
Identification of Orbivirus Non-Structural Protein 5 (NS5), Its Role and Interaction with RNA/DNA in Infected Cells





Abstract
:1. Introduction
2. Results
2.1. Bioinformatics Analyses

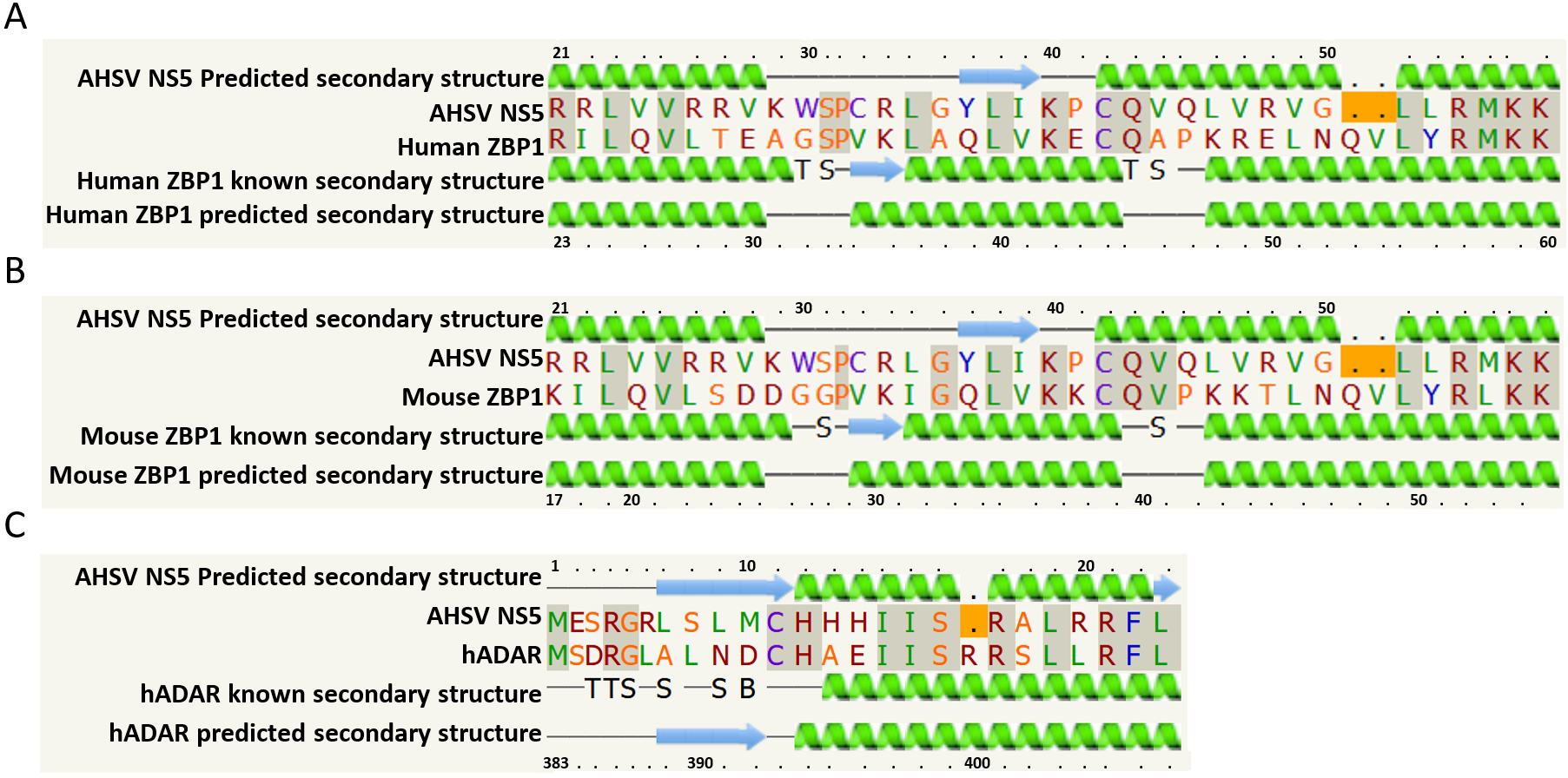{kind=link}
{kind=link}
{kind=link}
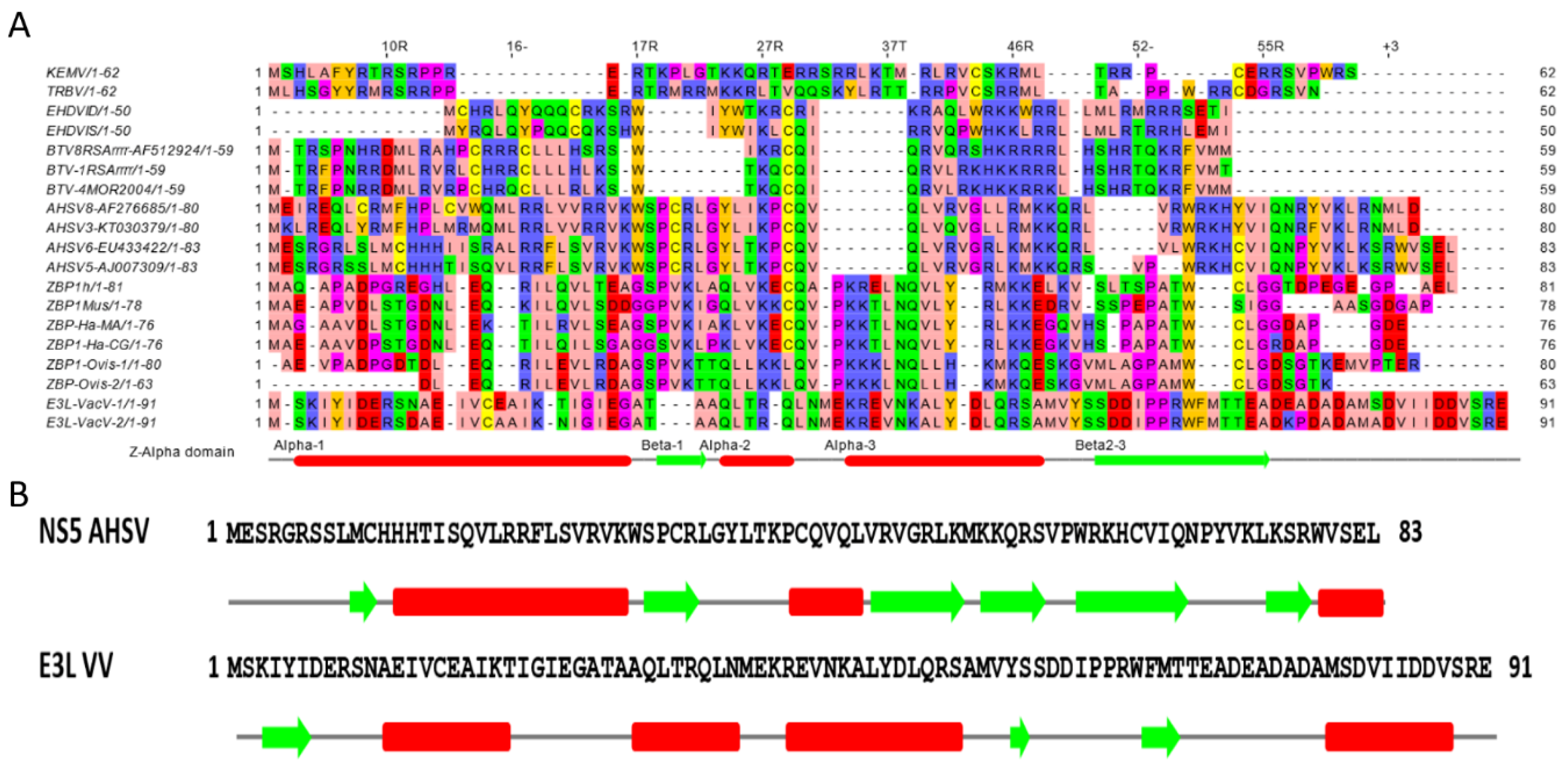{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) | |||
|---|---|---|---|
| Virus | aa Start Position | Monopartite NLS Sequence | Score |
| BTV-1 | 37 | VLRKHKKRRRL | 12.5 |
| 38 | LRKHKKRRRLH | 13.5 | |
| 39 | RKHKKRRRLH | 6 | |
| 39 | RKHKKRRRLHS | 12 | |
| BTV-8 | 37 | VQRSHKRRRRL | 12 |
| 39 | RSHKRRRRLH | 12 | |
| BTV-26 | 28 | VLRKHKKRRRL | 12.5 |
| 29 | LRKHKKRRRLH | 13.5 | |
| 30 | RKHKKRRRLH | 6 | |
| 30 | RKHKKRRRLHS | 12 | |
| AHSV-4 | 50 | GRLKMKKQRLVLW | 6.5 |
| EHDV-7Is | 28 | VQPWHKKLRRL | 6 |
| KEMV | None | ||
| (b) | |||
| Virus | aa Start Position | Bipartite NLS Sequence | Score |
| BTV-1 | 12 | RVRLCHRRCLLLHLKSWTKQCQIQRVLRKHKKR | 7.3 |
| BTV-8 | 26 | RSWIKRCQIQRVQRSHKRRRR | 12.5 |
| 36 | RVQRSHKRRRRLHSHRTQKRFVM | 9 | |
| BTV-26 | 5 | RQRRRRCRLLPSKFWTKRCQTQLVLRKHKKR | 7.7 |
| 5 | RQRRRRCRLLPSKFWTKRCQTQLVLRKHKKR | 6.6 | |
| 17 | KFWTKRCQTQLVLRKHKKRRR | 14 | |
| 17 | KFWTKRCQTQLVLRKHKKRRRL | 13.1 | |
| AHSV-4 | None | ||
| EHDV-7Is | None | ||
| KEMV | None | ||
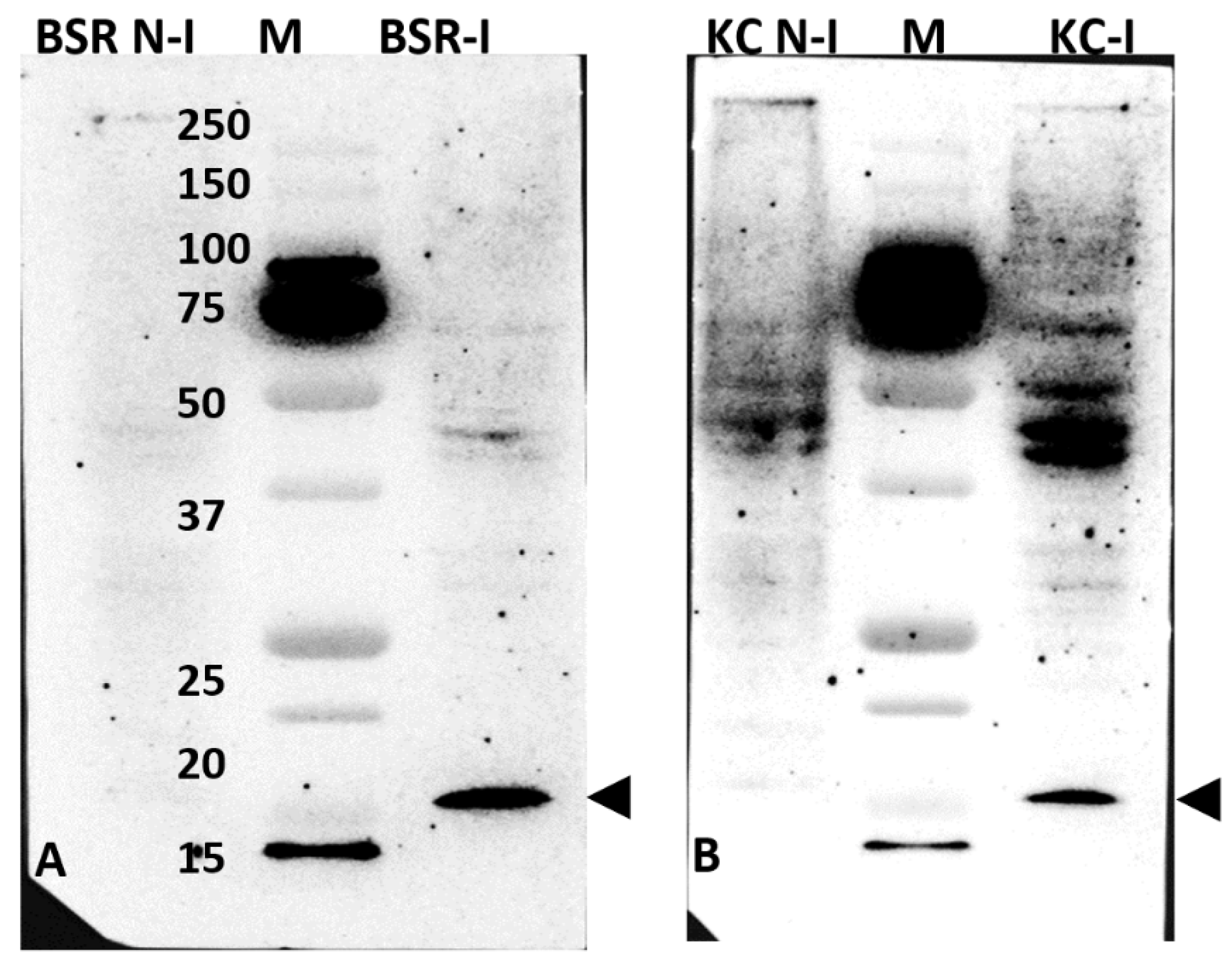
2.2. Western Blot Analysis of BTV-1-Infected BSR or KC Cell Lysates, Using Anti-Sera Raised against GST-NS5 Expressed in Bacteria
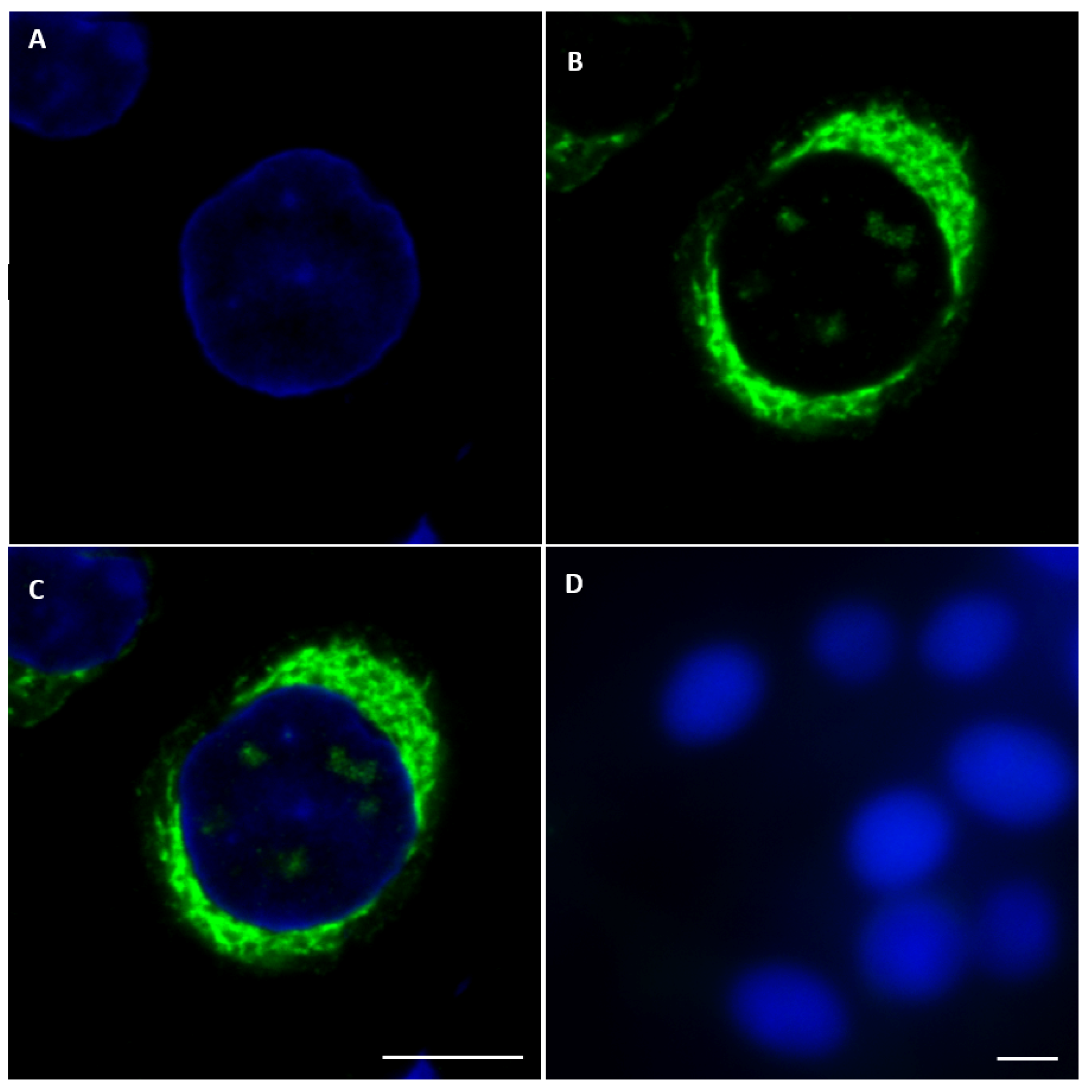
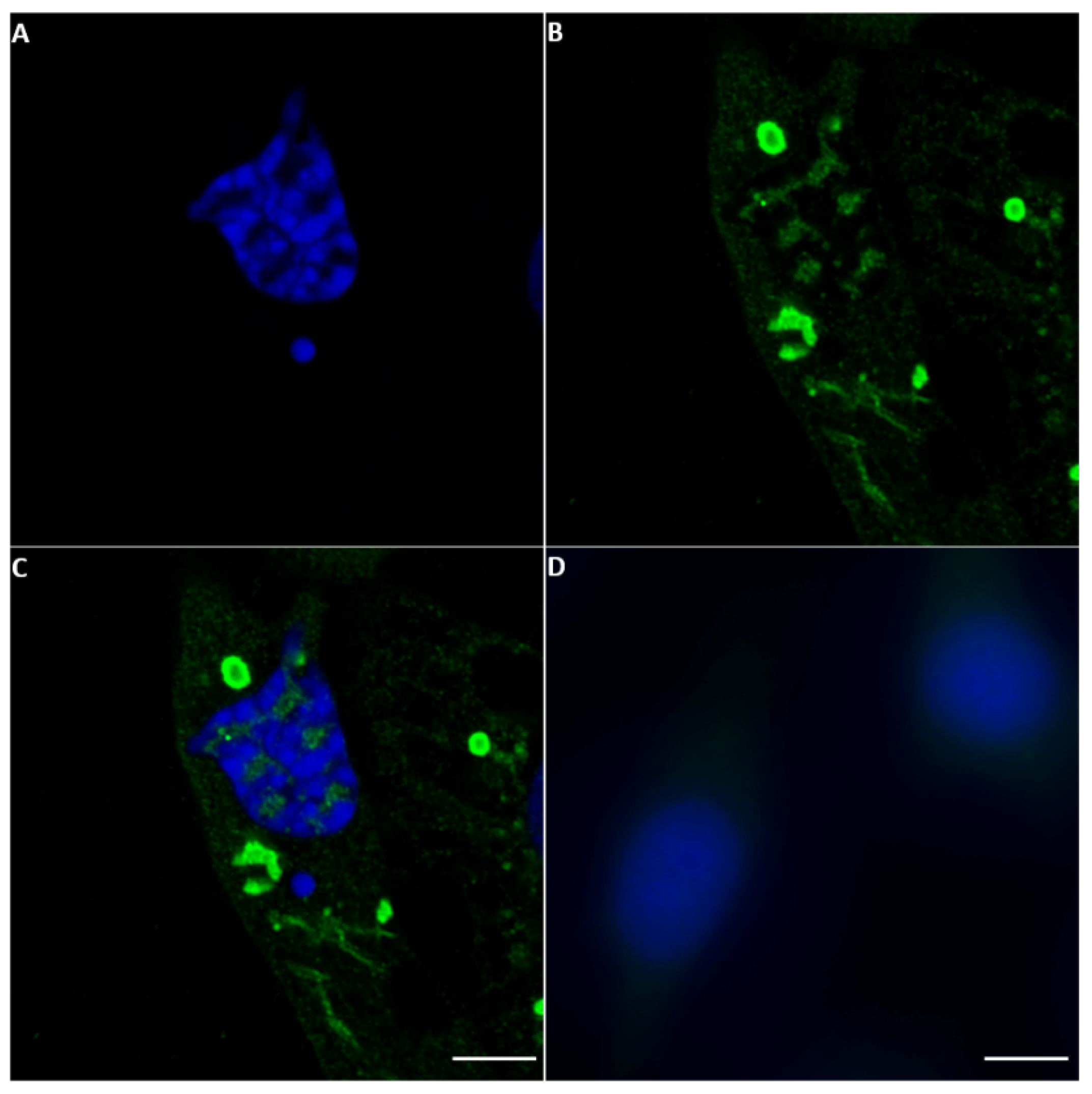
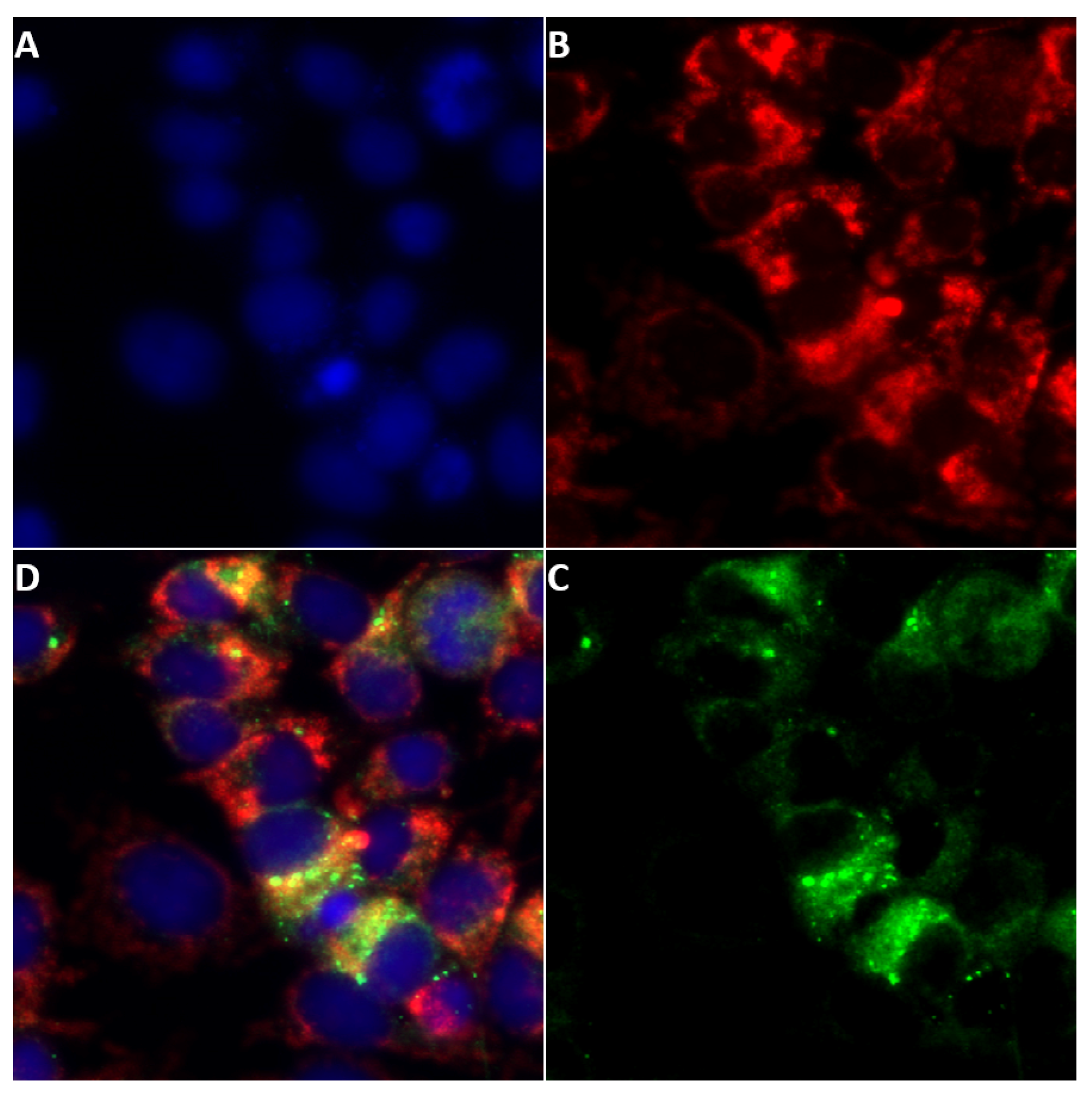
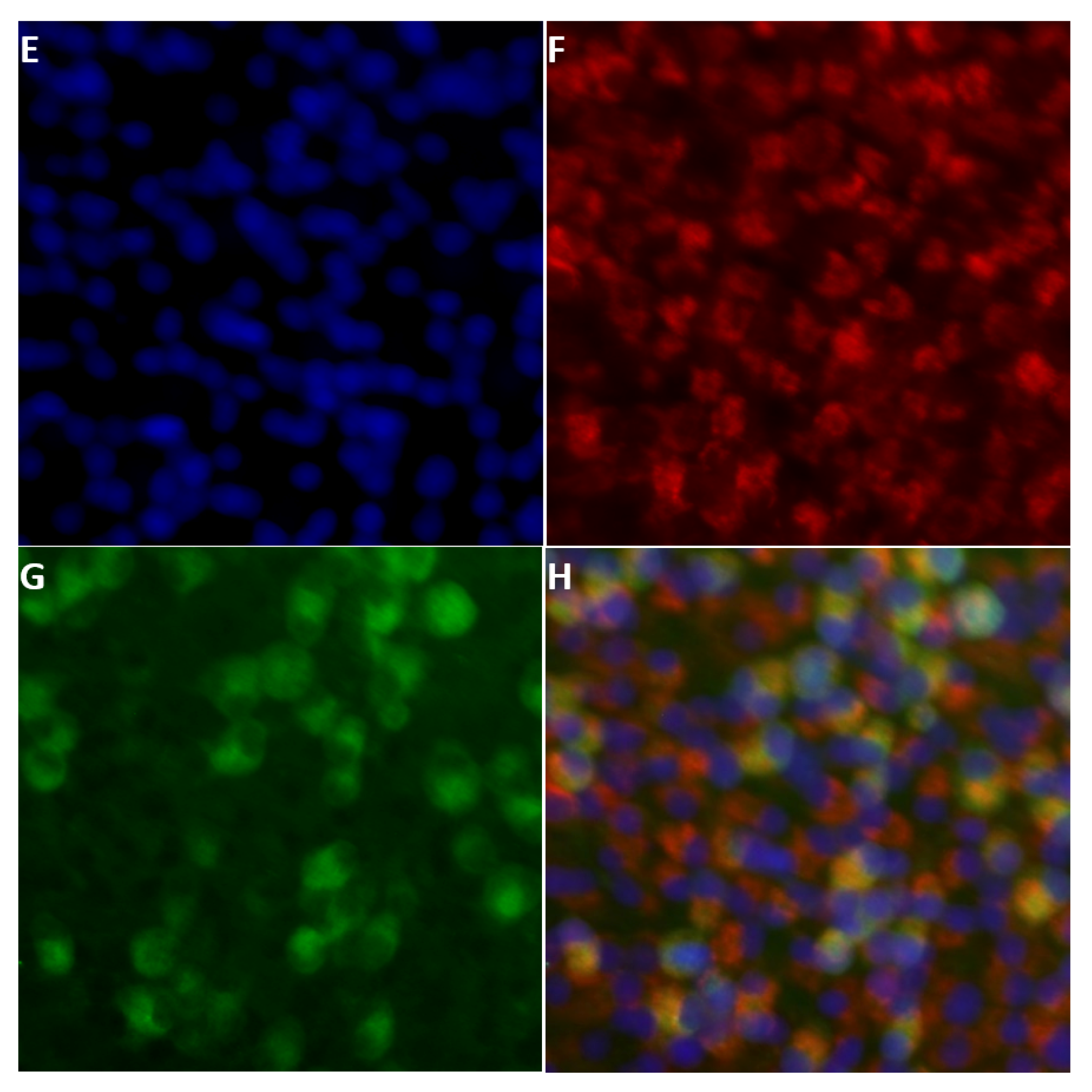
2.3. Detection of NS5 in Infected Cells by Fluorescence Microscopy
2.3.1. BTV NS5
2.3.2. AHSV NS5
2.3.3. KEMV NS5
2.3.4. EHDV NS5
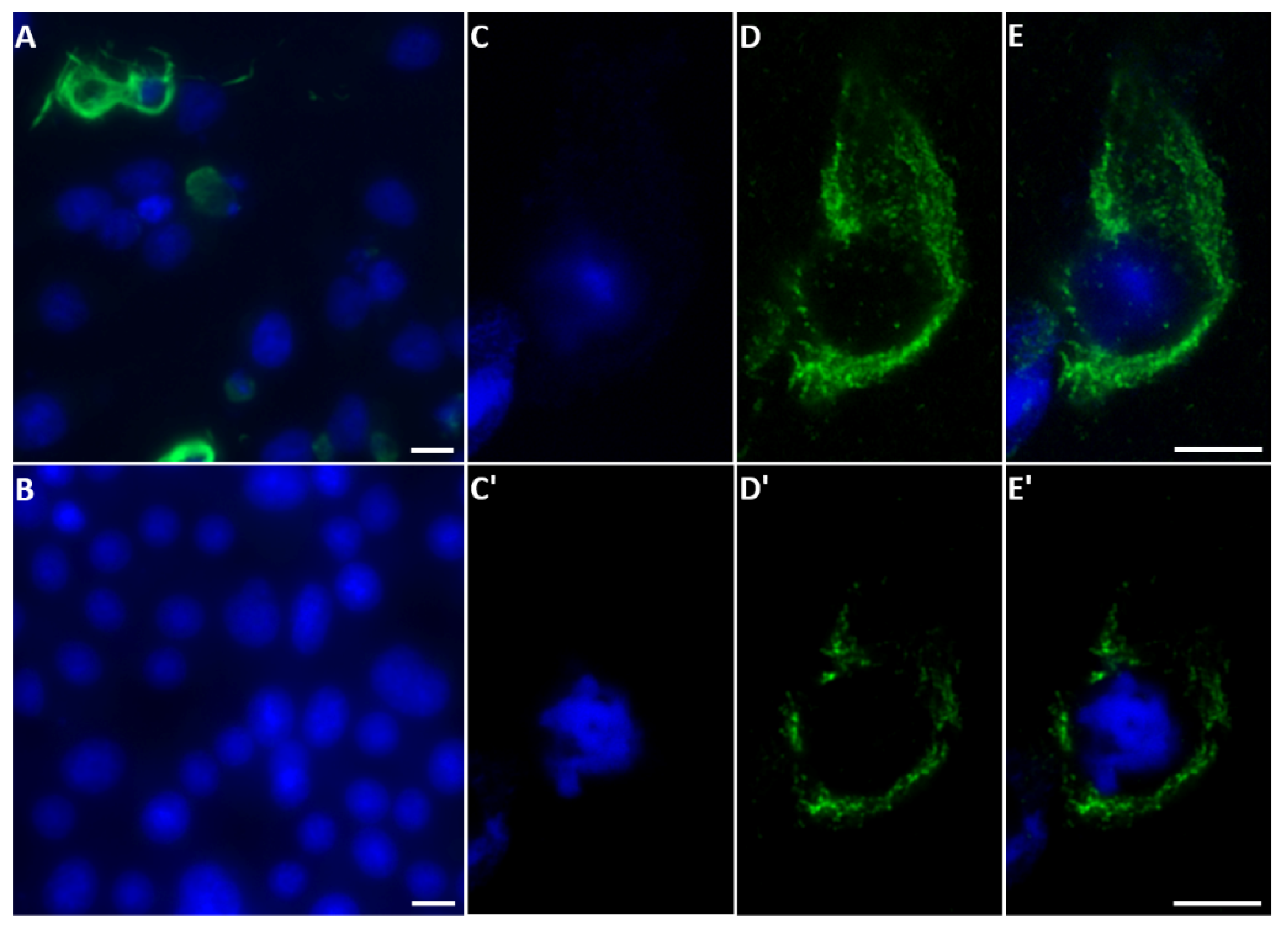
2.4. NS5 Localises to the Nucleolus during Early Stages of Orbivirus Infections
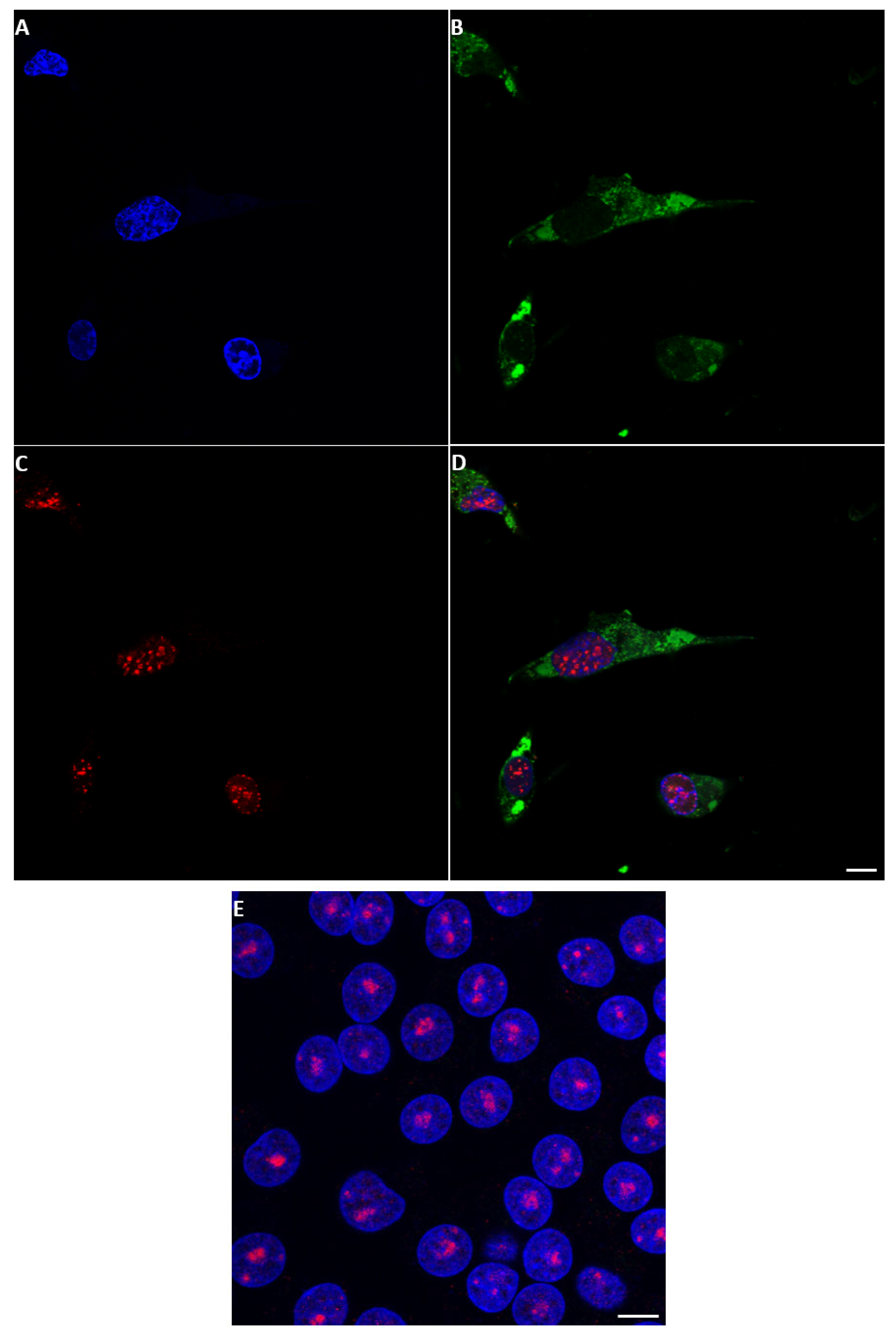
2.5. Co-Expression of NS5 and NS4 or VP5 in BSR Cells
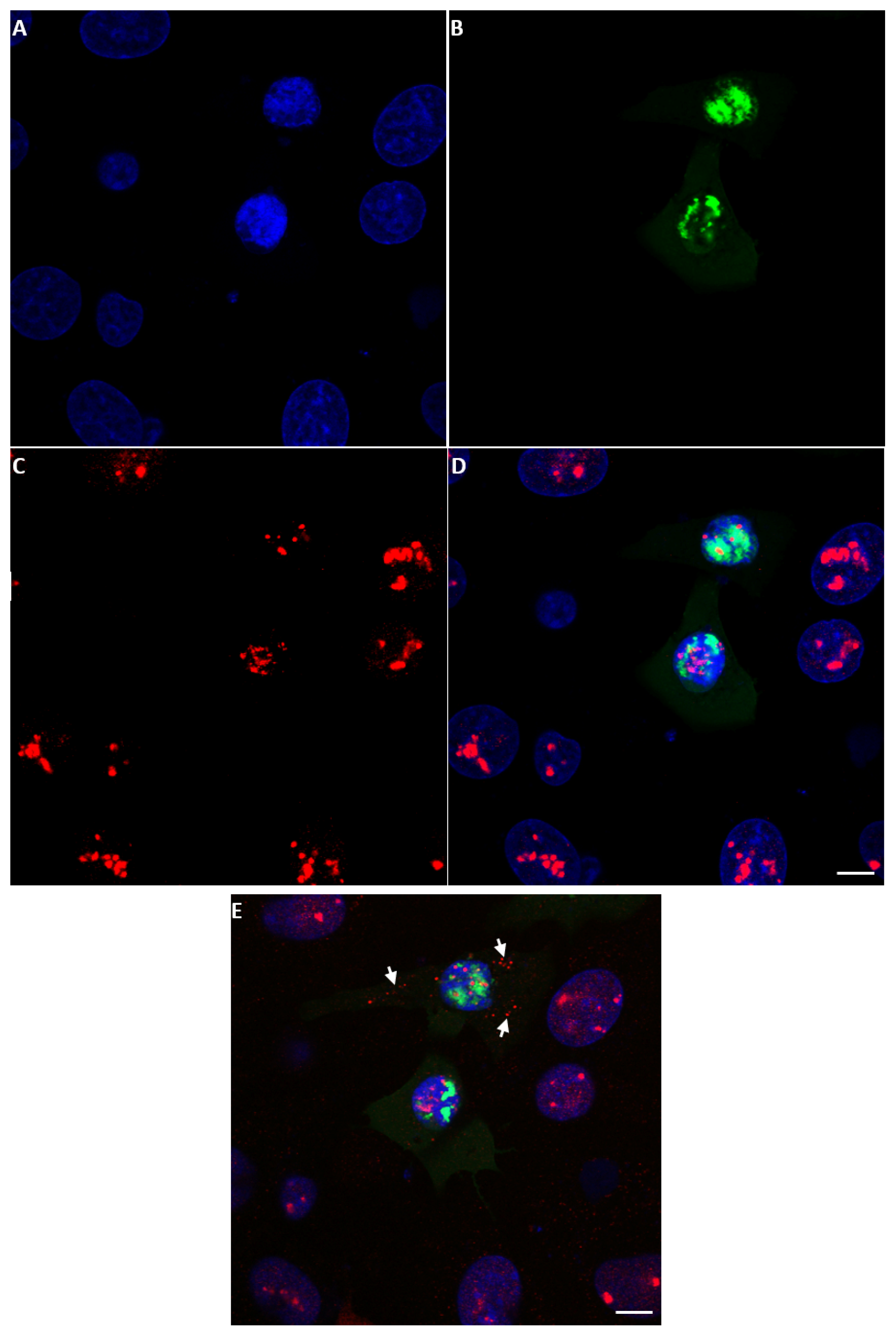
2.6. Localisation of NS5 to Mitochondria
2.7. Pulldown of Protein-Protein Complexes in BSR Cells Expressing His-Tagged NS5
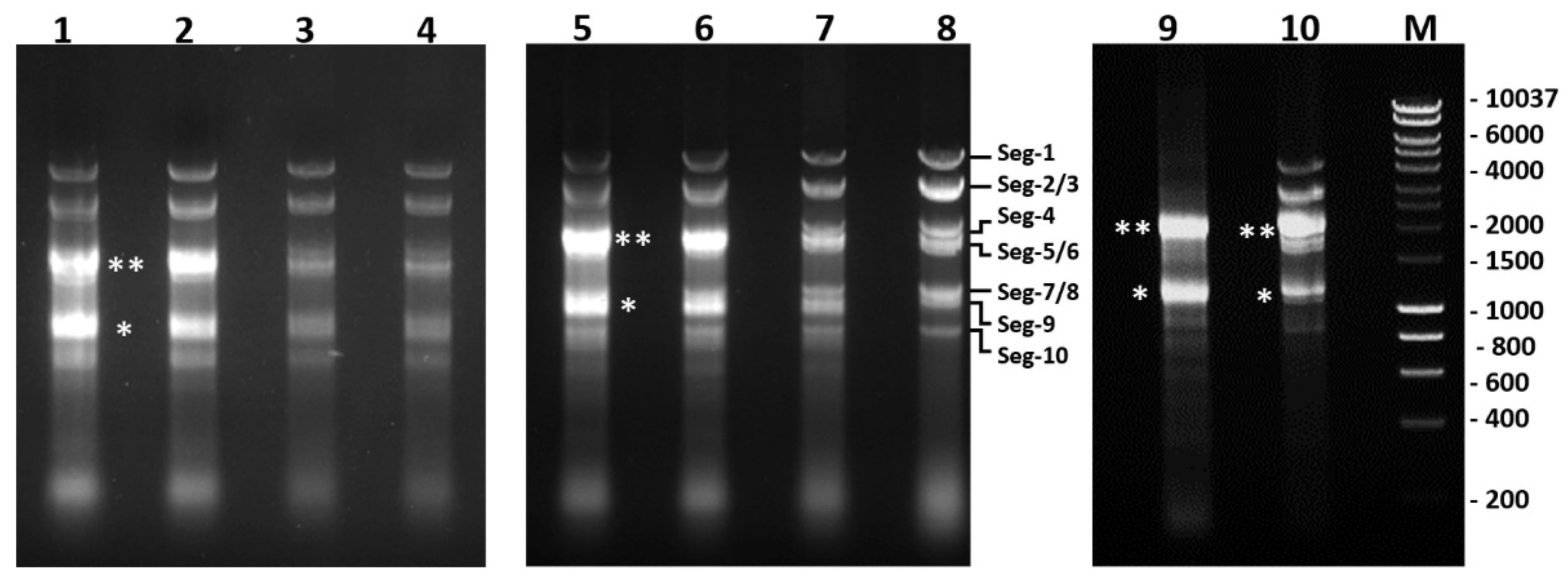
2.8. Replication of BTV-1RGC7, BTV-1ΔNS5, BTV-1ΔNS4 or BTV-1ΔNS5/ΔNS4 in BSR Cells
2.9. Replication of Recombinant Vaccinia Expressing NS5 in A549 Cells
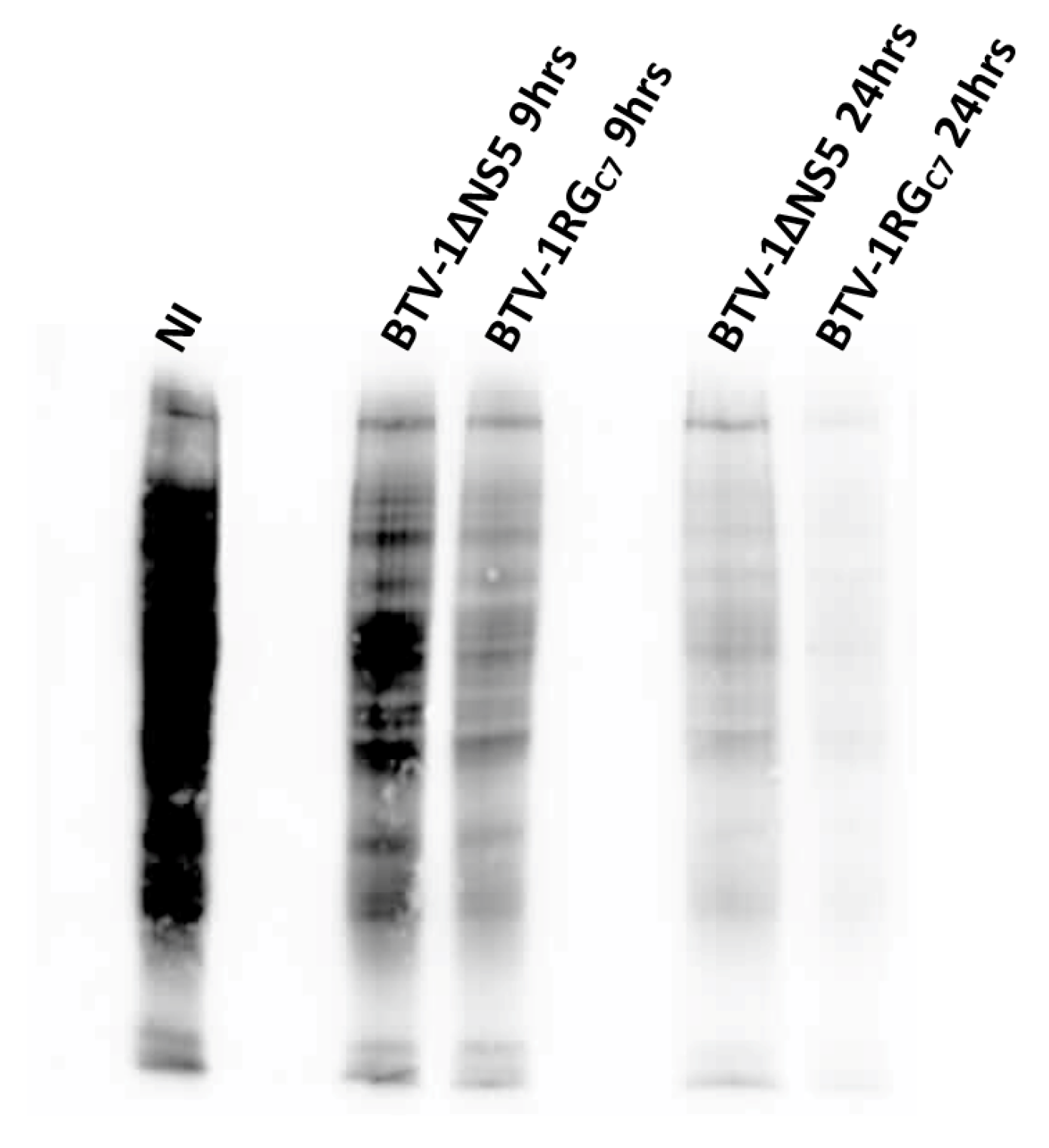
2.10. Assessment of Protein Synthesis in BSR Cells Infected with BTV-1RGC7 or BTV-1ΔNS5
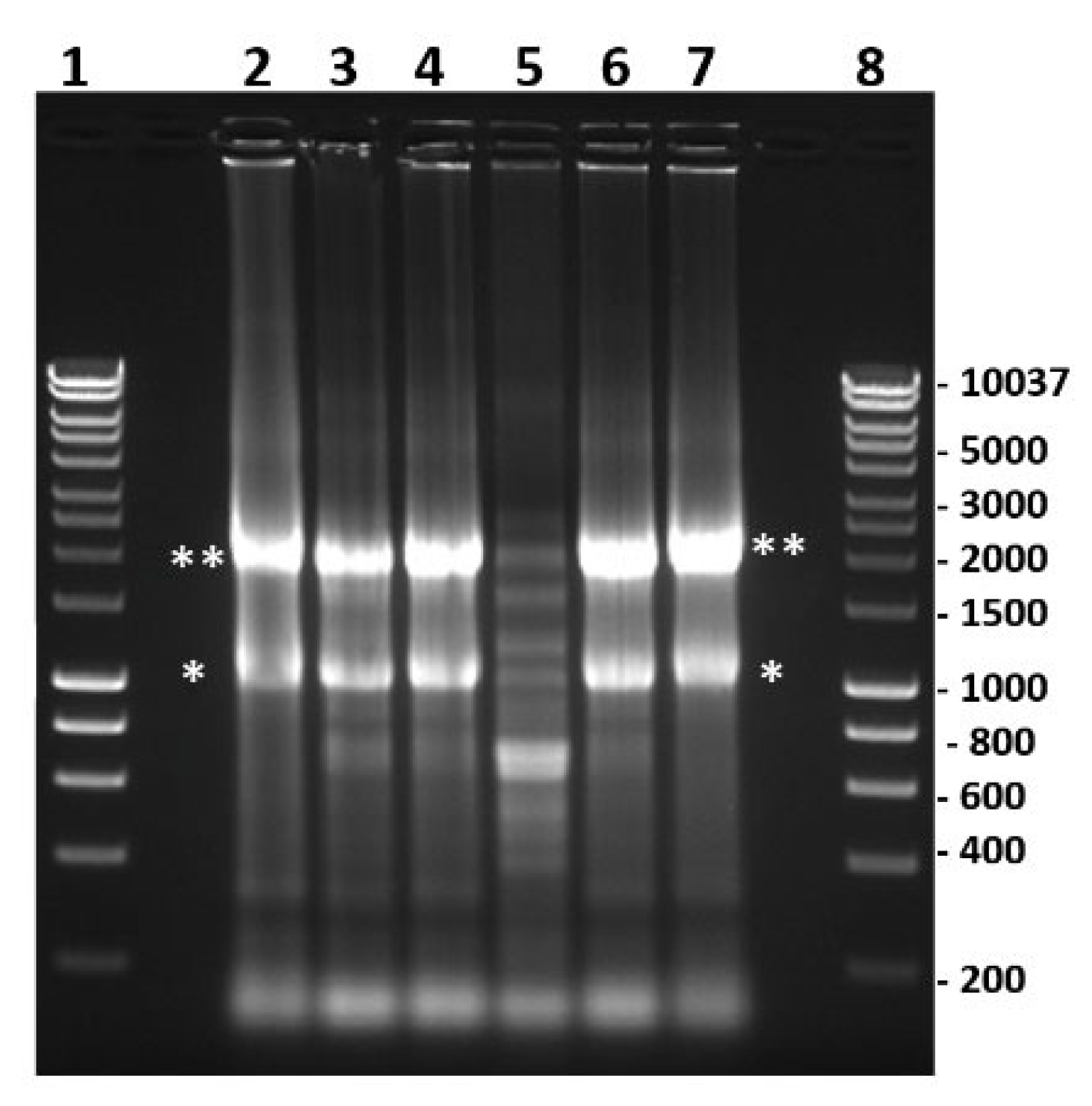
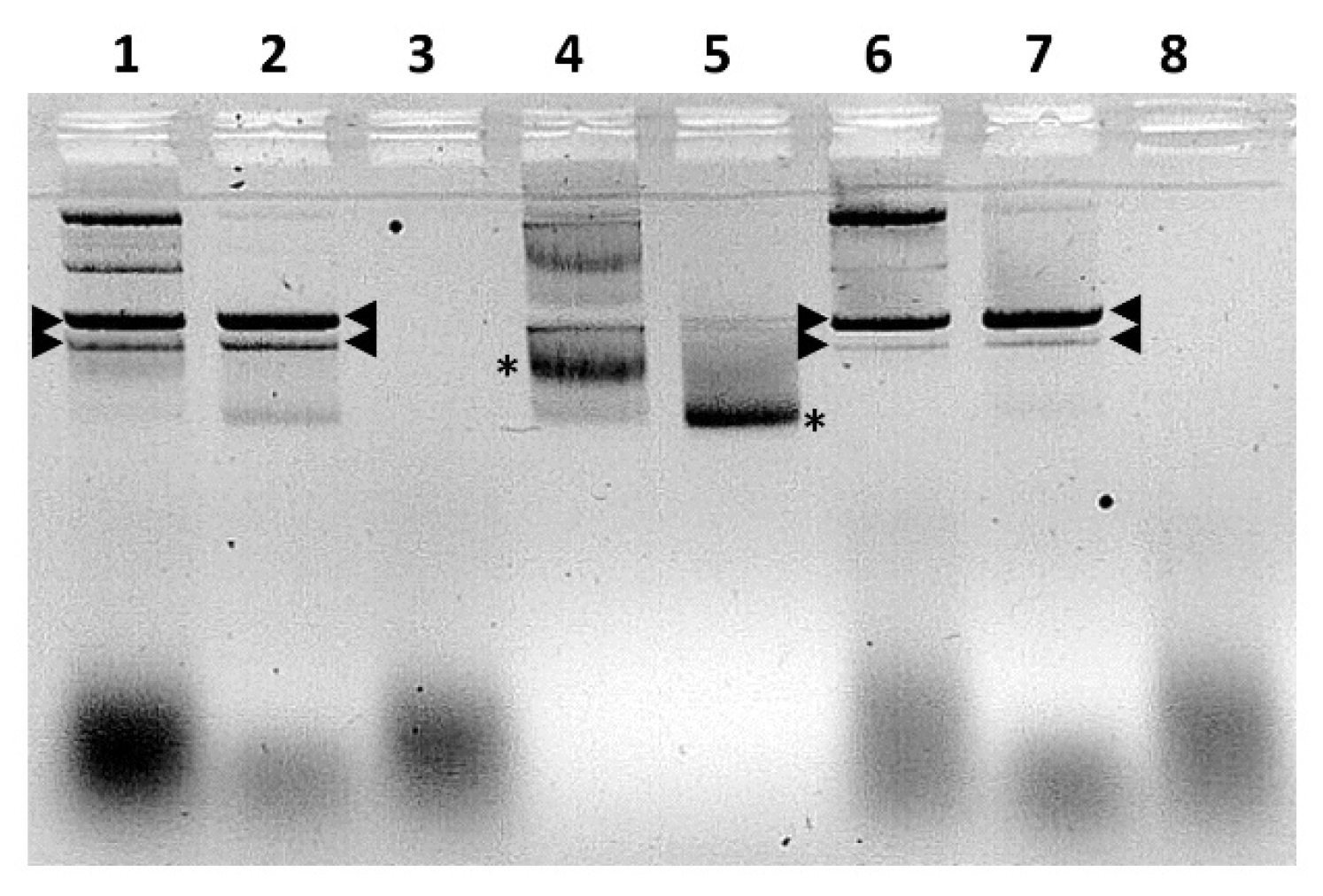
2.11. Nuclease Treatment of Bacterially Expressed and Purified NS5
2.12. Electromobility Shift Assays (EMSA)
2.13. Transfection of BSR Cells with pCI-BTV1NS5-6xHis or pCI-AHSVNS5-6xHis and Infection with BTV-1RGC7 or BTV-1ΔNS5
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Cell Cultures and Viruses
4.3. Bioinformatic Analyses
4.4. Preparation of cDNAS from Orbivirus dsRNA and Cloning of the ORFs of NS4, NS5 and/or VP5 in Expression Plasmids
4.5. Expression of NS5 of BTV, AHSV, EHDV, and KEMV in Bacteria
4.6. Western Blot Analysis of Infected Cell Cultures
4.7. Localisation of NS5 in Orbivirus Infected Cells by Confocal Fluorescence Microscopy
