
Neurotrophins: Expression of Brain–Lung Axis Development

 ,
, 


Abstract
:1. Introduction
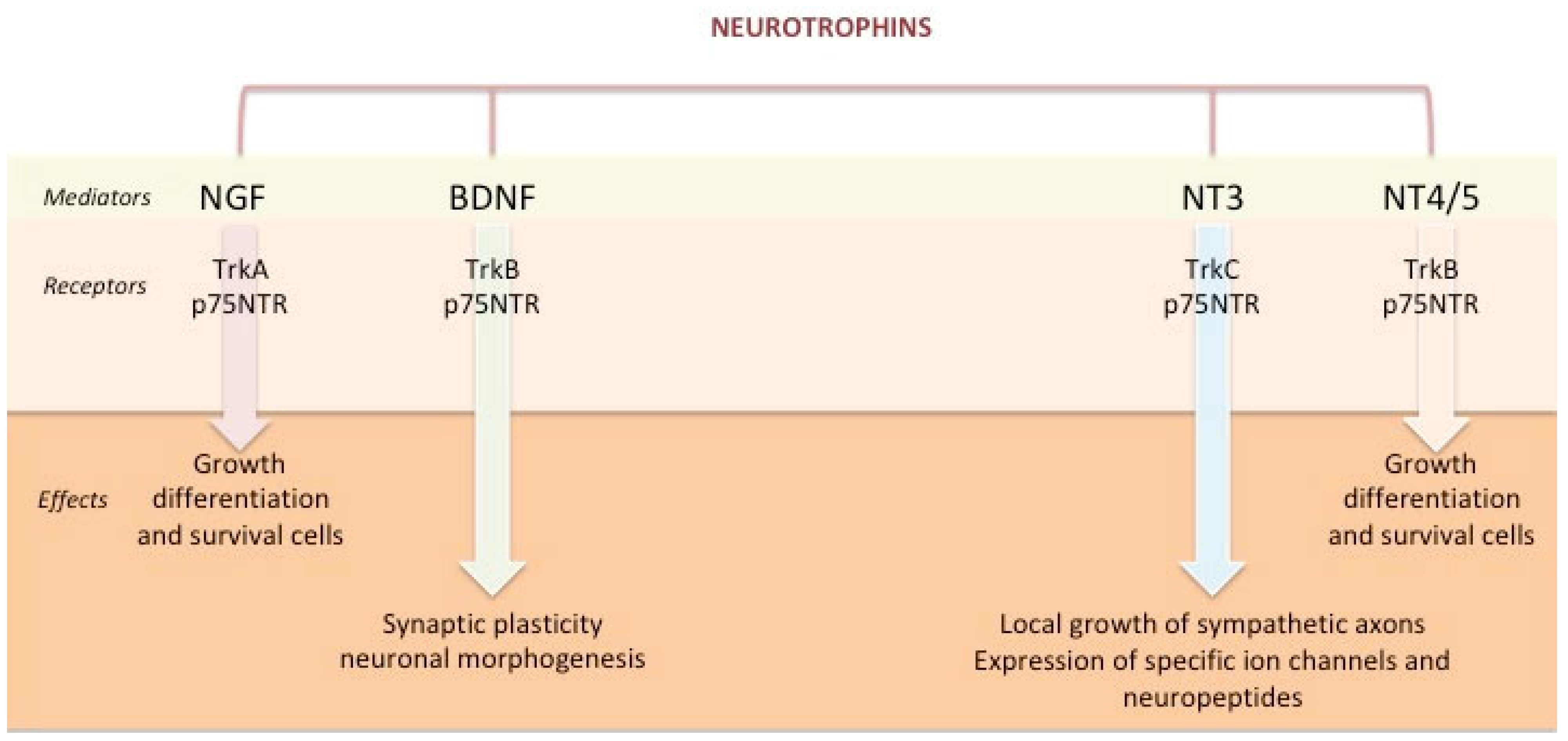
2. Neurotrophins: Role and Characteristics
3. Neurotrophins and Brain
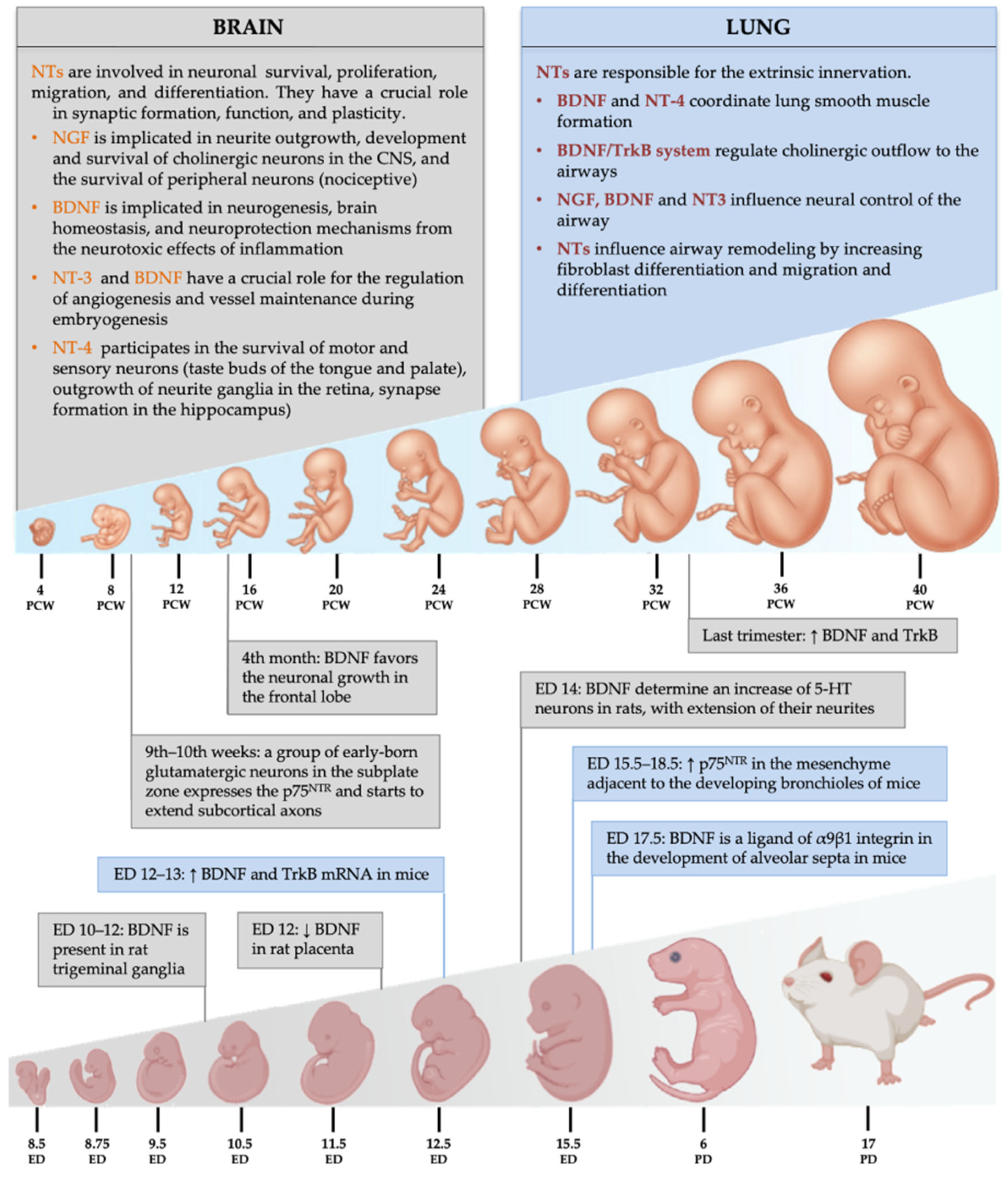
3.1. Neurotrophins and Brain Development
3.2. Neutrophins and Neurologic Diseases
4. Neurotrophins and Lung
4.1. Neurotrophins and Lung Development
4.2. Neurotrophins Expression in the Peripheral Tissues
4.3. Neurotrophins and Lung Diseases
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Prakash, Y.; Thompson, M.A.; Meuchel, L.; Pabelick, C.M.; Mantilla, C.B.; Zaidi, S.; Martin, R.J. Neurotrophins in lung health and disease. Expert Rev. Respir. Med. 2010, 4, 395–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubin, L.; Stabler, C.T.; Schumacher-Klinger, A.; Marcinkiewicz, C.; Lelkes, P.I.; Lazarovici, P. Neurotrophic factors and their receptors in lung development and implications in lung diseases. Cytokine Growth Factor Rev. 2021, 59, 84–94. [Google Scholar] [CrossRef]
- Prakash, Y.; Martin, R.J. Brain-derived neurotrophic factor in the airways. Pharmacol. Ther. 2014, 143, 74–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartkowska, K.; Turlejski, K.; Djavadian, R.L. Neurotrophins and their receptors in early development of the mammalian nervous system. Acta Neurobiol. Exp. 2010, 70, 454–467. [Google Scholar]
- Bothwell, M. NGF, BDNF, NT3, and NT4. Organotypic Models Drug Dev. 2014, 220, 3–15. [Google Scholar] [CrossRef]
- Pejhan, S.; Siu, V.M.; Ang, L.C.; Del Bigio, M.R.; Rastegar, M. Differential brain region-specific expression of MeCP2 and BDNF in Rett Syndrome patients: A distinct grey-white matter variation. Neuropathol. Appl. Neurobiol. 2020, 46, 735–750. [Google Scholar] [CrossRef]
- Purves, D.; Augustine, G.J.; Fitzpatrick, D. Neurotransmitter Synthesis. In Book Neuroscience, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2001. Available online: https://www.ncbi.nlm.nih.gov/books/NBK11110/ (accessed on 23 March 2023).
