Kang, M.J.; Ryu, B.K.; Chi, S.G. XAF1 directs apoptotic switch of p53 signaling through activation of HIPK2 and ZNF313. Proc. Natl. Acad. Sci. USA 2014, 111, 15532–15537. [Google Scholar] [CrossRef]
- DeDiego, M.L.; Martinez-Sobrido, L.; Topham, D.J. Novel Functions of IFI44L as a Feedback Regulator of Host Antiviral Responses. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed]
- Silverman, R.H. Viral encounters with 2′,5′-oligoadenylate synthetase and RNase L during the interferon antiviral response. J. Virol. 2007, 81, 12720–12729. [Google Scholar] [CrossRef]
- Sampson, D.L.; Fox, B.A.; Yager, T.D.; Bhide, S.; Cermelli, S.; McHugh, L.C.; Seldon, T.A.; Brandon, R.A.; Sullivan, E.; Zimmerman, J.J.; et al. A Four-Biomarker Blood Signature Discriminates Systemic Inflammation Due to Viral Infection Versus Other Etiologies. Sci. Rep. 2017, 7, 2914. [Google Scholar] [CrossRef]
- Paparisto, E.; Woods, M.W.; Coleman, M.D.; Moghadasi, S.A.; Kochar, D.S.; Tom, S.K.; Kohio, H.P.; Gibson, R.M.; Rohringer, T.J.; Hunt, N.R.; et al. Evolution-Guided Structural and Functional Analyses of the HERC Family Reveal an Ancient Marine Origin and Determinants of Antiviral Activity. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed]
- Langhans, J.; Schneele, L.; Trenkler, N.; von Bandemer, H.; Nonnenmacher, L.; Karpel-Massler, G.; Siegelin, M.D.; Zhou, S.; Halatsch, M.E.; Debatin, K.M.; et al. The effects of PI3K-mediated signalling on glioblastoma cell behaviour. Oncogenesis 2017, 6, 398. [Google Scholar] [CrossRef]
- Chresta, C.M.; Davies, B.R.; Hickson, I.; Harding, T.; Cosulich, S.; Critchlow, S.E.; Vincent, J.P.; Ellston, R.; Jones, D.; Sini, P.; et al. AZD8055 is a potent, selective, and orally bioavailable ATP-competitive mammalian target of rapamycin kinase inhibitor with in vitro and in vivo antitumor activity. Cancer Res. 2010, 70, 288–298. [Google Scholar] [CrossRef] [Green Version]
- Le Sage, V.; Cinti, A.; Amorim, R.; Mouland, A.J. Adapting the Stress Response: Viral Subversion of the mTOR Signaling Pathway. Viruses 2016, 8, 152. [Google Scholar] [CrossRef]
- Okada, M.; Suzuki, S.; Togashi, K.; Sugai, A.; Yamamoto, M.; Kitanaka, C. Targeting Folate Metabolism Is Selectively Cytotoxic to Glioma Stem Cells and Effectively Cooperates with Differentiation Therapy to Eliminate Tumor-Initiating Cells in Glioma Xenografts. Int. J. Mol. Sci. 2021, 22, 11633. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, A.; Wiesmann, T.; Vogelmeier, C.F.; Mack, E.; Skevaki, C.; Gaik, C.; Keller, C.; Figiel, J.; Sohlbach, K.; Rolfes, C.; et al. Ruxolitinib for the treatment of SARS-CoV-2 induced acute respiratory distress syndrome (ARDS). Leukemia 2020, 34, 2276–2278. [Google Scholar] [CrossRef] [PubMed]
- Lim, N.; Pavlidis, P. Evaluation of connectivity map shows limited reproducibility in drug repositioning. Sci. Rep. 2021, 11, 17624. [Google Scholar] [CrossRef] [PubMed]
- Safavi, F.; Nath, A. Silencing of immune activation with methotrexate in patients with COVID-19. J. Neurol. Sci. 2020, 415, 116942. [Google Scholar] [CrossRef]
- Cao, Y.; Wei, J.; Zou, L.; Jiang, T.; Wang, G.; Chen, L.; Huang, L.; Meng, F.; Huang, L.; Wang, N.; et al. Ruxolitinib in treatment of severe coronavirus disease 2019 (COVID-19): A multicenter, single-blind, randomized controlled trial. J. Allergy Clin. Immunol. 2020, 146, 137–146.e133. [Google Scholar] [CrossRef]
