
Update on the Role and Regulatory Mechanism of Extracellular Matrix in the Pathogenesis of Uterine Fibroids



Abstract
:1. Introduction
2. Extracellular Matrix and Regulation in Uterine Fibroids
2.1. ECM and Hormones
2.2. ECM and Growth Factors
2.3. ECM and Cytokines
2.4. Cell Types Contributing to ECM Production in Uterine Fibroids
3. ECM and Downstream Signaling
3.1. Stiff ECM and Progesterone Receptor Signaling
3.2. The Effect of Collagen Cross-Linking on Proliferation and Resistance to MMP Proteolytic Degradation
3.3. ECM Stiffness and Other Signaling
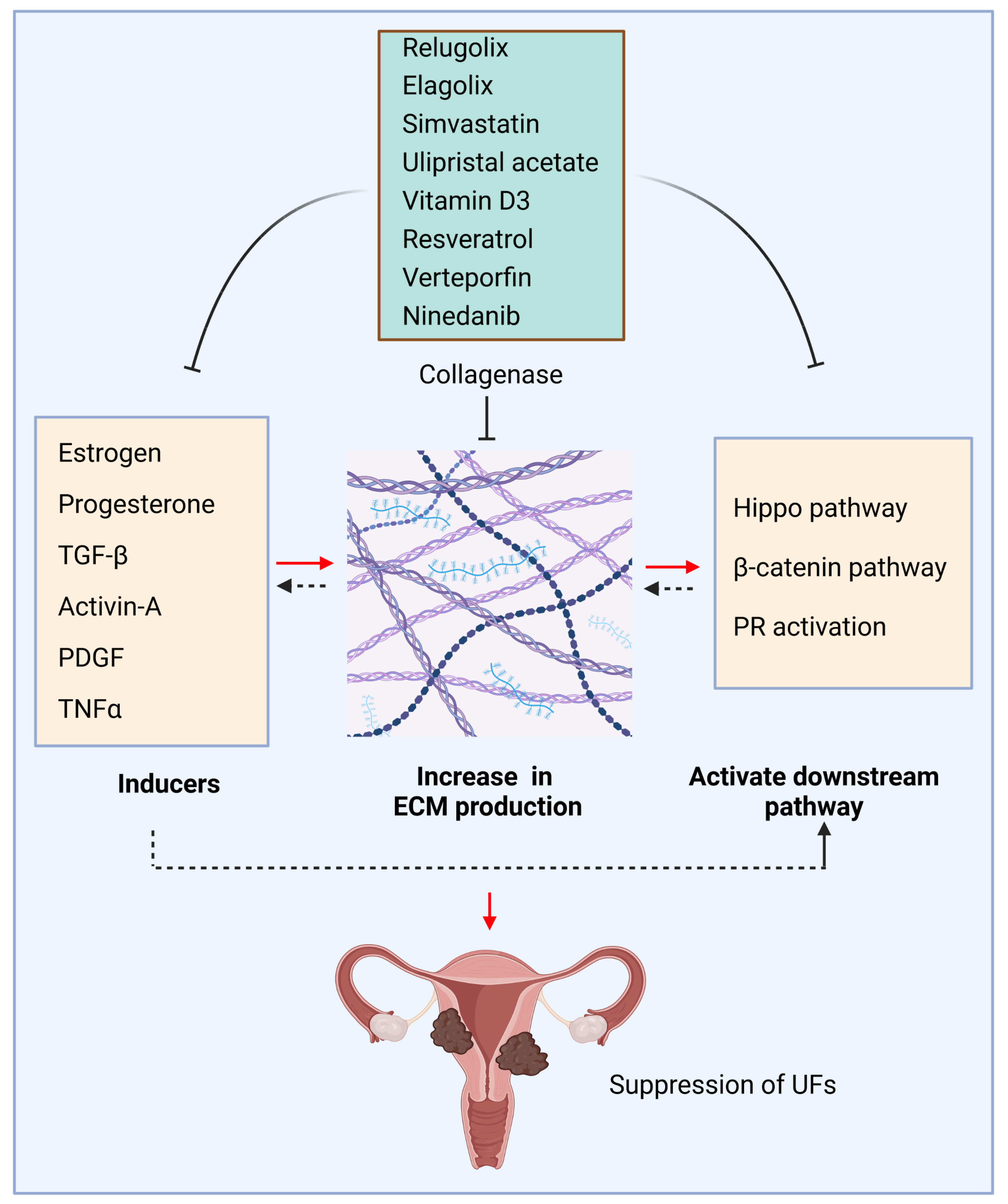
4. Targeting ECM
4.1. Targeting Hormone-Dependent Growth with ECM Changes
4.2. Collagenase
4.3. Vitamin D and ECM
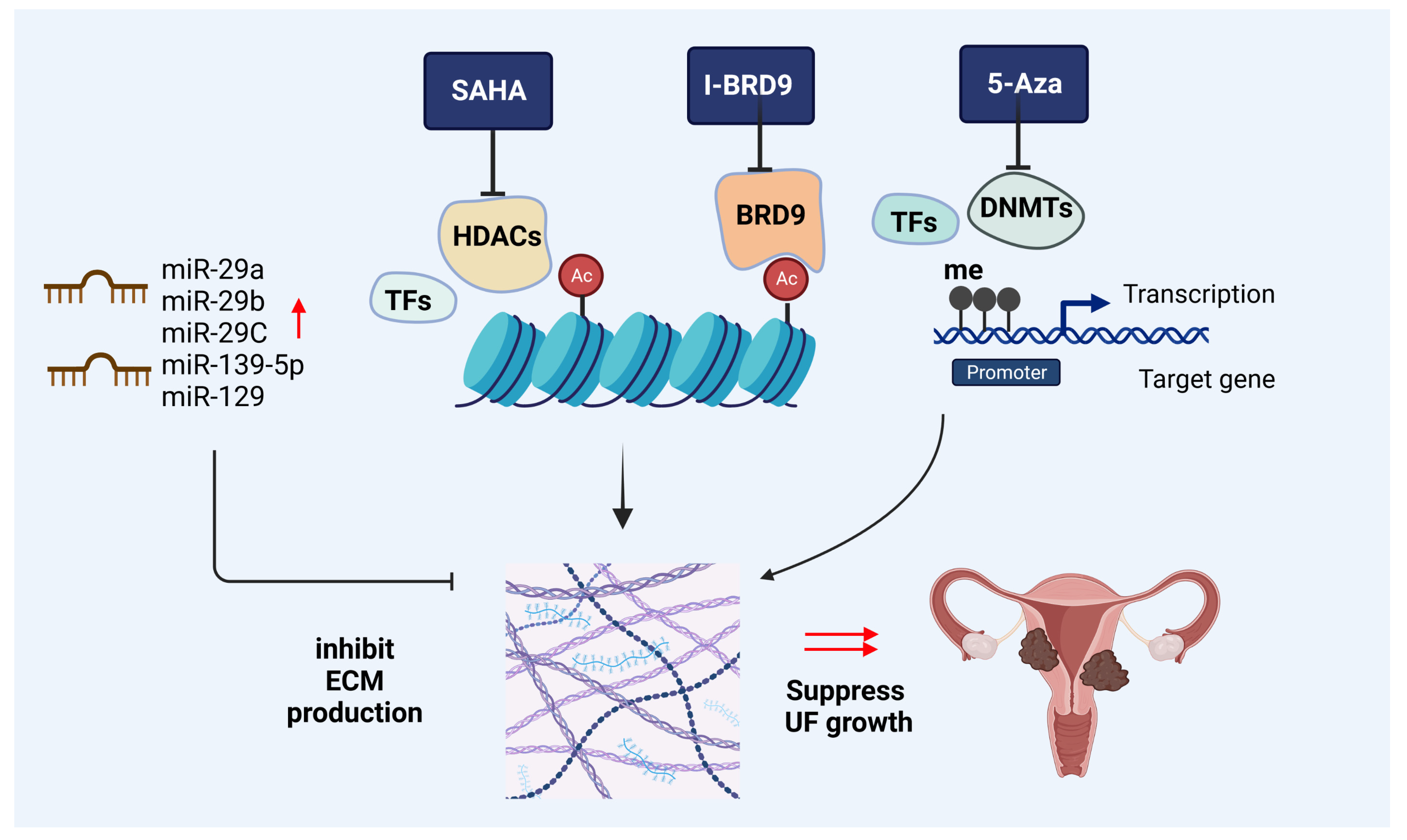
4.4. Epigenetic-Mediated ECM Changes
4.5. ECM and Other Inhibitors

{kind=link}
{kind=link}
| Inducers/Inhibitors | Biological Samples | Changes in ECM | Approach | Co-Changes | Publication Time | References |
|---|---|---|---|---|---|---|
| Activin A | UF cell line | Increase in ECM accumulation | WB | p38 MAPK | July 2021 | [33] |
| Activin A | UF cell line | Excessive ECM | WB, qPCR, and IC | p38 MAPK | October 2018 | [32] |
| Butylated hydroxytoluene | ELT-3 UF cell line | Increase in CO1A1 | WB, IF | PI3K/AKT and MAPK | August 2021 | [105] |
| Leptin | UF cell line | Increase in ECM formation | JAK2/STAT3 and MAPK/ERK | May 2022 | [106] | |
| TBBPA | 3D human UF spheroids | Increase in collagen and fibrosis | Masson’s trichrome stain, PCR, and light microscopy | TGF-β signaling | February 2022 | [107] |
| Cadmium prolonged exposure | UF cells | Decrease in collagens, FNs, laminins; increase in MMPs | IF and MMP antibody array | TGF-β signaling, cell proliferation, and migration | August 2021 | [108] |
| Collagenase Clostridium histolyticum | UF tissues | Decrease in ECM stiffness | Rheometry | Cell proliferation and Hippo signaling | July 2021 | [61] |
| CRSR | UF rat model | Decrease in ECM deposition | Microarray | MAPK, PPAR, Notch, and TGF-β | May 2019 | [109] |
| Decrease in FKBP51 | UF cells | Decrease in ECM formation | qPCR | Cell survival and proliferation | June 2022 | [28] |
| Fucoidan | ELT-3, human UF cells, xenograft model | Decrease in FN and COL1A1 | WB | TGF-β3 signaling | September 2018 | [110] |
| Isoliquiritigenin | ELT3, UtSMC, uterine myometrium hyperplasia mouse model | Decrease in ECM accumulation | WB and IHC | MMP, ERK1/2, p38, and JNK | August 2019 | [111] |
| miR-139-5p | UF cell line | Decrease in contractility of the ECM | Migration, collagen gel contraction, and wound healing | p38 MAPK | August 2021 | [95] |
| miR-21a-5p | UF and MM cell lines | FN, Collagen 1A1, CTGF, versican, and DPT | qPCR and collagen assay | TGF-β3 and MMPs | May 2018 | [112] |
| Relugolix and elagolix | UF cells | Decrease in collagen 1A1, FN, and versican | pERK/ERK | August 2022 | [66] | |
| Resveratrol | Xenograft and UF primary cells | Decrease in collagen I, FN, and a-SMA | WB, qPCR | Proliferation and apoptosis | April 2019 | [77] |
| Simvastatin | UF SCs | Decrease in collagen I and fibronectin | WB | TGF-β and β-catenin signaling | February 2022 | [97] |
| Simvastatin | UF cell lines and primary cells, 2D and 3D | Decrease in collagen I, collagen III, FN, versican, and brevican | WB and IHC | Apoptosis | December 2018 | [99] |
| S1P | UF and myometrium cells | Decrease in FN and collagen 1A1 | qPCR | Decrease in activin A | June 2021 | [113] |
| Butylated hydroxytoluene | UF primary cells | Decrease in collagen I and fibronectin | WB | Cell cycle and TGF-β3 signaling | February 2022 | [75] |
| Ultrasound-guided collagenase injection | UF tissues | Decrease in content, density, and fibers of ECM | Masson’s trichrome stain, second harmonic generation, and Picrosirius stain | Decrease in UF-related pain | September 2021 | [114] |
| Ulipristal acetate | UF patients | Decrease in versican and FN1 | Masson trichrome staining and IHC | Decrease in UF size | February 2018 | [104] |
| Vitamin D | Xenograft | Decrease in collagen I and plasminogen activator inhibitor 1 | WB | TGF-β3 signaling | January 2020 | [115] |
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, Q.; Ciebiera, M.; Bariani, M.V.; Ali, M.; Elkafas, H.; Boyer, T.G.; Al-Hendy, A. Comprehensive Review of Uterine Fibroids: Developmental Origin, Pathogenesis, and Treatment. Endocr. Rev. 2022, 43, 678–719. [Google Scholar] [CrossRef]