- Hu, Y.; Russek, S.J. BDNF and the diseased nervous system: A delicate balance between adaptive and pathological processes of gene regulation. J. Neurochem. 2008, 105, 1–17. [Google Scholar] [CrossRef]
- Liu, P.; Li, S.; Tang, L. Nerve Growth Factor: A Potential Therapeutic Target for Lung Diseases. Int. J. Mol. Sci. 2021, 22, 9112. [Google Scholar] [CrossRef]
- Skaper, S.D. The Biology of Neurotrophins, Signalling Pathways, and Functional Peptide Mimetics of Neurotrophins and their Receptors. CNS Neurol. Disord. Drug Targets 2008, 7, 46–62. [Google Scholar] [CrossRef]
- Underwood, C.K.; Coulson, E.J. The p75 neurotrophin receptor. Int. J. Biochem. Cell Biol. 2008, 40, 1664–1668. [Google Scholar] [CrossRef]
- Reichardt, L.F. Neurotrophin-regulated signalling pathways. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1545–1564. [Google Scholar] [CrossRef] [Green Version]
- Barbacid, M. Structural and functional properties of the TRK family of neurotrophin receptors. Ann. N. Y. Acad. Sci. 1995, 766, 442–458. [Google Scholar] [CrossRef]
- Chao, M.V. Neurotrophins and their receptors: A convergence point for many signalling pathways. Nat. Rev. Neurosci. 2003, 4, 299–309. [Google Scholar] [CrossRef]
- Hempstead, B.L. The many faces of p75NTR. Curr. Opin. Neurobiol. 2002, 12, 260–267. [Google Scholar] [CrossRef]
- Manchia, M.; Isayeva, U.; Collu, R.; Primavera, D.; Deriu, L.; Caboni, E.; Iaselli, M.N.; Sundas, D.; Tusconi, M.; Pinna, F.; et al. Converging Evidence Points to BDNF as Biomarker of Depressive Symptoms in Schizophrenia-Spectrum Disorders. Brain Sci. 2022, 12, 1666. [Google Scholar] [CrossRef]
- Huang, E.J.; Reichardt, L.F. Trk receptors: Roles in neuronal signal transduction. Annu. Rev. Biochem. 2003, 72, 609–642. [Google Scholar] [CrossRef] [Green Version]
- Levi-Montalcini, R. The nerve growth factor. Ann. N. Y. Acad. Sci. 2006, 118, 149–170. [Google Scholar] [CrossRef]
- Segal, R.A. Selectivity in neurotrophin signaling: Theme and Variations. Annu. Rev. Neurosci. 2003, 26, 299–330. [Google Scholar] [CrossRef]
- Krey, F.C.; Stocchero, B.A.; Creutzberg, K.C.; Heberle, B.A.; Tractenberg, S.G.; Xiang, L.; Wei, W.; Kluwe-Schiavon, B.; Viola, T.W. Neurotrophic Factor Levels in Preterm Infants: A Systematic Review and Meta-Analysis. Front. Neurol. 2021, 12, 643576. [Google Scholar] [CrossRef]
- Yao, Q.; Zaidi, S.I.; Haxhiu, M.A.; Martin, R.J. Neonatal Lung and Airway Injury: A Role for Neurotrophins. Semin. Perinatol. 2006, 30, 156–162. [Google Scholar] [CrossRef]
- Zagrebelsky, M.; Tacke, C.; Korte, M. BDNF signaling during the lifetime of dendritic spines. Cell Tissue Res. 2020, 382, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Bystron, I.; Blakemore, C.; Rakic, P. Development of the human cerebral cortex: Boulder Committee revisited. Nat. Rev. Neurosci. 2008, 9, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Bernd, P. The Role of Neurotrophins During Early Development. Gene Expr. 2008, 14, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Davies, A.M. The emerging generality of the neurotrophic hypothesis. Trends Neurosci. 1988, 11, 243–244. [Google Scholar] [CrossRef] [PubMed]
- Markus, A.; Zhong, J.; Snider, W.D. Raf and Akt Mediate Distinct Aspects of Sensory Axon Growth. Neuron 2002, 35, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Sahay, A.; Kale, A.; Joshi, S. Role of neurotrophins in pregnancy and offspring brain development. Neuropeptides 2020, 83, 102075. [Google Scholar] [CrossRef]
- Lewin, G.R. Neurotrophins and the specification of neuronal phenotype. Philos. Trans. R. Soc. B Biol. Sci. 1996, 351, 405–411. [Google Scholar] [CrossRef]