4.8. Localisation of Recombinant Expressed BTV-NS5 by Fluorescence Microscopy in Cells Transfected with pCI-BTV1NS5-GFP, pCI-BTVNS4/pCI-BTV1NS5 or pCI-BTV1NS5-GFP/VP5
4.9. Localisation of NS5 to Mitochondria
4.10. Pulldown of Protein–Protein Complexes Formed in BSR Cells Expressing 6xHis-Tagged NS5
4.11. Generation of BTV-1 Deletion Mutants (ΔNS4, ΔNS5 and ΔNS4/ΔNS5) by Reverse Genetics Using the BTV-1RGC7 Backbone
4.12. Replication of BTV-1RGC7, BTV-1ΔNS5, BTV-1ΔNS4 or BTV-1ΔNS5/ΔNS4 in Mammalian Cells
4.13. Pulldown of NS5-RNA Complexes from BSR Cells Expressing 6xHis-Tagged NS5 of BTV or AHSV, and Infected with BTV-1RGC7 or BTV-1ΔNS5
4.14. Protein Synthesis in BSR Cells Infected with BTV-1RGC7 or BTV-1ΔNS5
4.15. Transfection of BSR Cells with pCI-BTV1NS5-6xHis or pCI-AHSVNS5-6xHis and Infection with BTV-1RGC7 or BTV-1ΔNS5
4.16. Electromobility Shift Assays (EMSA)
4.17. Cloning of NS5 ORF in a Vaccinia Shuttle Vector, Generation of Vaccinia-ΔE3L/+NS5 Recombinant Viruses and Replication in Interferon Treated Cells
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Attoui, H.; Mohd Jaafar, F.; Mertens, P.P.C. The Reovirales a New Taxonomic Order: Families Sedoreoviridae and Spinareoviridae. In Proceedings of the 13th International DsRNA Virus Symposium, Houffalize, Belgium, 24–28 September 2018. [Google Scholar]
- Matthijnssens, J.; Attoui, H.; Banyai, K.; Brussaard, C.P.; Danthi, P.; Del Vas, M.; Dermody, T.S.; Duncan, R.; Fang, Q.; Johne, R.; et al. Ictv Virus Taxonomy Profile: Sedoreoviridae 2022. J. Gen. Virol. 2022, 103, 001782. [Google Scholar] [CrossRef] [PubMed]
- Mohd Jaafar, F.; Belhouchet, M.; Belaganahalli, M.; Tesh, R.B.; Mertens, P.P.; Attoui, H. Full-genome characterisation of Orungo, Lebombo and Changuinola viruses provides evidence for co-evolution of orbiviruses with their arthropod vectors. PLoS ONE 2014, 9, e86392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belaganahalli, M.N.; Maan, S.; Maan, N.S.; Brownlie, J.; Tesh, R.; Attoui, H.; Mertens, P.P. Genetic characterization of the tick-borne orbiviruses. Viruses 2015, 7, 2185–2209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attoui, H.; Mertens, P.P.C.; Becnel, J.; Belaganahalli, M.; Bergoin, M.; Brussaard, C.P.; Chappell, J.D.; Ciarlet, M.; del Vas, M.; Dermody, T.S.; et al. Reoviridae. In Virus Taxonomy. The Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier-Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Mertens, P.P.; Brown, F.; Sangar, D.V. Assignment of the genome segments of bluetongue virus type 1 to the proteins which they encode. Virology 1984, 135, 207–217. [Google Scholar] [CrossRef]
- Grimes, J.M.; Burroughs, J.N.; Gouet, P.; Diprose, J.M.; Malby, R.; Zientara, S.; Mertens, P.P.; Stuart, D.I. The atomic structure of the bluetongue virus core. Nature 1998, 395, 470–478. [Google Scholar] [CrossRef]
- Stuart, D.I.; Gouet, P.; Grimes, J.; Malby, R.; Diprose, J.; Zientara, S.; Burroughs, J.N.; Mertens, P.P. Structural studies of orbivirus particles. Arch. Virol. Suppl. 1998, 14, 235–250. [Google Scholar] [CrossRef]