- Segars, J.H.; Al-Hendy, A. Uterine Leiomyoma: New Perspectives on an Old Disease. Semin. Reprod. Med. 2017, 35, 471–472. [Google Scholar] [CrossRef]
- Bulun, S.E.; Moravek, M.B.; Yin, P.; Ono, M.; Coon, J.S.t.; Dyson, M.T.; Navarro, A.; Marsh, E.E.; Zhao, H.; Maruyama, T.; et al. Uterine Leiomyoma Stem Cells: Linking Progesterone to Growth. Semin. Reprod. Med. 2015, 33, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, C.M.; Steiner, C.A. Hysterectomy rates in the United States 1990–1997. Obstet. Gynecol. 2002, 99, 229–234. [Google Scholar] [CrossRef]
- Cardozo, E.R.; Clark, A.D.; Banks, N.K.; Henne, M.B.; Stegmann, B.J.; Segars, J.H. The estimated annual cost of uterine leiomyomata in the United States. Am. J. Obstet. Gynecol. 2012, 206, 211.e1–e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mas, A.; Stone, L.; O’Connor, P.M.; Yang, Q.; Kleven, D.; Simon, C.; Walker, C.L.; Al-Hendy, A. Developmental Exposure to Endocrine Disruptors Expands Murine Myometrial Stem Cell Compartment as a Prerequisite to Leiomyoma Tumorigenesis. Stem Cells 2017, 35, 666–678. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Ali, M.; El Andaloussi, A.; Al-Hendy, A. The emerging spectrum of early life exposure-related inflammation and epigenetic therapy. Cancer Stud. Mol. Med. 2018, 4, 13–23. [Google Scholar] [CrossRef]
- Orciani, M.; Caffarini, M.; Biagini, A.; Lucarini, G.; Delli Carpini, G.; Berretta, A.; Di Primio, R.; Ciavattini, A. Chronic Inflammation May Enhance Leiomyoma Development by the Involvement of Progenitor Cells. Stem Cells Int. 2018, 2018, 1716246. [Google Scholar] [CrossRef]
- Chuang, T.D.; Khorram, O. miR-200c regulates IL8 expression by targeting IKBKB: A potential mediator of inflammation in leiomyoma pathogenesis. PLoS ONE 2014, 9, e95370. [Google Scholar] [CrossRef] [PubMed]
- Muralimanoharan, S.; Shamby, R.; Stansbury, N.; Schenken, R.; de la Pena Avalos, B.; Javanmardi, S.; Dray, E.; Sung, P.; Boyer, T.G. Aberrant R-loop-induced replication stress in MED12-mutant uterine fibroids. Sci. Rep. 2022, 12, 6169. [Google Scholar] [CrossRef] [PubMed]
- Prusinski Fernung, L.E.; Yang, Q.; Sakamuro, D.; Kumari, A.; Mas, A.; Al-Hendy, A. Endocrine disruptor exposure during development increases incidence of uterine fibroids by altering DNA repair in myometrial stem cells. Biol. Reprod. 2018, 99, 735–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Nair, S.; Laknaur, A.; Ismail, N.; Diamond, M.P.; Al-Hendy, A. The Polycomb Group Protein EZH2 Impairs DNA Damage Repair Gene Expression in Human Uterine Fibroids. Biol. Reprod. 2016, 94, 69. [Google Scholar] [CrossRef] [PubMed]
- Stewart, E.A.; Laughlin-Tommaso, S.K.; Catherino, W.H.; Lalitkumar, S.; Gupta, D.; Vollenhoven, B. Uterine fibroids. Nat. Rev. Dis. Primers. 2016, 2, 16043. [Google Scholar] [CrossRef] [PubMed]
- Bulun, S.E. Uterine fibroids. N. Engl. J. Med. 2013, 369, 1344–1355. [Google Scholar] [CrossRef] [Green Version]
- Jayes, F.L.; Liu, B.; Feng, L.; Aviles-Espinoza, N.; Leikin, S.; Leppert, P.C. Evidence of biomechanical and collagen heterogeneity in uterine fibroids. PLoS ONE 2019, 14, e0215646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yatsenko, S.A.; Mittal, P.; Wood-Trageser, M.A.; Jones, M.W.; Surti, U.; Edwards, R.P.; Sood, A.K.; Rajkovic, A. Highly heterogeneous genomic landscape of uterine leiomyomas by whole exome sequencing and genome-wide arrays. Fertil. Steril. 2017, 107, 457–466.e459. [Google Scholar] [CrossRef] [Green Version]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and Biological Attributes of Matrix Metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar] [CrossRef] [Green Version]
- Rogers, R.; Norian, J.; Malik, M.; Christman, G.; Abu-Asab, M.; Chen, F.; Korecki, C.; Iatridis, J.; Catherino, W.H.; Tuan, R.S.; et al. Mechanical homeostasis is altered in uterine leiomyoma. Am. J. Obstet. Gynecol. 2008, 198, 474.e1–474.e11. [Google Scholar] [CrossRef] [Green Version]
- Rafique, S.; Segars, J.H.; Leppert, P.C. Mechanical Signaling and Extracellular Matrix in Uterine Fibroids. Semin. Reprod. Med. 2017, 35, 487–493. [Google Scholar] [CrossRef]