- Antonakopoulos, N.; Iliodromiti, Z.; Mastorakos, G.; Iavazzo, C.; Valsamakis, G.; Salakos, N.; Papageorghiou, A.; Margeli, A.; Kalantaridou, S.; Creatsas, G.; et al. Association between Brain-Derived Neurotrophic Factor (BDNF) Levels in 2nd Trimester Amniotic Fluid and Fetal Development. Mediat. Inflamm. 2018, 2018, 8476217. [Google Scholar] [CrossRef] [Green Version]
- Benn, K.; Passos, M.; Jayaram, A.; Harris, M.; Bongiovanni, A.M.; Skupski, D.; Witkin, S.S. Association Between Neurotrophin 4 and Long-Chain Polyunsaturated Fatty Acid Levels in Mid-Trimester Amniotic Fluid. Reprod. Sci. 2014, 21, 1395–1400. [Google Scholar] [CrossRef]
- Christian, L.M.; Mitchell, A.M.; Gillespie, S.L.; Palettas, M. Serum brain-derived neurotrophic factor (BDNF) across pregnancy and postpartum: Associations with race, depressive symptoms, and low birth weight. Psychoneuroendocrinology 2016, 74, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Gumus, C.; Yazici, I.P.; Yazici, K.U.; Ustundag, B. Increased Serum Brain-derived Neurotrophic Factor, Nerve Growth Factor, Glial-derived Neurotrophic Factor and Galanin Levels in Children with Attention Deficit Hyperactivity Disorder, and the Effect of 10 Weeks Methylphenidate Treatment. Clin. Psychopharmacol. Neurosci. 2022, 20, 635–648. [Google Scholar] [CrossRef]
- Li, W.; Pozzo-Miller, L. BDNF deregulation in Rett syndrome. Neuropharmacology 2014, 76, 737–746. [Google Scholar] [CrossRef] [Green Version]
- Nelson, K.B.; Grether, J.K.; Croen, L.A.; Dambrosia, J.M.; Dickens, B.F.; Jelliffe, L.L.; Hansen, R.L.; Phillips, T.M. Neuropeptides and neurotrophins in neonatal blood of children with autism or mental retardation. Ann. Neurol. 2001, 49, 597–606. [Google Scholar] [CrossRef]
- Sajdel-Sulkowska, E.M.; Xu, M.; Koibuchi, N. Increase in Cerebellar Neurotrophin-3 and Oxidative Stress Markers in Autism. Cerebellum 2009, 8, 366–372. [Google Scholar] [CrossRef]
- Sgritta, M.; Vignoli, B.; Pimpinella, D.; Griguoli, M.; Santi, S.; Bialowas, A.; Wiera, G.; Zacchi, P.; Malerba, F.; Marchetti, C.; et al. Impaired synaptic plasticity in an animal model of autism exhibiting early hippocampal GABAergic-BDNF/TrkB signaling alterations. iScience 2022, 26, 105728. [Google Scholar] [CrossRef]
- Gevezova, M.; Minchev, D.; Pacheva, I.; Todorova, T.; Yordanova, R.; Timova, E.; Ivanov, I.; Sarafian, V. Association of NGF and Mitochondrial Respiration with Autism Spectrum Disorder. Int. J. Mol. Sci. 2022, 23, 11917. [Google Scholar] [CrossRef]
- Camuso, S.; La Rosa, P.; Fiorenza, M.T.; Canterini, S. Pleiotropic effects of BDNF on the cerebellum and hippocampus: Implications for neurodevelopmental disorders. Neurobiol. Dis. 2022, 163, 105606. [Google Scholar] [CrossRef]
- Francis, K.; Dougali, A.; Sideri, K.; Kroupis, C.; Vasdekis, V.; Dima, K.; Douzenis, A. Brain-derived neurotrophic factor (BDNF) in children with ASD and their parents: A 3-year follow-up. Acta Psychiatr. Scand. 2018, 137, 433–441. [Google Scholar] [CrossRef]
- Bryn, V.; Halvorsen, B.; Ueland, T.; Isaksen, J.; Kolkova, K.; Ravn, K.; Skjeldal, O. Brain derived neurotrophic factor (BDNF) and autism spectrum disorders (ASD) in childhood. Eur. J. Paediatr. Neurol. EJPN 2015, 19, 411–414. [Google Scholar] [CrossRef]
- Alleva, E.; Santucci, D. Psychosocial vs. “physical” stress situations in rodents and humans: Role of neurotrophins. Physiol. Behav. 2001, 73, 313–320. [Google Scholar] [CrossRef]
- Miao, Z.; Wang, Y.; Sun, Z. The Relationships Between Stress, Mental Disorders, and Epigenetic Regulation of BDNF. Int. J. Mol. Sci. 2020, 21, 1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, Z.; Mao, F.; Liang, J.; Szyf, M.; Wang, Y.; Sun, Z.S. Anxiety-Related Behaviours Associated with microRNA-206-3p and BDNF Expression in Pregnant Female Mice Following Psychological Social Stress. Mol. Neurobiol. 2018, 55, 1097–1111. [Google Scholar] [CrossRef] [PubMed]