- Gouet, P.; Diprose, J.M.; Grimes, J.M.; Malby, R.; Burroughs, J.N.; Zientara, S.; Stuart, D.I.; Mertens, P.P. The highly ordered double-stranded RNA genome of bluetongue virus revealed by crystallography. Cell 1999, 97, 481–490. [Google Scholar] [CrossRef] [Green Version]
- Diprose, J.M.; Burroughs, J.N.; Sutton, G.C.; Goldsmith, A.; Gouet, P.; Malby, R.; Overton, I.; Zientara, S.; Mertens, P.P.; Stuart, D.I.; et al. Translocation portals for the substrates and products of a viral transcription complex: The bluetongue virus core. EMBO J. 2001, 20, 7229–7239. [Google Scholar] [CrossRef]
- Stauber, N.; Martinez-Costas, J.; Sutton, G.; Monastyrskaya, K.; Roy, P. Bluetongue virus VP6 protein binds ATP and exhibits an RNA-dependent ATPase function and a helicase activity that catalyze the unwinding of double-stranded RNA substrates. J. Virol. 1997, 71, 7220–7226. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.; Wu, W.; Cui, Y.; Roy, P.; Zhou, Z.H. Bluetongue virus capsid protein VP5 perforates membranes at low endosomal pH during viral entry. Nat. Microbiol. 2021, 6, 1424–1432. [Google Scholar] [CrossRef]
- Maan, S.; Maan, N.S.; Samuel, A.R.; Rao, S.; Attoui, H.; Mertens, P.P.C. Analysis and phylogenetic comparisons of full-length VP2 genes of the 24 bluetongue virus serotypes. J. Gen. Virol. 2007, 88, 621–630. [Google Scholar] [CrossRef]
- Belhouchet, M.; Mohd Jaafar, F.; Firth, A.E.; Grimes, J.M.; Mertens, P.P.; Attoui, H. Detection of a fourth orbivirus non-structural protein. PLoS ONE 2011, 6, e25697. [Google Scholar] [CrossRef] [Green Version]
- Ratinier, M.; Caporale, M.; Golder, M.; Franzoni, G.; Allan, K.; Nunes, S.F.; Armezzani, A.; Bayoumy, A.; Rixon, F.; Shaw, A.; et al. Identification and characterization of a novel non-structural protein of bluetongue virus. PLoS Pathog. 2011, 7, e1002477. [Google Scholar] [CrossRef] [Green Version]
- Kerviel, A.; Ge, P.; Lai, M.; Jih, J.; Boyce, M.; Zhang, X.; Zhou, Z.H.; Roy, P. Atomic structure of the translation regulatory protein NS1 of bluetongue virus. Nat. Microbiol. 2019, 4, 837–845. [Google Scholar] [CrossRef] [Green Version]
- Owens, R.J.; Limn, C.; Roy, P. Role of an arbovirus nonstructural protein in cellular pathogenesis and virus release. J. Virol. 2004, 78, 6649–6656. [Google Scholar] [CrossRef] [Green Version]
- Huismans, H.; Els, H.J. Characterization of the tubules associated with the replication of three different orbiviruses. Virology 1979, 92, 397–406. [Google Scholar] [CrossRef]
- Boyce, M.; Celma, C.C.; Roy, P. Bluetongue virus non-structural protein 1 is a positive regulator of viral protein synthesis. Virol. J. 2012, 9, 178. [Google Scholar] [CrossRef] [Green Version]
- Butan, C.; Tucker, P. Insights into the role of the non-structural protein 2 (NS2) in Bluetongue virus morphogenesis. Virus Res. 2010, 151, 109–117. [Google Scholar] [CrossRef]
- Lymperopoulos, K.; Noad, R.; Tosi, S.; Nethisinghe, S.; Brierley, I.; Roy, P. Specific binding of Bluetongue virus NS2 to different viral plus-strand RNAs. Virology 2006, 353, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Hyatt, A.D.; Zhao, Y.; Roy, P. Release of bluetongue virus-like particles from insect cells is mediated by BTV nonstructural protein NS3/NS3A. Virology 1993, 193, 592–603. [Google Scholar] [CrossRef]