- Leppert, P.C.; Jayes, F.L.; Segars, J.H. The extracellular matrix contributes to mechanotransduction in uterine fibroids. Obstet. Gynecol. Int. 2014, 2014, 783289. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, H.; Ishi, K.; Serna, V.A.; Kakazu, R.; Bulun, S.E.; Kurita, T. Progesterone is essential for maintenance and growth of uterine leiomyoma. Endocrinology 2010, 151, 2433–2442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borahay, M.A.; Asoglu, M.R.; Mas, A.; Adam, S.; Kilic, G.S.; Al-Hendy, A. Estrogen Receptors and Signaling in Fibroids: Role in Pathobiology and Therapeutic Implications. Reprod. Sci. 2017, 24, 1235–1244. [Google Scholar] [CrossRef]
- Zbucka, M.; Miltyk, W.; Bielawski, T.; Surazynski, A.; Palka, J.; Wolczynski, S. Mechanism of collagen biosynthesis up-regulation in cultured leiomyoma cells. Folia. Histochem. Cytobiol. 2007, 45 (Suppl. S1), S181–S185. [Google Scholar] [PubMed]
- Islam, M.S.; Ciavattini, A.; Petraglia, F.; Castellucci, M.; Ciarmela, P. Extracellular matrix in uterine leiomyoma pathogenesis: A potential target for future therapeutics. Hum. Reprod. Update 2018, 24, 59–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, M.; Britten, J.; Cox, J.; Patel, A.; Catherino, W.H. Gonadotropin-releasing hormone analogues inhibit leiomyoma extracellular matrix despite presence of gonadal hormones. Fertil. Steril. 2016, 105, 214–224. [Google Scholar] [CrossRef] [Green Version]
- Rein, T. FK506 binding protein 51 integrates pathways of adaptation: FKBP51 shapes the reactivity to environmental change. Bioessays 2016, 38, 894–902. [Google Scholar] [CrossRef]
- Li, L.; Lou, Z.; Wang, L. The role of FKBP5 in cancer aetiology and chemoresistance. Br. J. Cancer 2011, 104, 19–23. [Google Scholar] [CrossRef] [Green Version]
- New, E.P.; Semerci, N.; Ozmen, A.; Guo, X.; Jonnalagadda, V.A.; Kim, J.W.; Anderson, M.L.; Guzeloglu-Kayisli, O.; Imudia, A.N.; Lockwood, C.J.; et al. FKBP51 Contributes to Uterine Leiomyoma Pathogenesis by Inducing Cell Proliferation and Extracellular Matrix Deposition. Reprod. Sci. 2022, 29, 1939–1949. [Google Scholar] [CrossRef]
- DiMauro, A.; Seger, C.; Minor, B.; Amitrano, A.M.; Okeke, I.; Taya, M.; Rackow, A.R.; Kumar, D.; Kottman, R.M.; Bhagavath, B.; et al. Prolactin is Expressed in Uterine Leiomyomas and Promotes Signaling and Fibrosis in Myometrial Cells. Reprod. Sci. 2022, 29, 2525–2535. [Google Scholar] [CrossRef]
- Yang, Q.; Al-Hendy, A. The Regulatory Functions and the Mechanisms of Long Non-Coding RNAs in Cervical Cancer. Cells 2022, 11, 1149. [Google Scholar] [CrossRef]
- Joseph, D.S.; Malik, M.; Nurudeen, S.; Catherino, W.H. Myometrial cells undergo fibrotic transformation under the influence of transforming growth factor beta-3. Fertil. Steril. 2010, 93, 1500–1508. [Google Scholar] [CrossRef]
- Bao, H.; Sin, T.K.; Zhang, G. Activin A induces leiomyoma cell proliferation, extracellular matrix (ECM) accumulation and myofibroblastic transformation of myometrial cells via p38 MAPK. Biochem. Biophys. Res. Commun. 2018, 504, 447–453. [Google Scholar] [CrossRef]
- Bao, H.; Sin, T.K.; Zhang, G. Activin A induces tumorigenesis of leiomyoma via regulation of p38beta MAPK-mediated signal cascade. Biochem. Biophys. Res. Commun. 2020, 529, 379–385. [Google Scholar] [CrossRef]
- Islam, M.S.; Catherino, W.H.; Protic, O.; Janjusevic, M.; Gray, P.C.; Giannubilo, S.R.; Ciavattini, A.; Lamanna, P.; Tranquilli, A.L.; Petraglia, F.; et al. Role of activin-A and myostatin and their signaling pathway in human myometrial and leiomyoma cell function. J. Clin. Endocrinol. Metab. 2014, 99, E775–E785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadopoulos, N.; Lennartsson, J. The PDGF/PDGFR pathway as a drug target. Mol. Aspects Med. 2018, 62, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Folestad, E.; Kunath, A.; Wagsater, D. PDGF-C and PDGF-D signaling in vascular diseases and animal models. Mol. Aspects Med. 2018, 62, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Suo, G.; Jiang, Y.; Cowan, B.; Wang, J.Y. Platelet-derived growth factor C is upregulated in human uterine fibroids and regulates uterine smooth muscle cell growth. Biol. Reprod. 2009, 81, 749–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, M.; Wang, H.; Zhang, Y.; Lu, S.; Wang, Z. Expression and functional analysis of platelet-derived growth factor in uterine leiomyomata. Cancer Biol. Ther. 2006, 5, 28–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, I.L. Cytokine-mediated inflammation, tumorigenesis, and disease-associated JAK/STAT/SOCS signaling circuits in the CNS. Brain Res. Brain Res. Rev. 2005, 48, 166–177. [Google Scholar] [CrossRef]