- Erickson, J.T.; Conover, J.C.; Borday, V.; Champagnat, J.; Barbacid, M.; Yancopoulos, G.; Katz, D.M. Mice Lacking Brain-Derived Neurotrophic Factor Exhibit Visceral Sensory Neuron Losses Distinct from Mice Lacking NT4 and Display a Severe Developmental Deficit in Control of Breathing. J. Neurosci. 1996, 16, 5361–5371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crystal, R.G.; Randell, S.H.; Engelhardt, J.F.; Voynow, J.; Sunday, M.E. Airway Epithelial Cells: Current Concepts and Challenges. Proc. Am. Thorac. Soc. 2008, 5, 772–777. [Google Scholar] [CrossRef]
- Manti, S.; Piedimonte, G. An overview on the RSV-mediated mechanisms in the onset of non-allergic asthma. Front. Pediatr. 2022, 10, 998296. [Google Scholar] [CrossRef]
- Fox, A.J.; Patel, H.J.; Barnes, P.J.; Belvisi, M.G. Release of nerve growth factor by human pulmonary epithelial cells: Role in airway infammatory diseases. Eur. J. Pharmacol. 2001, 424, 159–162. [Google Scholar] [CrossRef]
- García-Suárez, O.; Pérez-Pinera, P.; Laurà, R.; Germana, A.; Esteban, I.; Cabo, R.; Silos-Santiago, I.; Cobo, J.; Vega, J. TrkB is necessary for the normal development of the lung. Respir. Physiol. Neurobiol. 2009, 167, 281–291. [Google Scholar] [CrossRef]
- Radzikinas, K.L.; Aven, Z.; Jiang, Z.; Tran, T.; Paez-Cortez, K.; Boppidi, K.; Lu, J.; Fine, A.; Ai, X. A Shh/miR-206/BDNF cascade coordinates innervation and formation of airway smooth muscle. J. Neurosci. 2011, 31, 15407–15415. [Google Scholar] [CrossRef] [Green Version]
- Giblin, S.P.; Midwood, K.S. Tenascin-C: Form versus function. Cell Adhes. Migr. 2014, 9, 48–82. [Google Scholar] [CrossRef] [Green Version]
- Prakash, Y.S.; Iyanoye, A.; Ay, B.; Mantilla, C.B.; Pabelick, C.M. Neurotrophin effects on in-tracellular Ca2+ and force in airway smooth muscle. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 291, L447–L456. [Google Scholar] [CrossRef] [Green Version]
- Bai, T.R. Evidence for airway remodeling in chronic asthma. Curr. Opin. Allergy Clin. Immunol. 2010, 10, 82–86. [Google Scholar] [CrossRef]
- Olgart, C.; Frossard, N. Human lung fibroblasts secrete nerve growth factor: Effect of inflammatory cytokines and glucocorticoids. Eur. Respir. J. 2001, 18, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Bahat-Stroomza, M.; Barhum, Y.; Levy, Y.S.; Karpov, O.; Bulvik, S.; Melamed, E.; Offen, D. Induction of Adult Human Bone Marrow Mesenchymal Stromal Cells into Functional Astrocyte-Like Cells: Potential for Restorative Treatment in Parkinson’s Disease. J. Mol. Neurosci. 2009, 39, 199–210. [Google Scholar] [CrossRef]
- Ricci, A.; Graziano, P.; Bronzetti, E.; Saltini, C.; Sciacchitano, S.; Cherubini, E.; Renzoni, E.; Du Bois, R.M.; Grutters, J.C.; Mariotta, S. Increased pulmonary neurotrophin protein in idiopathic interstitial pneumonias. Sarcoidosis Vasc. Diffuse Lung Dis. 2007, 24, 13–23. [Google Scholar]
- Antonelli, A.; Lapucci, G.; Vigneti, E.; Bonini, S.; Aloe, L. Human Lung Fibroblast Response to NGF, IL-1β, and Dexamethsone. Lung 2005, 183, 337–351. [Google Scholar] [CrossRef]
- Braun, A.; Lommatzsch, M.; Neuhaus-Steinmetz, U.; Quarcoo, D.; Glaab, T.; McGregor, G.P.; Fischer, A.; Renz, H. Brain-derived neurotrophic factor (BDNF) contributes to neuronal dysfunction in a model of allergic airway inflammation. Br. J. Pharmacol. 2004, 141, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Braun, A.; Lommatzsch, M.; Lewin, G.R.; Virchow, J.C.; Renz, H. Neurotrophins: A link between airway inflammation and airway smooth muscle contractility in asthma? Int. Arch. Allergy Immunol. 1999, 118, 163–165. [Google Scholar] [CrossRef]