- O’Hara, R.S.; Meyer, A.J.; Burroughs, J.N.; Pullen, L.; Martin, L.A.; Mertens, P.P. Development of a mouse model system, coding assignments and identification of the genome segments controlling virulence of African horse sickness virus serotypes 3 and 8. Arch. Virol. Suppl. 1998, 14, 259–279. [Google Scholar] [CrossRef] [PubMed]
- Labadie, T.; Roy, P. A non-enveloped arbovirus released in lysosome-derived extracellular vesicles induces super-infection exclusion. PLoS Pathog. 2020, 16, e1009015. [Google Scholar] [CrossRef] [PubMed]
- Firth, A.E. Bioinformatic analysis suggests that the Orbivirus VP6 cistron encodes an overlapping gene. Virol. J. 2008, 5, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sealfon, R.S.; Lin, M.F.; Jungreis, I.; Wolf, M.Y.; Kellis, M.; Sabeti, P.C. FRESCo: Finding regions of excess synonymous constraint in diverse viruses. Genome Biol. 2015, 16, 38. [Google Scholar] [CrossRef] [Green Version]
- Stewart, M.; Hardy, A.; Barry, G.; Pinto, R.M.; Caporale, M.; Melzi, E.; Hughes, J.; Taggart, A.; Janowicz, A.; Varela, M.; et al. Characterization of a second open reading frame in genome segment 10 of bluetongue virus. J. Gen. Virol. 2015, 96, 3280–3293. [Google Scholar] [CrossRef]
- Martin, R.M.; Ter-Avetisyan, G.; Herce, H.D.; Ludwig, A.K.; Lattig-Tunnemann, G.; Cardoso, M.C. Principles of protein targeting to the nucleolus. Nucleus 2015, 6, 314–325. [Google Scholar] [CrossRef] [Green Version]
- Kosugi, S.; Hasebe, M.; Tomita, M.; Yanagawa, H. Systematic identification of cell cycle-dependent yeast nucleocytoplasmic shuttling proteins by prediction of composite motifs. Proc. Natl. Acad. Sci. USA 2009, 106, 10171–10176. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Kanneganti, T.D. The regulation of the ZBP1-NLRP3 inflammasome and its implications in pyroptosis, apoptosis, and necroptosis (PANoptosis). Immunol. Rev. 2020, 297, 26–38. [Google Scholar] [CrossRef]
- Bartas, M.; Slychko, K.; Brazda, V.; Cerven, J.; Beaudoin, C.A.; Blundell, T.L.; Pecinka, P. Searching for New Z-DNA/Z-RNA Binding Proteins Based on Structural Similarity to Experimentally Validated Zalpha Domain. Int. J. Mol. Sci. 2022, 23, 768. [Google Scholar] [CrossRef]
- Mizuno, N.; Voordouw, G.; Miki, K.; Sarai, A.; Higuchi, Y. Crystal structure of dissimilatory sulfite reductase D (DsrD) protein–possible interaction with B- and Z-DNA by its winged-helix motif. Structure 2003, 11, 1133–1140. [Google Scholar] [CrossRef]
- Ha, S.C.; Lokanath, N.K.; Van Quyen, D.; Wu, C.A.; Lowenhaupt, K.; Rich, A.; Kim, Y.G.; Kim, K.K. A poxvirus protein forms a complex with left-handed Z-DNA: Crystal structure of a Yatapoxvirus Zalpha bound to DNA. Proc. Natl. Acad. Sci. USA 2004, 101, 14367–14372. [Google Scholar] [CrossRef] [Green Version]
- Komoto, S.; Kanai, Y.; Fukuda, S.; Kugita, M.; Kawagishi, T.; Ito, N.; Sugiyama, M.; Matsuura, Y.; Kobayashi, T.; Taniguchi, K. Reverse Genetics System Demonstrates that Rotavirus Nonstructural Protein NSP6 Is Not Essential for Viral Replication in Cell Culture. J. Virol. 2017, 91, e00695–e00717. [Google Scholar] [CrossRef] [Green Version]