- Giblin, M.J.; Ontko, C.D.; Penn, J.S. Effect of cytokine-induced alterations in extracellular matrix composition on diabetic retinopathy-relevant endothelial cell behaviors. Sci. Rep. 2022, 12, 12955. [Google Scholar] [CrossRef]
- Uitto, J.; Kouba, D. Cytokine modulation of extracellular matrix gene expression: Relevance to fibrotic skin diseases. J. Dermatol. Sci. 2000, 24 (Suppl. S1), S60–S69. [Google Scholar] [CrossRef]
- Ciavattini, A.; Di Giuseppe, J.; Stortoni, P.; Montik, N.; Giannubilo, S.R.; Litta, P.; Islam, M.S.; Tranquilli, A.L.; Reis, F.M.; Ciarmela, P. Uterine fibroids: Pathogenesis and interactions with endometrium and endomyometrial junction. Obstet. Gynecol. Int. 2013, 2013, 173184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciebiera, M.; Wlodarczyk, M.; Wrzosek, M.; Wojtyla, C.; Blazej, M.; Nowicka, G.; Lukaszuk, K.; Jakiel, G. TNF-alpha serum levels are elevated in women with clinically symptomatic uterine fibroids. Int. J. Immunopathol. Pharmacol. 2018, 32, 2058738418779461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.S.; Protic, O.; Giannubilo, S.R.; Toti, P.; Tranquilli, A.L.; Petraglia, F.; Castellucci, M.; Ciarmela, P. Uterine leiomyoma: Available medical treatments and new possible therapeutic options. J. Clin. Endocrinol. Metab. 2013, 98, 921–934. [Google Scholar] [CrossRef] [Green Version]
- Peddada, S.D.; Laughlin, S.K.; Miner, K.; Guyon, J.P.; Haneke, K.; Vahdat, H.L.; Semelka, R.C.; Kowalik, A.; Armao, D.; Davis, B.; et al. Growth of uterine leiomyomata among premenopausal black and white women. Proc. Natl. Acad. Sci. USA 2008, 105, 19887–19892. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.; Takai, K.; Weaver, V.M.; Werb, Z. Extracellular matrix degradation and remodeling in development and disease. Cold Spring Harb. Perspect Biol. 2011, 3. [Google Scholar] [CrossRef] [PubMed]
- Holdsworth-Carson, S.J.; Zaitseva, M.; Vollenhoven, B.J.; Rogers, P.A. Clonality of smooth muscle and fibroblast cell populations isolated from human fibroid and myometrial tissues. Mol. Hum. Reprod. 2014, 20, 250–259. [Google Scholar] [CrossRef] [Green Version]
- Goad, J.; Rudolph, J.; Zandigohar, M.; Tae, M.; Dai, Y.; Wei, J.J.; Bulun, S.E.; Chakravarti, D.; Rajkovic, A. Single-cell sequencing reveals novel cellular heterogeneity in uterine leiomyomas. Hum. Reprod. 2022, 37, 2334–2349. [Google Scholar] [CrossRef] [PubMed]
- Luo, N.; Guan, Q.; Zheng, L.; Qu, X.; Dai, H.; Cheng, Z. Estrogen-mediated activation of fibroblasts and its effects on the fibroid cell proliferation. Transl. Res. 2014, 163, 232–241. [Google Scholar] [CrossRef]
- Moore, A.B.; Yu, L.; Swartz, C.D.; Zheng, X.; Wang, L.; Castro, L.; Kissling, G.E.; Walmer, D.K.; Robboy, S.J.; Dixon, D. Human uterine leiomyoma-derived fibroblasts stimulate uterine leiomyoma cell proliferation and collagen type I production, and activate RTKs and TGF beta receptor signaling in coculture. Cell Commun. Signal. 2010, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordeiro Mitchell, C.N.; Islam, M.S.; Afrin, S.; Brennan, J.; Psoter, K.J.; Segars, J.H. Mechanical stiffness augments ligand-dependent progesterone receptor B activation via MEK 1/2 and Rho/ROCK-dependent signaling pathways in uterine fibroid cells. Fertil. Steril. 2021, 116, 255–265. [Google Scholar] [CrossRef]
- Kamel, M.; Wagih, M.; Kilic, G.S.; Diaz-Arrastia, C.R.; Baraka, M.A.; Salama, S.A. Overhydroxylation of Lysine of Collagen Increases Uterine Fibroids Proliferation: Roles of Lysyl Hydroxylases, Lysyl Oxidases, and Matrix Metalloproteinases. Biomed. Res. Int. 2017, 2017, 5316845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, W.; Khan, S.K.; Gvozdenovic-Jeremic, J.; Kim, Y.; Dahlman, J.; Kim, H.; Park, O.; Ishitani, T.; Jho, E.H.; Gao, B.; et al. Hippo signaling interactions with Wnt/beta-catenin and Notch signaling repress liver tumorigenesis. J. Clin. Investig. 2017, 127, 137–152. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, M.K.; Shao, C.; Wang, J.; Wei, Q.; Wang, X.; Collier, Z.; Tang, S.; Liu, H.; Zhang, F.; Huang, J.; et al. Wnt/beta-catenin signaling plays an ever-expanding role in stem cell self-renewal, tumorigenesis and cancer chemoresistance. Genes. Dis. 2016, 3, 11–40. [Google Scholar] [CrossRef] [Green Version]