- Virchow, J.C.; Julius, P.; Lommatzsch, M.; Luttmann, W.; Renz, H.; Braun, A. Neurotrophins Are Increased in Bronchoalveolar Lavage Fluid after Segmental Allergen Provocation. Am. J. Respir. Crit. Care Med. 1998, 158, 2002–2005. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, E.; Gong, H.; Grimes, R.; Benoit, D.; Vazquez, L. p75NTR andTrk receptors are expressed in reciprocal patterns in a wide variety of non-neural tissues during rat embryonic development, indicating independent receptor functions. J. Comp. Neurol. 1998, 391, 407–428. [Google Scholar] [CrossRef]
- Weinacker, A.; Ferrando, R.; Elliott, M.; Hogg, J.; Balmes, J.; Sheppard, D. Distribution of integrins alpha v beta 6 and alpha 9 beta 1 and their known ligands, fibronectin and tenascin, in human airways. Am. J. Respir. Cell Mol. Biol. 1995, 12, 547–556. [Google Scholar] [CrossRef]
- Kerschensteiner, M.; Gallmeier, E.; Behrens, L.; Leal, V.V.; Misgeld, T.; Klinkert, W.E.F.; Kolbeck, R.; Hoppe, E.; Oropeza-Wekerle, R.-L.; Bartke, I.; et al. Activated Human T Cells, B Cells, and Monocytes Produce Brain-derived Neurotrophic Factor In Vitro and in Inflammatory Brain Lesions: A Neuroprotective Role of Inflammation? J. Exp. Med. 1999, 189, 865–870. [Google Scholar] [CrossRef] [PubMed]
- Santambrogio, L.; Benedetti, M.; Chao, M.V.; Muzaffar, R.; Kulig, K.; Gabellini, N.; Hochwald, G. Nerve growth factor production by lymphocytes. J. Immunol. 1994, 153, 4488–4495. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Gleich, G.J.; Butterfield, J.H.; Kita, H. Human eosinophils produce neurotro-phins and secrete nerve growth factor on immunologic stimuli. Blood 2002, 99, 2214–2220. [Google Scholar] [CrossRef] [PubMed]
- Noga, O.; Englmann, C.; Hanf, G.; Grützkau, A.; Guhl, S.; Kunkel, G. Activation of the specific neurotrophin receptors TrkA, TrkB and TrkC influences the function of eosinophils. Clin. Exp. Allergy 2002, 32, 1348–1354. [Google Scholar] [CrossRef]
- Sonar, S.S.; Schwinge, D.; Kilic, A.; Yildirim, A.O.; Conrad, M.L.; Seidler, K.; Muller, B.; Renz, H.; Nockher, W.A. Nerve growth factor enhances Clara cell proliferation after lung injury. Eur. Respir. J. 2010, 36, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Meuchel, L.; Townsend, E.; Thompson, M.; Pabelick, C.; Prakash, Y. Effect of neurotrophins on NO generation in airway epi-thelial cells. In Proceedings of the International Conference of the American Thoracic Society, New Orleans, LA, USA, 14–19 May 2010. [Google Scholar]
- Racké, K.; Matthiesen, S. The airway cholinergic system: Physiology and pharmacology. Pulm. Pharmacol. Ther. 2004, 17, 181–198. [Google Scholar] [CrossRef]
- Piedimonte, G. Contribution of neuroimmune mechanisms to airway inflammation and remodeling during and after respiratory syncytial virus infection. Pediatr. Infect. Dis. J. 2003, 22 (Suppl. S2), S66–S74. [Google Scholar] [CrossRef]
- Hoyle, G.W.; Graham, R.M.; Finkelstein, J.B.; Nguyen, K.-P.T.; Gozal, D.; Friedman, M. Hyperinnervation of the Airways in Transgenic Mice Overexpressing Nerve Growth Factor. Am. J. Respir. Cell Mol. Biol. 1998, 18, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Manti, S.; Brown, P.; Perez, M.K.; Piedimonte, G. The Role of Neurotrophins in Inflammation and Allergy. Vitam. Horm. 2017, 104, 313–341. [Google Scholar] [CrossRef]
- Xu, M.; Remillard, C.V.; Sachs, B.D.; Makino, A.; Platoshyn, O.; Yao, W.; Dillmann, W.H.; Akassoglou, K.; Yuan, J.X.-J. p75 neurotrophin receptor regulates agonist-induced pulmonary vasoconstriction. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1529–H1538. [Google Scholar] [CrossRef] [Green Version]
- Ricci, A.; Greco, S.; Amenta, F.; Bronzetti, E.; Felici, L.; Rossodivita, I.; Sabbatini, M.; Mariotta, S. Neurotrophins and Neurotrophin Receptors in Human Pulmonary Arteries. J. Vasc. Res. 2000, 37, 355–363. [Google Scholar] [CrossRef]