- Holloway, G.; Johnson, R.I.; Kang, Y.; Dang, V.T.; Stojanovski, D.; Coulson, B.S. Rotavirus NSP6 localizes to mitochondria via a predicted N-terminal a-helix. J. Gen. Virol. 2015, 96, 3519–3524. [Google Scholar] [CrossRef]
- Kar, A.K.; Bhattacharya, B.; Roy, P. Bluetongue virus RNA binding protein NS2 is a modulator of viral replication and assembly. BMC Mol. Biol. 2007, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, E.K.; Clavarino, G.; Ceppi, M.; Pierre, P. SUnSET, a nonradioactive method to monitor protein synthesis. Nat. Methods 2009, 6, 275–277. [Google Scholar] [CrossRef]
- David, A.; Dolan, B.P.; Hickman, H.D.; Knowlton, J.J.; Clavarino, G.; Pierre, P.; Bennink, J.R.; Yewdell, J.W. Nuclear translation visualized by ribosome-bound nascent chain puromycylation. J. Cell Biol. 2012, 197, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Huismans, H. Host cell protein synthesis after infection with bluetongue virus and reovirus. Virology 1971, 46, 500–503. [Google Scholar] [CrossRef]
- Huismans, H. Protein synthesis in bluetongue virus-infected cells. Virology 1979, 92, 385–396. [Google Scholar] [CrossRef]
- Vanecko, S.; Laskowski, M., Sr. Studies of the specificity of deoxyribonuclease I. II. Hydrolysis of oligonucleotides carrying a monoesterified phosphate on carbon 3′. J. Biol. Chem. 1961, 236, 1135–1140. [Google Scholar] [CrossRef]
- Lafer, E.M.; Sousa, R.; Rich, A. Anti-Z-DNA antibody binding can stabilize Z-DNA in relaxed and linear plasmids under physiological conditions. EMBO J. 1985, 4, 3655–3660. [Google Scholar] [CrossRef]
- Moll, J.R.; Acharya, A.; Gal, J.; Mir, A.A.; Vinson, C. Magnesium is required for specific DNA binding of the CREB B-ZIP domain. Nucleic Acids Res. 2002, 30, 1240–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuriakose, T.; Kanneganti, T.D. ZBP1: Innate Sensor Regulating Cell Death and Inflammation. Trends Immunol. 2018, 39, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, T.; Behlke, J.; Lowenhaupt, K.; Heinemann, U.; Rich, A. Structure of the DLM-1-Z-DNA complex reveals a conserved family of Z-DNA-binding proteins. Nat. Struct. Biol. 2001, 8, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Rothenburg, S.; Schwartz, T.; Koch-Nolte, F.; Haag, F. Complex regulation of the human gene for the Z-DNA binding protein DLM-1. Nucleic Acids Res. 2002, 30, 993–1000. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Comella, N.; Tognazzi, K.; Brown, L.F.; Dvorak, H.F.; Kocher, O. Cloning of DLM-1, a novel gene that is up-regulated in activated macrophages, using RNA differential display. Gene 1999, 240, 157–163. [Google Scholar] [CrossRef]
- Deigendesch, N.; Koch-Nolte, F.; Rothenburg, S. ZBP1 subcellular localization and association with stress granules is controlled by its Z-DNA binding domains. Nucleic Acids Res. 2006, 34, 5007–5020. [Google Scholar] [CrossRef] [Green Version]
- Pham, H.T.; Park, M.Y.; Kim, K.K.; Kim, Y.G.; Ahn, J.H. Intracellular localization of human ZBP1: Differential regulation by the Z-DNA binding domain, Zalpha, in splice variants. Biochem. Biophys. Res. Commun. 2006, 348, 145–152. [Google Scholar] [CrossRef]