- Deng, F.; Peng, L.; Li, Z.; Tan, G.; Liang, E.; Chen, S.; Zhao, X.; Zhi, F. YAP triggers the Wnt/beta-catenin signalling pathway and promotes enterocyte self-renewal, regeneration and tumorigenesis after DSS-induced injury. Cell Death Dis. 2018, 9, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moumen, M.; Chiche, A.; Decraene, C.; Petit, V.; Gandarillas, A.; Deugnier, M.A.; Glukhova, M.A.; Faraldo, M.M. Myc is required for beta-catenin-mediated mammary stem cell amplification and tumorigenesis. Mol. Cancer 2013, 12, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, Y.A.; Jamaluddin, M.F.B.; Adebayo, M.; Bajwa, P.; Scott, R.J.; Dharmarajan, A.M.; Nahar, P.; Tanwar, P.S. Extracellular matrix (ECM) activates beta-catenin signaling in uterine fibroids. Reproduction 2018, 155, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Shahin, S.M.; Sabri, N.A.; Al-Hendy, A.; Yang, Q. Activation of beta-Catenin Signaling and its Crosstalk With Estrogen and Histone Deacetylases in Human Uterine Fibroids. J. Clin. Endocrinol. Metab. 2020, 105, e1517–e1535. [Google Scholar] [CrossRef]
- Feng, J.; Gou, J.; Jia, J.; Yi, T.; Cui, T.; Li, Z. Verteporfin, a suppressor of YAP-TEAD complex, presents promising antitumor properties on ovarian cancer. Onco. Targets Ther. 2016, 9, 5371–5381. [Google Scholar] [CrossRef] [Green Version]
- Purdy, M.P.; Ducharme, M.; Haak, A.J.; Ravix, J.; Tan, Q.; Sicard, D.; Prakash, Y.S.; Tschumperlin, D.J.; Stewart, E.A. YAP/TAZ are Activated by Mechanical and Hormonal Stimuli in Myometrium and Exhibit Increased Baseline Activation in Uterine Fibroids. Reprod. Sci. 2020, 27, 1074–1085. [Google Scholar] [CrossRef]
- Islam, M.S.; Afrin, S.; Singh, B.; Jayes, F.L.; Brennan, J.T.; Borahay, M.A.; Leppert, P.C.; Segars, J.H. Extracellular matrix and Hippo signaling as therapeutic targets of antifibrotic compounds for uterine fibroids. Clin. Transl. Med. 2021, 11, e475. [Google Scholar] [CrossRef]
- Bozzini, N.; Rodrigues, C.J.; Petti, D.A.; Bevilacqua, R.G.; Goncalves, S.P.; Pinotti, J.A. Effects of treatment with gonadotropin releasing hormone agonist on the uterine leiomyomata structure. Acta. Obstet. Gynecol. Scand. 2003, 82, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Schlaff, W.D.; Ackerman, R.T.; Al-Hendy, A.; Archer, D.F.; Barnhart, K.T.; Bradley, L.D.; Carr, B.R.; Feinberg, E.C.; Hurtado, S.M.; Kim, J.; et al. Elagolix for Heavy Menstrual Bleeding in Women with Uterine Fibroids. N. Engl. J. Med. 2020, 382, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Al-Hendy, A.; Lukes, A.S.; Poindexter, A.N., 3rd; Venturella, R.; Villarroel, C.; Critchley, H.O.D.; Li, Y.; McKain, L.; Arjona Ferreira, J.C.; Langenberg, A.G.M.; et al. Treatment of Uterine Fibroid Symptoms with Relugolix Combination Therapy. N. Engl. J. Med. 2021, 384, 630–642. [Google Scholar] [CrossRef]
- Donnez, J.; Taylor, H.S.; Stewart, E.A.; Bradley, L.; Marsh, E.; Archer, D.; Al-Hendy, A.; Petraglia, F.; Watts, N.; Gotteland, J.P.; et al. Linzagolix with and without hormonal add-back therapy for the treatment of symptomatic uterine fibroids: Two randomised, placebo-controlled, phase 3 trials. Lancet 2022, 400, 896–907. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.; Britten, J.; Malik, M.; Catherino, W.H. Relugolix and elagolix directly inhibit leiomyoma extracellular matrix production in 2-dimesnional and 3-dimensional cell cultures. FS Sci. 2022, 3, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Jayes, F.L.; Liu, B.; Moutos, F.T.; Kuchibhatla, M.; Guilak, F.; Leppert, P.C. Loss of stiffness in collagen-rich uterine fibroids after digestion with purified collagenase Clostridium histolyticum. Am. J. Obstet. Gynecol. 2016, 215, 596.e1–596.e8. [Google Scholar] [CrossRef]
- Ciebiera, M.; Wlodarczyk, M.; Ciebiera, M.; Zareba, K.; Lukaszuk, K.; Jakiel, G. Vitamin D and Uterine Fibroids-Review of the Literature and Novel Concepts. Int. J. Mol. Sci. 2018, 19, 2051. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, R.; Tabrizi, R.; Hessami, K.; Ashari, H.; Nowrouzi-Sohrabi, P.; Hosseini-Bensenjan, M.; Asadi, N. Correlation of low serum vitamin-D with uterine leiomyoma: A systematic review and meta-analysis. Reprod. Biol. Endocrinol. 2020, 18, 85. [Google Scholar] [CrossRef]