- Meuchel, L.; Thompson, M.; Pabelick, C.; Prakash, Y. Neurotrophins induce NO generation in pulmonary artery endothelial cells. In Proceedings of the Experimental Biology, Anaheim, CA, USA, 24–28 April 2010. [Google Scholar]
- Caporali, A.; Emanueli, C. Cardiovascular Actions of Neurotrophins. Physiol. Rev. 2009, 89, 279–308. [Google Scholar] [CrossRef] [Green Version]
- Naeem, A.; Ahmed, I.; Silveyra, P. Bronchopulmonary Dysplasia: An Update on Ex-perimental Therapeutics. Eur. Med. J. 2019, 4, 20–29. [Google Scholar] [CrossRef]
- Manti, S.; Galdo, F.; Parisi, G.F.; Napolitano, M.; Decimo, F.; Leonardi, S.; Miraglia Del Giudice, M. Long-term effects of bronchopulmonary dys-plasia on lung function: A pilot study in preschool children’s cohort. J. Asthma 2021, 58, 1186–1193. [Google Scholar] [CrossRef]
- Wang, S.-H.; Tsao, P.-N. Phenotypes of Bronchopulmonary Dysplasia. Int. J. Mol. Sci. 2020, 21, 6112. [Google Scholar] [CrossRef]
- Holzfurtner, L.; Shahzad, T.; Dong, Y.; Rekers, L.; Selting, A.; Staude, B.; Lauer, T.; Schmidt, A.; Rivetti, S.; Zimmer, K.-P.; et al. When inflammation meets lung development—An update on the pathogenesis of bronchopulmonary dysplasia. Mol. Cell. Pediatr. 2022, 9, 7. [Google Scholar] [CrossRef]
- Sopi, R.B.; Martin, R.J.; Haxhiu, M.A.; Dreshaj, I.A.; Yao, Q.; Jafri, A.; Zaidi, S.I. Role of brain-derived neurotrophic factor in hy-peroxia-induced enhancement of contractility and impairment of relaxation in lung pa-renchyma. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 295, L348–L355. [Google Scholar] [CrossRef]
- Pulvirenti, G.; Parisi, G.F.; Giallongo, A.; Papale, M.; Manti, S.; Savasta, S.; Licari, A.; Marseglia, G.L.; Leonardi, S. Lower Airway Microbiota. Front. Pediatr. 2019, 7, 393. [Google Scholar] [CrossRef] [Green Version]
- Leonardi, S.; Cuppari, C.; Lanzafame, A.; Attardo, D.; Tardino, L.; Parisi, G.; Giacchi, V.; Manti, S.; Arrigo, T. Exhaled breath temperature in asthmatic children. J. Biol. Regul. Homeost. Agents 2015, 29 (Suppl. S1), 47–54. [Google Scholar]
- GINA 2019. Available online: https://ginasthma.org/wp-content/uploads/2019/01/2018-GINA.pdf (accessed on 21 January 2023).
- Prakash, Y.S.; Thompson, M.A.; Pabelick, C.M. Brain-Derived Neurotrophic Factor in TNF-α Modulation of Ca2+ in Human Airway Smooth Muscle. Am. J. Respir. Cell Mol. Biol. 2009, 41, 603–611. [Google Scholar] [CrossRef] [Green Version]
- Krüttgen, A.; Schneider, I.; Weis, J. The Dark Side of the NGF Family: Neurotrophins in Neoplasias. Brain Pathol. 2006, 16, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Ricci, A.; Greco, S.; Mariotta, S.; Felici, L.; Bronzetti, E.; Cavazzana, A.; Cardillo, G.; Amenta, F.; Bisetti, A.; Barbolini, G. Neurotrophins and Neurotrophin Receptors in Human Lung Cancer. Am. J. Respir. Cell Mol. Biol. 2001, 25, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Nico, B.; Mangieri, D.; Benagiano, V.; Crivellato, E.; Ribatti, D. Nerve growth factor as an angiogenic factor. Microvasc. Res. 2008, 75, 135–141. [Google Scholar] [CrossRef]
- Spoto, G.; Valentini, G.; Saia, M.C.; Butera, A.; Amore, G.; Salpietro, V.; Nicotera, A.G.; Di Rosa, G. Synaptopathies in Developmental and Epileptic Encephalopathies: A Focus on Pre-synaptic Dysfunction. Front. Neurol. 2022, 13, 826211. [Google Scholar] [CrossRef]
- Amore, G.; Spoto, G.; Ieni, A.; Vetri, L.; Quatrosi, G.; Di Rosa, G.; Nicotera, A.G. A Focus on the Cerebellum: From Embryogenesis to an Age-Related Clinical Perspective. Front. Syst. Neurosci. 2021, 15, 646052. [Google Scholar] [CrossRef]
- Skogstrand, K.; Hagen, C.M.; Borbye-Lorenzen, N.; Christiansen, M.; Bybjerg-Grauholm, J.; Bækvad-Hansen, M.; Werge, T.; Børglum, A.; Mors, O.; Nordentoft, M.; et al. Reduced neonatal brain-derived neurotrophic factor is associated with autism spectrum disorders. Transl. Psychiatry 2019, 9, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Neurotrophin | Receptor | Brain Region | Effects | Implication in Diseases | References |