- Salvetti, A.; Greco, A. Viruses and the nucleolus: The fatal attraction. Biochim. Biophys. Acta 2014, 1842, 840–847. [Google Scholar] [CrossRef]
- Mohd Jaafar, F.; Attoui, H.; Mertens, P.P.C.; de Micco, P.; de Lamballerie, X. Structural organization of an encephalitic human isolate of Banna virus (genus Seadornavirus, family Reoviridae). J. Gen. Virol. 2005, 86, 1147–1157. [Google Scholar] [CrossRef]
- Piron, M.; Vende, P.; Cohen, J.; Poncet, D. Rotavirus RNA-binding protein NSP3 interacts with eIF4GI and evicts the poly(A) binding protein from eIF4F. EMBO J. 1998, 17, 5811–5821. [Google Scholar] [CrossRef] [Green Version]
- Gratia, M.; Sarot, E.; Vende, P.; Charpilienne, A.; Baron, C.H.; Duarte, M.; Pyronnet, S.; Poncet, D. Rotavirus NSP3 Is a Translational Surrogate of the Poly(A) Binding Protein-Poly(A) Complex. J. Virol. 2015, 89, 8773–8782. [Google Scholar] [CrossRef] [Green Version]
- Matthews, D.A. Adenovirus protein V induces redistribution of nucleolin and B23 from nucleolus to cytoplasm. J. Virol. 2001, 75, 1031–1038. [Google Scholar] [CrossRef] [Green Version]
- Luijsterburg, M.S.; White, M.F.; van Driel, R.; Dame, R.T. The major architects of chromatin: Architectural proteins in bacteria, archaea and eukaryotes. Crit. Rev. Biochem. Mol. Biol. 2008, 43, 393–418. [Google Scholar] [CrossRef]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Maelfait, J.; Liverpool, L.; Bridgeman, A.; Ragan, K.B.; Upton, J.W.; Rehwinkel, J. Sensing of viral and endogenous RNA by ZBP1/DAI induces necroptosis. EMBO J. 2017, 36, 2529–2543. [Google Scholar] [CrossRef]
- Koehler, H.; Cotsmire, S.; Langland, J.; Kibler, K.V.; Kalman, D.; Upton, J.W.; Mocarski, E.S.; Jacobs, B.L. Inhibition of DAI-dependent necroptosis by the Z-DNA binding domain of the vaccinia virus innate immune evasion protein, E3. Proc. Natl. Acad. Sci. USA 2017, 114, 11506–11511. [Google Scholar] [CrossRef] [Green Version]
- Beattie, E.; Kauffman, E.B.; Martinez, H.; Perkus, M.E.; Jacobs, B.L.; Paoletti, E.; Tartaglia, J. Host-range restriction of vaccinia virus E3L-specific deletion mutants. Virus Genes 1996, 12, 89–94. [Google Scholar] [CrossRef]
- Shors, T.; Jacobs, B.L. Complementation of deletion of the vaccinia virus E3L gene by the Escherichia coli RNase III gene. Virology 1997, 227, 77–87. [Google Scholar] [CrossRef]
- Chang, H.W.; Uribe, L.H.; Jacobs, B.L. Rescue of vaccinia virus lacking the E3L gene by mutants of E3L. J. Virol. 1995, 69, 6605–6608. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.; Ding, T.; Zuo, Z.; Xu, Z.; Deng, J.; Wei, Z. Regulation of MAVS Expression and Signaling Function in the Antiviral Innate Immune Response. Front. Immunol. 2020, 11, 1030. [Google Scholar] [CrossRef]
- Ekanayaka, P.; Lee, B.H.; Weerawardhana, A.; Chathuranga, K.; Park, J.H.; Lee, J.S. Inhibition of MAVS Aggregation-Mediated Type-I Interferon Signaling by Foot-and-Mouth Disease Virus VP3. Viruses 2021, 13, 1776. [Google Scholar] [CrossRef]