- Rein, L.; Calero, K.; Shah, R.; Ojielo, C.; Hudock, K.M.; Lodhi, S.; Sadaka, F.; Bellam, S.; Palma, C.; Hager, D.N.; et al. Randomized Phase 3 Trial of Ruxolitinib for COVID-19-Associated Acute Respiratory Distress Syndrome. Crit. Care Med. 2022, 50, 1701–1713. [Google Scholar] [CrossRef]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets—Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Cech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Furumichi, M.; Tanabe, M.; Hirakawa, M. KEGG for representation and analysis of molecular networks involving diseases and drugs. Nucleic Acids Res. 2010, 38, D355–D360. [Google Scholar] [CrossRef] [Green Version]
- Reimand, J.; Kull, M.; Peterson, H.; Hansen, J.; Vilo, J. g:Profiler—A web-based toolset for functional profiling of gene lists from large-scale experiments. Nucleic Acids Res. 2007, 35, W193–W200. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Chin, C.H.; Chen, S.H.; Wu, H.H.; Ho, C.W.; Ko, M.T.; Lin, C.Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8 (Suppl. S4), S11. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| GO Category | Term Name | Genes Enriched | % | p-Value |
|---|---|---|---|---|
| MF | Positive regulation of 2′-5′-oligoadenylate synthetase activity | 3 | 2.1 | 1.25 × 10−3 |
| Positive regulation of Toll-like receptor 4 binding | 3 | 2.1 | 1.25 × 10−3 | |
| Positive regulation of arachidonic acid binding | 3 | 2.1 | 6.17 × 10−3 | |
| Negative regulation cytoskeleton protein binding | 32 | 14.55 | 1.00 × 10−4 | |
| Negative regulation of nucleoside-triphosphatase regulator activity | 22 | 10 | 1.10 × 10−4 | |
| BP | Immune system process | 76 | 57.58 | 1.26 × 10−19 |
| Immune response | 62 | 46.97 | 1.55 × 10−16 | |
| Defense response to other organisms | 44 | 33.33 | 1.88 × 10−16 | |
| Regulation of trans-synaptic signaling | 32 | 15.09 | 1.78 × 10−13 | |
| Chemical synaptic transmission | 38 | 17.92 | 7.34 × 10−12 | |
| CC | Cytoplasmic vesicle | 45 | 31.25 | 1.66 × 10−6 |
| Intracellular vesicle | 45 | 31.25 | 1.77 × 10−6 | |
| Secretory granule | 24 | 16.67 | 8.60 × 10−6 | |
| Synapse | 64 | 29.22 | 1.08 × 10−20 | |
| Cell junction | 71 | 32.42 | 6.38 × 10−15 |
| Pathway Source | Pathway Name | Genes Enriched | % | p-Value |
|---|---|---|---|---|
| KEGG | Influenza A | 13 | 14.94 | 2.35 × 10−6 |
| KEGG | Coronavirus disease—COVID-19 | 11 | 12.64 | 3.15 × 10−3 |
| KEGG | NOD-like receptor signaling pathway | 9 | 10.34 | 1.02 × 10−2 |
| KEGG | cGMP-PKG signaling pathway | 12 | 12.9 | 3.57 × 10−5 |
| KEGG | Growth hormone synthesis, secretion, and action | 10 | 10.75 | 8.91 × 10−5 |
| Reactome | Immune system | 56 | 54.9 | 3.22 × 10−13 |
| Reactome | Interferon signaling | 15 | 14.71 | 1.60 × 10−7 |
| Reactome | Innate immune system | 33 | 32.35 | 3.53 × 10−7 |
| Reactome | Neuronal system | 18 | 14.06 | 4.87 × 10−4 |
| Reactome | Signaling by receptor tyrosine kinases | 19 | 14.84 | 1.93 × 10−3 |
| MCC | Closeness | Degree | Bottleneck | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Rank | Name | Score | Rank | Name | Score | Rank | Name | Score | Rank | Name | Score |
| 1 | STAT1 | 367,933 | 1 | STAT1 | 35.93333 | 1 | STAT1 | 17 | 1 | RAC2 | 52 |
| 2 | CXCL10 | 367,932 | 2 | SPI1 | 35.91667 | 2 | CXCL10 | 14 | 2 | SPI1 | 43 |
| 3 | SAMD9L | 367,920 | 3 | RAC2 | 34.78333 | 3 | SPI1 | 13 | 3 | STAT1 | 33 |
| 3 | XAF1 | 367,920 | 4 | S100A9 | 32.45 | 3 | RAC2 | 13 | 4 | PRKCE | 14 |
| 3 | IFI44L | 367,920 | 5 | CXCL10 | 31.13571 | 5 | S100A9 | 11 | 5 | BAIAP2 | 13 |
| 3 | OAS2 | 367,920 | 6 | LILRB2 | 30.66667 | 6 | OAS3 | 10 | 6 | DLG4 | 11 |
| 3 | ISG15 | 367,920 | 7 | S100A8 | 30.2 | 6 | SAMD9L | 10 | 7 | ADCY5 | 10 |
| 8 | OAS1 | 362,886 | 8 | PRKCE | 29.93333 | 6 | XAF1 | 10 | 7 | S100A9 | 10 |
| 9 | OAS3 | 362,881 | 9 | TREM1 | 29.08333 | 6 | OAS1 | 10 | 9 | CACNA1C | 8 |
| 10 | HERC5 | 362,880 | 10 | GBP5 | 28.71905 | 6 | IFI44L | 10 | 10 | FXR1 | 6 |
| Category | Term/Function | Genes Enriched | Percentage | p-Value |