|---|---|---|---|---|---|
| BDNF | TrkB | Hippocampus | Promotion of mossy fibers sprouting of the dentate granule cells | ↓ BDNF in deficits of learning and memory ↓ BDNF in ASD ↑ BDNF in seizure activity in temporal lobe epilepsy | [8,16,30,32,36,38,39,40] |
| TrkB/ P75NTR | Brainstem and cerebellum | Maturation of GABAergic neurons and development of synapses | ↓ BDNF in RTT syndrome | ||
| NA | HPA axis | Survival and homeostasis of dopaminergic neurons | Controversial data on BDNF in ADHD | ||
| TrkB/ P75NTR | Frontal cortices and hippocampus | Survival, proliferation, differentiation, and synaptic plasticity | ↓ BDNF in schizophrenia ↓ BDNF in depression | ||
| TrkB | CNS | Survival, proliferation, differentiation, and synaptic plasticity | ↑ BDNF in neonatal blood of children with ASD and ID | ||
| NGF | TrkA | Basal forebrain and prefrontal cortex | Survival of cholinergic neurons and involvement in learning processes and attention systems | ↑ NGF in ADHD | [37] |
| TrkA/ P75NTR | CNS | Homeostasis of mitochondria | ↑ NGF in ASD | ||
| NT-3 | NA | Cerebellum | Survival of neurons and cerebellar growth | ↑ NT-4 in ASD | [27] |
| NT-4 | TrkB | CNS | Survival, proliferation, differentiation, and synaptic plasticity | ↑ NT-4 in neonatal blood of children with ASD and ID | [30] |
| Airways Elements/Tissue | Targets | Effects | Implication in Diseases | References |
|---|---|---|---|---|
| Bronchial epithelium | TrkB P75NTR | Lung development Inhibit apoptosis and/or promote cell survival Airway remodeling NO release | ↑ BDNF in asthma ↑ BDNF and TrkB, ↓ p75NTR with infection ↑ Airway thickening in inflammation/asthma ↑ TrkB, p75NTR with cigarette smoke | [44,45,46,47] |
| Airway smooth muscle | TrkB P75NTR | Airway development and remodeling Increased airway contractility Cell proliferation | ↑ BDNF, TrkB with hyperoxia in fetal/neonatal ASM ↑ P75NTR and TrkB in inflammation/allergic diseases ↑ BDNF in airway inflammation ↑ ECM deposition ↑ TrkB, p75NTR with cigarette smoke ↑ ASM contractility with cigarette smoke and in inflammation | [1,2,3,41,48,49,50,51] |
| Nerves | TrkB P75NTR | NK1 receptor expression Neurotransmitter release Ach content of sensory nerves | ↑ BDNF in asthma ↑ Neuroplasticity ↑ Release of cholinergic neurotransmitters ↑ Neuromediated airway hyperresponsiveness | [3,44] |
| Fibroblasts | TrkB P75NTR | Proliferation Migration Differentiation | ↑ BDNF and TrkB in inflammation | [52,53,54,55,56] |
| Blood | ND | ND | ↑ BDNF in asthma ↑ p75NTR with cigarette smoke | [1,8,38,57] |
| BAL | ND | ND | ↑ BDNF in asthma | [58,59] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manti, S.; Xerra, F.; Spoto, G.; Butera, A.; Gitto, E.; Di Rosa, G.; Nicotera, A.G. Neurotrophins: Expression of Brain–Lung Axis Development. Int. J. Mol. Sci. 2023, 24, 7089. https://doi.org/10.3390/ijms24087089
Manti S, Xerra F, Spoto G, Butera A, Gitto E, Di Rosa G, Nicotera AG. Neurotrophins: Expression of Brain–Lung Axis Development. International Journal of Molecular Sciences. 2023; 24(8):7089. https://doi.org/10.3390/ijms24087089
Chicago/Turabian StyleManti, Sara, Federica Xerra, Giulia Spoto, Ambra Butera, Eloisa Gitto, Gabriella Di Rosa, and Antonio Gennaro Nicotera. 2023. "Neurotrophins: Expression of Brain–Lung Axis Development" International Journal of Molecular Sciences 24, no. 8: 7089. https://doi.org/10.3390/ijms24087089
APA StyleManti, S., Xerra, F., Spoto, G., Butera, A., Gitto, E., Di Rosa, G., & Nicotera, A. G. (2023). Neurotrophins: Expression of Brain–Lung Axis Development. International Journal of Molecular Sciences, 24(8), 7089. https://doi.org/10.3390/ijms24087089