- Ohta, A.; Nishiyama, Y. Mitochondria and viruses. Mitochondrion 2011, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mibayashi, M.; Martinez-Sobrido, L.; Loo, Y.M.; Cardenas, W.B.; Gale, M., Jr.; Garcia-Sastre, A. Inhibition of retinoic acid-inducible gene I-mediated induction of beta interferon by the NS1 protein of influenza A virus. J. Virol. 2007, 81, 514–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pourcelot, M.; Amaral Moraes, R.; Fablet, A.; Breard, E.; Sailleau, C.; Viarouge, C.; Postic, L.; Zientara, S.; Caignard, G.; Vitour, D. The VP3 Protein of Bluetongue Virus Associates with the MAVS Complex and Interferes with the RIG-I-Signaling Pathway. Viruses 2021, 13, 230. [Google Scholar] [CrossRef] [PubMed]
- Attoui, H.; Monsion, B.; Klonjkowski, B.; Zientara, S.; Mertens, P.P.C.; Mohd Jaafar, F. Identification of the Genome Segments of Bluetongue Virus Type 26/Type 1 Reassortants Influencing Horizontal Transmission in a Mouse Model. Viruses 2021, 13, 2208. [Google Scholar] [CrossRef] [PubMed]
- Attoui, H.; Billoir, F.; Cantaloube, J.F.; Biagini, P.; de Micco, P.; de Lamballerie, X. Strategies for the sequence determination of viral dsRNA genomes. J. Virol. Methods 2000, 89, 147–158. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Bernhofer, M.; Dallago, C.; Karl, T.; Satagopam, V.; Heinzinger, M.; Littmann, M.; Olenyi, T.; Qiu, J.; Schutze, K.; Yachdav, G.; et al. PredictProtein—Predicting Protein Structure and Function for 29 Years. Nucleic Acids Res. 2021, 49, W535–W540. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Xu, J. RaptorX: Exploiting structure information for protein alignment by statistical inference. Proteins 2011, 79 (Suppl. S10), 161–171. [Google Scholar] [CrossRef] [Green Version]
- Schneider, K.; Zimmer, D.; Nielsen, H.; Herrmann, J.M.; Muhlhaus, T. iMLP, a predictor for internal matrix targeting-like sequences in mitochondrial proteins. Biol. Chem. 2021, 402, 937–943. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Maan, S.; Rao, S.; Maan, N.S.; Anthony, S.J.; Attoui, H.; Samuel, A.R.; Mertens, P.P. Rapid cDNA synthesis and sequencing techniques for the genetic study of bluetongue and other dsRNA viruses. J. Virol. Methods 2007, 143, 132–139. [Google Scholar] [CrossRef]
- Engler, C.; Kandzia, R.; Marillonnet, S. A one pot, one step, precision cloning method with high throughput capability. PLoS ONE 2008, 3, e3647. [Google Scholar] [CrossRef] [Green Version]
- Mohd Jaafar, F.; Attoui, H.; Gallian, P.; Biagini, P.; Cantaloube, J.F.; de Micco, P.; de Lamballerie, X. Recombinant VP7-based enzyme-linked immunosorbent assay for detection of immunoglobulin G antibodies to Colorado tick fever virus. J. Clin. Microbiol. 2003, 41, 2102–2105. [Google Scholar] [CrossRef] [Green Version]
- Pullinger, G.D.; Guimera Busquets, M.; Nomikou, K.; Boyce, M.; Attoui, H.; Mertens, P.P. Identification of the Genome Segments of Bluetongue Virus Serotype 26 (Isolate KUW2010/02) that Restrict Replication in a Culicoides sonorensis Cell Line (KC Cells). PLoS ONE 2016, 11, e0149709. [Google Scholar] [CrossRef] [Green Version]
- Mendez, I.I.; Hermann, L.L.; Hazelton, P.R.; Coombs, K.M. A comparative analysis of freon substitutes in the purification of reovirus and calicivirus. J. Virol. Methods 2000, 90, 59–67. [Google Scholar] [CrossRef]