- Halder, S.K.; Osteen, K.G.; Al-Hendy, A. 1,25-dihydroxyvitamin d3 reduces extracellular matrix-associated protein expression in human uterine fibroid cells. Biol. Reprod. 2013, 89, 150. [Google Scholar] [CrossRef]
- Brakta, S.; Diamond, J.S.; Al-Hendy, A.; Diamond, M.P.; Halder, S.K. Role of vitamin D in uterine fibroid biology. Fertil. Steril. 2015, 104, 698–706. [Google Scholar] [CrossRef] [Green Version]
- Wawruszak, A.; Gumbarewicz, E.; Okon, E.; Jeleniewicz, W.; Czapinski, J.; Halasa, M.; Okla, K.; Smok-Kalwat, J.; Bocian, A.; Rivero-Muller, A.; et al. Histone deacetylase inhibitors reinforce the phenotypical markers of breast epithelial or mesenchymal cancer cells but inhibit their migratory properties. Cancer. Manag. Res. 2019, 11, 8345–8358. [Google Scholar] [CrossRef] [Green Version]
- Eckschlager, T.; Plch, J.; Stiborova, M.; Hrabeta, J. Histone Deacetylase Inhibitors as Anticancer Drugs. Int. J. Mol. Sci. 2017, 18, 1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Falahati, A.; Khosh, A.; Mohammed, H.; Kang, W.; Corachan, A.; Bariani, M.V.; Boyer, T.G.; Al-Hendy, A. Targeting Class I Histone Deacetylases in Human Uterine Leiomyosarcoma. Cells 2022, 11, 3801. [Google Scholar] [CrossRef]
- Carbajo-Garcia, M.C.; Garcia-Alcazar, Z.; Corachan, A.; Monleon, J.; Trelis, A.; Faus, A.; Pellicer, A.; Ferrero, H. Histone deacetylase inhibition by suberoylanilide hydroxamic acid: A therapeutic approach to treat human uterine leiomyoma. Fertil. Steril. 2022, 117, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Venturelli, S.; Berger, A.; Bocker, A.; Busch, C.; Weiland, T.; Noor, S.; Leischner, C.; Schleicher, S.; Mayer, M.; Weiss, T.S.; et al. Resveratrol as a pan-HDAC inhibitor alters the acetylation status of histone [corrected] proteins in human-derived hepatoblastoma cells. PLoS ONE 2013, 8, e73097. [Google Scholar] [CrossRef]
- Chen, H.Y.; Lin, P.H.; Shih, Y.H.; Wang, K.L.; Hong, Y.H.; Shieh, T.M.; Huang, T.C.; Hsia, S.M. Natural Antioxidant Resveratrol Suppresses Uterine Fibroid Cell Growth and Extracellular Matrix Formation In Vitro and In Vivo. Antioxidants 2019, 8, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisawa, T.; Filippakopoulos, P. Functions of bromodomain-containing proteins and their roles in homeostasis and cancer. Nat. Rev. Mol. Cell Biol. 2017, 18, 246–262. [Google Scholar] [CrossRef]
- Kulikowski, E.; Rakai, B.D.; Wong, N.C.W. Inhibitors of bromodomain and extra-terminal proteins for treating multiple human diseases. Med. Res. Rev. 2021, 41, 223–245. [Google Scholar] [CrossRef]
- Cheung, K.L.; Kim, C.; Zhou, M.M. The Functions of BET Proteins in Gene Transcription of Biology and Diseases. Front. Mol. Biosci. 2021, 8, 728777. [Google Scholar] [CrossRef]
- Lu, L.; Chen, Z.; Lin, X.; Tian, L.; Su, Q.; An, P.; Li, W.; Wu, Y.; Du, J.; Shan, H.; et al. Inhibition of BRD4 suppresses the malignancy of breast cancer cells via regulation of Snail. Cell Death Differ. 2020, 27, 255–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Ali, M.; He, C.; Boyer, T.G.; Al-Hendy, A. Inhibition of BRD proteins suppresses the phenotype of uterine fibroids via regulation of N6-methyladenosine regulators. Fertil. Steril. 2021, 116, e11. [Google Scholar] [CrossRef]
- Yang, Q.; Mas, A.; Diamond, M.P.; Al-Hendy, A. The Mechanism and Function of Epigenetics in Uterine Leiomyoma Development. Reprod. Sci. 2016, 23, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Navarro, A.; Yin, P.; Ono, M.; Monsivais, D.; Moravek, M.B.; Coon, J.S.t.; Dyson, M.T.; Wei, J.J.; Bulun, S.E. 5-Hydroxymethylcytosine promotes proliferation of human uterine leiomyoma: A biological link to a new epigenetic modification in benign tumors. J. Clin. Endocrinol. Metab. 2014, 99, E2437–E2445. [Google Scholar] [CrossRef]
- Navarro, A.; Yin, P.; Monsivais, D.; Lin, S.M.; Du, P.; Wei, J.J.; Bulun, S.E. Genome-wide DNA methylation indicates silencing of tumor suppressor genes in uterine leiomyoma. PLoS ONE 2012, 7, e33284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbajo-Garcia, M.C.; Corachan, A.; Segura-Benitez, M.; Monleon, J.; Escrig, J.; Faus, A.; Pellicer, A.; Cervello, I.; Ferrero, H. 5-aza-2′-deoxycitidine inhibits cell proliferation, extracellular matrix formation and Wnt/beta-catenin pathway in human uterine leiomyomas. Reprod. Biol. Endocrinol. 2021, 19, 106. [Google Scholar] [CrossRef]