|---|---|---|---|---|
| Molecular function | 2′-5′-oligoadenylate synthetase activity | 3 | 30 | 7.83 × 10−8 |
| Adenylyltransferase activity | 3 | 30 | 8.74 × 10−5 | |
| Double-stranded RNA binding | 3 | 30 | 1.46 × 10−3 | |
| Nucleotidyltransferase activity | 3 | 30 | 7.40 × 10−3 | |
| Biological Processes | Defense response to virus | 8 | 80 | 2.42 × 10−11 |
| Defense response to symbiont | 8 | 80 | 2.42 × 10−11 | |
| Response to virus | 8 | 80 | 2.91 × 10−10 | |
| Type I interferon signaling pathway | 6 | 60 | 2.00 × 10−9 | |
| Cellular response to type I interferon | 6 | 60 | 2.14 × 10−9 | |
| Pathways | Coronavirus disease—COVID-19 | 6 | 60 | 2.52 × 10−8 |
| NOD-like receptor signaling pathway | 4 | 40 | 1.55 × 10−4 | |
| Antiviral mechanism by IFN-stimulated genes | 6 | 60 | 8.34 × 10−10 | |
| Interferon signaling | 7 | 70 | 1.08 × 10−9 | |
| Cytokine signaling in immune system | 8 | 80 | 7.65 × 10−8 |
| Based on Hub Genes Module | |||||
|---|---|---|---|---|---|
| Top 10 Drugs | |||||
| S. No. | Score | Mechanism of Action | Status | Year of Approval | |
| 1 | RUXOLITINIB | −0.89 | JAK inhibitor | Approved | 2011 |
| 2 | LENALIDOMIDE | −0.86 | Carcinogen | Approved | 2005 |
| 3 | PAZOPANIB | −0.83 | VEGFR inhibitor|KIT inhibitor|PDGFR inhibitor | Approved | 2009 |
| 4 | PF-04457845 | −0.83 | FAAH inhibitor | Phase: 2 | N/A |
| 5 | PRT-062070 | −0.82 | JAK inhibitor|Syk inhibitor | Phase: 2 | N/A |
| 6 | TRAPIDIL | −0.81 | PDGFR inhibitor | Phase: 1 | N/A |
| 7 | EUGENOL | −0.81 | Androgen receptor antagonist | Phase: 1 | N/A |
| 8 | SR-59230A | −0.79 | Adrenergic receptor antagonist | Phase: 1 | N/A |
| 9 | UNC-669 | −0.78 | L3MBTL antagonist | Phase: 1 | N/A |
| Based on Total Differentially Expressed Genes | |||||
| Top 10 Drugs | |||||
| 1 | Linezolid | −0.54 | Bacterial 50S ribosomal subunit inhibitor | Approved | 2000 |
| 2 | Erastin | −0.54 | Ion channel antagonist | N/A | |
| 3 | Enalapril | −0.53 | ACE inhibitor | Approved | 1985 |
| 4 | AZD-8055 | −0.53 | MTOR inhibitor | Phase: 1 | N/A |
| 5 | Carbamazepine | −0.52 | Carboxamide antiepileptic | Approved | 1968 |
| 6 | BX-795 | −0.52 | IKK inhibitor | N/A | |
| 7 | Methotrexate | −0.52 | Dihydrofolate reductase inhibitor | Approved | 1953 |
| 8 | Eugenol | −0.51 | Androgen receptor antagonist | Phase: 1 | N/A |
| 9 | Tolterodine | −0.51 | Acetylcholine receptor antagonist | Approved | 1998 |
| 10 | Selumetinib | −0.51 | MEK inhibitor | Approved | 2020 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alzahrani, F.A.; Khan, M.F.; Ahmad, V. Recognition of Differentially Expressed Molecular Signatures and Pathways Associated with COVID-19 Poor Prognosis in Glioblastoma Patients. Int. J. Mol. Sci. 2023, 24, 3562. https://doi.org/10.3390/ijms24043562
Alzahrani FA, Khan MF, Ahmad V. Recognition of Differentially Expressed Molecular Signatures and Pathways Associated with COVID-19 Poor Prognosis in Glioblastoma Patients. International Journal of Molecular Sciences. 2023; 24(4):3562. https://doi.org/10.3390/ijms24043562
Chicago/Turabian StyleAlzahrani, Faisal A., Mohd Faheem Khan, and Varish Ahmad. 2023. "Recognition of Differentially Expressed Molecular Signatures and Pathways Associated with COVID-19 Poor Prognosis in Glioblastoma Patients" International Journal of Molecular Sciences 24, no. 4: 3562. https://doi.org/10.3390/ijms24043562
APA StyleAlzahrani, F. A., Khan, M. F., & Ahmad, V. (2023). Recognition of Differentially Expressed Molecular Signatures and Pathways Associated with COVID-19 Poor Prognosis in Glioblastoma Patients. International Journal of Molecular Sciences, 24(4), 3562. https://doi.org/10.3390/ijms24043562