- Mohd Jaafar, F.; Belhouchet, M.; Vitour, D.; Adam, M.; Breard, E.; Zientara, S.; Mertens, P.P.; Attoui, H. Immunisation with bacterial expressed VP2 and VP5 of bluetongue virus (BTV) protect alpha/beta interferon-receptor knock-out (IFNAR(-/-)) mice from homologous lethal challenge. Vaccine 2014, 32, 4059–4067. [Google Scholar] [CrossRef]
- Hofmann, M.; Griot, C.; Chaignat, V.; Perler, L.; Thur, B. Bluetongue disease reaches Switzerland. Schweiz. Arch. Tierheilkd. 2008, 150, 49–56. [Google Scholar] [CrossRef]
- Hofmann, M.A.; Renzullo, S.; Mader, M.; Chaignat, V.; Worwa, G.; Thuer, B. Genetic characterization of toggenburg orbivirus, a new bluetongue virus, from goats, Switzerland. Emerg. Infect. Dis. 2008, 14, 1855–1861. [Google Scholar] [CrossRef]
- Attoui, H.; Billoir, F.; Bruey, J.M.; de Micco, P.; de Lamballerie, X. Serologic and molecular diagnosis of Colorado tick fever viral infections. Am. J. Trop. Med. Hyg. 1998, 59, 763–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]



















| (a) | |||||||
|---|---|---|---|---|---|---|---|
| Infection with BTV-1ΔNS5 | Normalized Mean Ct Values for Non-Transfected (Ctrl) and Transfected Cells | Standard Deviation | |||||
| Experiment | Viral RNA Detected | Ctrl | NS5 (AHSV) | NS5 (BTV) | Ctrl | NS5 (AHSV) | NS5 (BTV) |
| I | BTV-1 Seg-1 | 18.09 | 11.61 | 10.94 | 0.16 | 0.06 | 0.03 |
| BTV-1 Seg-3 | 19.87 | 11.79 | 11.46 | 0.28 | 0.03 | 0.15 | |
| BTV-1 Seg-7 | 16.39 | 10.94 | 10.25 | 0.16 | 0.08 | 0.08 | |
| II | BTV-1 Seg-1 | 14.81 | 11.28 | 10.80 | 0.14 | 0.07 | 0.19 |
| BTV-1 Seg-3 | 19.32 | 11.87 | 11.16 | 0.41 | 0.12 | 0.09 | |
| BTV-1 Seg-7 | 16.17 | 11.76 | 11.05 | 0.18 | 0.06 | 0.04 | |
| (b) | |||||||
| Infection with BTV-1RGC7 | Normalized Mean Ct Values for Non-Transfected (Ctrl) and Transfected Cells | Standard Deviation | |||||
| Viral RNA Detected | Ctrl | NS5 (AHSV) | NS5 (BTV) | NS5 (AHSV) | NS5 (BTV) | ||
| BTV-1 Seg-1 | 17.80 | 17.60 | 18.01 | 0.82 | 0.08 | ||
| BTV-1 Seg-3 | 17.02 | 16.17 | 15.86 | 0.36 | 0.13 | ||
| BTV-1 Seg-7 | 18.63 | 17.24 | 17.02 | 0.16 | 0.18 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohd Jaafar, F.; Monsion, B.; Mertens, P.P.C.; Attoui, H. Identification of Orbivirus Non-Structural Protein 5 (NS5), Its Role and Interaction with RNA/DNA in Infected Cells. Int. J. Mol. Sci. 2023, 24, 6845. https://doi.org/10.3390/ijms24076845
Mohd Jaafar F, Monsion B, Mertens PPC, Attoui H. Identification of Orbivirus Non-Structural Protein 5 (NS5), Its Role and Interaction with RNA/DNA in Infected Cells. International Journal of Molecular Sciences. 2023; 24(7):6845. https://doi.org/10.3390/ijms24076845
Chicago/Turabian StyleMohd Jaafar, Fauziah, Baptiste Monsion, Peter P. C. Mertens, and Houssam Attoui. 2023. "Identification of Orbivirus Non-Structural Protein 5 (NS5), Its Role and Interaction with RNA/DNA in Infected Cells" International Journal of Molecular Sciences 24, no. 7: 6845. https://doi.org/10.3390/ijms24076845
APA StyleMohd Jaafar, F., Monsion, B., Mertens, P. P. C., & Attoui, H. (2023). Identification of Orbivirus Non-Structural Protein 5 (NS5), Its Role and Interaction with RNA/DNA in Infected Cells. International Journal of Molecular Sciences, 24(7), 6845. https://doi.org/10.3390/ijms24076845