- Chuang, T.D.; Panda, H.; Luo, X.; Chegini, N. miR-200c is aberrantly expressed in leiomyomas in an ethnic-dependent manner and targets ZEBs, VEGFA, TIMP2, and FBLN5. Endocr. Relat. Cancer 2012, 19, 541–556. [Google Scholar] [CrossRef] [Green Version]
- Chuang, T.D.; Luo, X.; Panda, H.; Chegini, N. miR-93/106b and their host gene, MCM7, are differentially expressed in leiomyomas and functionally target F3 and IL-8. Mol. Endocrinol. 2012, 26, 1028–1042. [Google Scholar] [CrossRef]
- Chuang, T.D.; Rehan, A.; Khorram, O. Tranilast induces MiR-200c expression through blockade of RelA/p65 activity in leiomyoma smooth muscle cells. Fertil. Steril. 2020, 113, 1308–1318. [Google Scholar] [CrossRef]
- Ciebiera, M.; Wlodarczyk, M.; Zgliczynski, S.; Lozinski, T.; Walczak, K.; Czekierdowski, A. The Role of miRNA and Related Pathways in Pathophysiology of Uterine Fibroids-From Bench to Bedside. Int. J. Mol. Sci. 2020, 21, 3016. [Google Scholar] [CrossRef] [Green Version]
- Marsh, E.E.; Steinberg, M.L.; Parker, J.B.; Wu, J.; Chakravarti, D.; Bulun, S.E. Decreased expression of microRNA-29 family in leiomyoma contributes to increased major fibrillar collagen production. Fertil. Steril. 2016, 106, 766–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiang, W.; Liu, Z.; Serna, V.A.; Druschitz, S.A.; Liu, Y.; Espona-Fiedler, M.; Wei, J.J.; Kurita, T. Down-regulation of miR-29b is essential for pathogenesis of uterine leiomyoma. Endocrinology 2014, 155, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.D.; Khorram, O. Mechanisms underlying aberrant expression of miR-29c in uterine leiomyoma. Fertil. Steril. 2016, 105, 236–245.e231. [Google Scholar] [CrossRef] [Green Version]
- Zota, A.R.; Geller, R.J.; VanNoy, B.N.; Marfori, C.Q.; Tabbara, S.; Hu, L.Y.; Baccarelli, A.A.; Moawad, G.N. Phthalate Exposures and MicroRNA Expression in Uterine Fibroids: The FORGE Study. Epigenet. Insights 2020, 13, 2516865720904057. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.H.; Kim, H.; Lee, I.; Lee, J.H.; Cho, S.; Choi, Y.S. MicroRNA-139-5p Regulates Fibrotic Potentials via Modulation of Collagen Type 1 and Phosphorylated p38 MAPK in Uterine Leiomyoma. Yonsei. Med. J. 2021, 62, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.L.; Zhao, L.; Han, S.C.; Bi, J.L.; Liu, H.X.; Yue, C.; Lin, L. MiR-129 is involved in the occurrence of uterine fibroid through inhibiting TET1. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 4419–4426. [Google Scholar] [CrossRef]
- Afrin, S.; Ali, M.; El Sabeh, M.; Yang, Q.; Al-Hendy, A.; Borahay, M.A. Simvastatin inhibits stem cell proliferation in human leiomyoma via TGF-beta3 and Wnt/beta-Catenin pathways. J. Cell Mol. Med. 2022, 26, 1684–1698. [Google Scholar] [CrossRef]
- Borahay, M.A.; Vincent, K.; Motamedi, M.; Sbrana, E.; Kilic, G.S.; Al-Hendy, A.; Boehning, D. Novel effects of simvastatin on uterine fibroid tumors: In vitro and patient-derived xenograft mouse model study. Am. J. Obstet. Gynecol. 2015, 213, 196.e1–196.e8. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.; Britten, J.; Borahay, M.; Segars, J.; Catherino, W.H. Simvastatin, at clinically relevant concentrations, affects human uterine leiomyoma growth and extracellular matrix production. Fertil. Steril. 2018, 110, 1398–1407.e1. [Google Scholar] [CrossRef]
- Afrin, S.; Islam, M.S.; Patzkowsky, K.; Malik, M.; Catherino, W.H.; Segars, J.H.; Borahay, M.A. Simvastatin ameliorates altered mechanotransduction in uterine leiomyoma cells. Am. J. Obstet. Gynecol. 2020, 223, 733.e1–733.e14. [Google Scholar] [CrossRef]
- Galliano, D. Ulipristal acetate in uterine fibroids. Fertil. Steril. 2015, 103, 359–360. [Google Scholar] [CrossRef]
- Talaulikar, V.S.; Manyonda, I.T. Ulipristal acetate: A novel option for the medical management of symptomatic uterine fibroids. Adv. Ther. 2012, 29, 655–663. [Google Scholar] [CrossRef]
- Talaulikar, V.S.; Manyonda, I. Ulipristal acetate for use in moderate to severe symptoms of uterine fibroids. Womens Health 2014, 10, 565–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, J.; Malik, M.; Britten, J.; Lewis, T.; Catherino, W.H. Ulipristal Acetate and Extracellular Matrix Production in Human Leiomyomas In Vivo: A Laboratory Analysis of a Randomized Placebo Controlled Trial. Reprod. Sci. 2018, 25, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.F.; Chen, H.Y.; Ali, M.; Shieh, T.M.; Huang, Y.J.; Wang, K.L.; Chang, H.Y.; Huang, T.C.; Hong, Y.H.; Hsia, S.M. The Role of Cell Proliferation and Extracellular Matrix Accumulation Induced by Food Additive Butylated Hydroxytoluene in Uterine Leiomyoma. Nutrients 2021, 13, 3074. [Google Scholar] [CrossRef] [PubMed]
- Reschke, L.; Afrin, S.; El Sabah, M.; Charewycz, N.; Miyashita-Ishiwata, M.; Borahay, M.A. Leptin induces leiomyoma cell proliferation and extracellular matrix deposition via JAK2/STAT3 and MAPK/ERK pathways. FS Sci. 2022, 3, 383–391. [Google Scholar] [CrossRef]
- Liu, J.; Yu, L.; Castro, L.; Yan, Y.; Clayton, N.P.; Bushel, P.; Flagler, N.D.; Scappini, E.; Dixon, D. Short-term tetrabromobisphenol A exposure promotes fibrosis of human uterine fibroid cells in a 3D culture system through TGF-beta signaling. FASEB J. 2022, 36, e22101. [Google Scholar] [CrossRef]
- Yan, Y.; Liu, J.; Lawrence, A.; Dykstra, M.J.; Fannin, R.; Gerrish, K.; Tucker, C.J.; Scappini, E.; Dixon, D. Prolonged cadmium exposure alters benign uterine fibroid cell behavior, extracellular matrix components, and TGFB signaling. FASEB J. 2021, 35, e21738. [Google Scholar] [CrossRef]
- Yu, C.H.; Zhao, J.S.; Zhao, H.; Peng, T.; Shen, D.C.; Xu, Q.X.; Li, Y.; Webb, R.C.; Wang, M.H.; Shi, X.M.; et al. Transcriptional profiling of uterine leiomyoma rats treated by a traditional herb pair, Curcumae rhizoma and Sparganii rhizoma. Braz. J. Med. Biol. Res. 2019, 52, e8132. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.Y.; Huang, T.C.; Lin, L.C.; Shieh, T.M.; Wu, C.H.; Wang, K.L.; Hong, Y.H.; Hsia, S.M. Fucoidan Inhibits the Proliferation of Leiomyoma Cells and Decreases Extracellular Matrix-Associated Protein Expression. Cell Physiol. Biochem. 2018, 49, 1970–1986. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.H.; Kung, H.L.; Chen, H.Y.; Huang, K.C.; Hsia, S.M. Isoliquiritigenin Suppresses E2-Induced Uterine Leiomyoma Growth through the Modulation of Cell Death Program and the Repression of ECM Accumulation. Cancers 2019, 11, 1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardozo, E.R.; Foster, R.; Karmon, A.E.; Lee, A.E.; Gatune, L.W.; Rueda, B.R.; Styer, A.K. MicroRNA 21a-5p overexpression impacts mediators of extracellular matrix formation in uterine leiomyoma. Reprod. Biol. Endocrinol. 2018, 16, 46. [Google Scholar] [CrossRef] [PubMed]
- Bernacchioni, C.; Ciarmela, P.; Vannuzzi, V.; Greco, S.; Vannuccini, S.; Malentacchi, F.; Pellegrino, P.; Capezzuoli, T.; Sorbi, F.; Cencetti, F.; et al. Sphingosine 1-phosphate signaling in uterine fibroids: Implication in activin A pro-fibrotic effect. Fertil. Steril. 2021, 115, 1576–1585. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Sims, H.; Trueheart, I.; Simpson, K.; Wang, K.C.; Patzkowsky, K.; Wegman, T.; Soma, J.M.; Dixon, R.; Jayes, F.; et al. A Phase I Clinical Trial to Assess Safety and Tolerability of Injectable Collagenase in Women with Symptomatic Uterine Fibroids. Reprod. Sci. 2021, 28, 2699–2709. [Google Scholar] [CrossRef] [PubMed]
- Corachan, A.; Ferrero, H.; Escrig, J.; Monleon, J.; Faus, A.; Cervello, I.; Pellicer, A. Long-term vitamin D treatment decreases human uterine leiomyoma size in a xenograft animal model. Fertil. Steril. 2020, 113, 205–216.e4. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Q.; Al-Hendy, A. Update on the Role and Regulatory Mechanism of Extracellular Matrix in the Pathogenesis of Uterine Fibroids. Int. J. Mol. Sci. 2023, 24, 5778. https://doi.org/10.3390/ijms24065778
Yang Q, Al-Hendy A. Update on the Role and Regulatory Mechanism of Extracellular Matrix in the Pathogenesis of Uterine Fibroids. International Journal of Molecular Sciences. 2023; 24(6):5778. https://doi.org/10.3390/ijms24065778
Chicago/Turabian StyleYang, Qiwei, and Ayman Al-Hendy. 2023. "Update on the Role and Regulatory Mechanism of Extracellular Matrix in the Pathogenesis of Uterine Fibroids" International Journal of Molecular Sciences 24, no. 6: 5778. https://doi.org/10.3390/ijms24065778
APA StyleYang, Q., & Al-Hendy, A. (2023). Update on the Role and Regulatory Mechanism of Extracellular Matrix in the Pathogenesis of Uterine Fibroids. International Journal of Molecular Sciences, 24(6), 5778. https://doi.org/10.3390/ijms24065778