
Can Neutrophils Prevent Nosocomial Pneumonia after Serious Injury?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. A Serious Risk of the Development of Pneumonia after Trauma
1.2. Mitochondrial DAMPs Released from Injured Tissues Predispose to Pneumonia Due to Reduced PMN Recruitment to the Lungs
1.3. Novel, Non-Antibiotic, Cell-Based Therapies May Prevent Pneumonia after Trauma
1.4. Granulocyte Transfusion
2. Our Vision: Application of PMN to Human Lungs Will Prevent/Cure Nosocomial Pneumonia after Injury
2.1. What Kind of PMN Can Be Applied Safely to the Lungs of Seriously Injured Trauma Patients to Prevent Nosocomial Pneumonia?
2.1.1. Any PMN Isolated from Human Donors Will Work?
2.1.2. Autologous PMN
2.1.3. PMN Modified to Be Negative for HLA
2.2. How to Store PMN before Applying Them to Trauma Patients
Cryopreservation of Human PMN
3. Supporting Data
3.1. Mouse Injury and Lung Infection Model
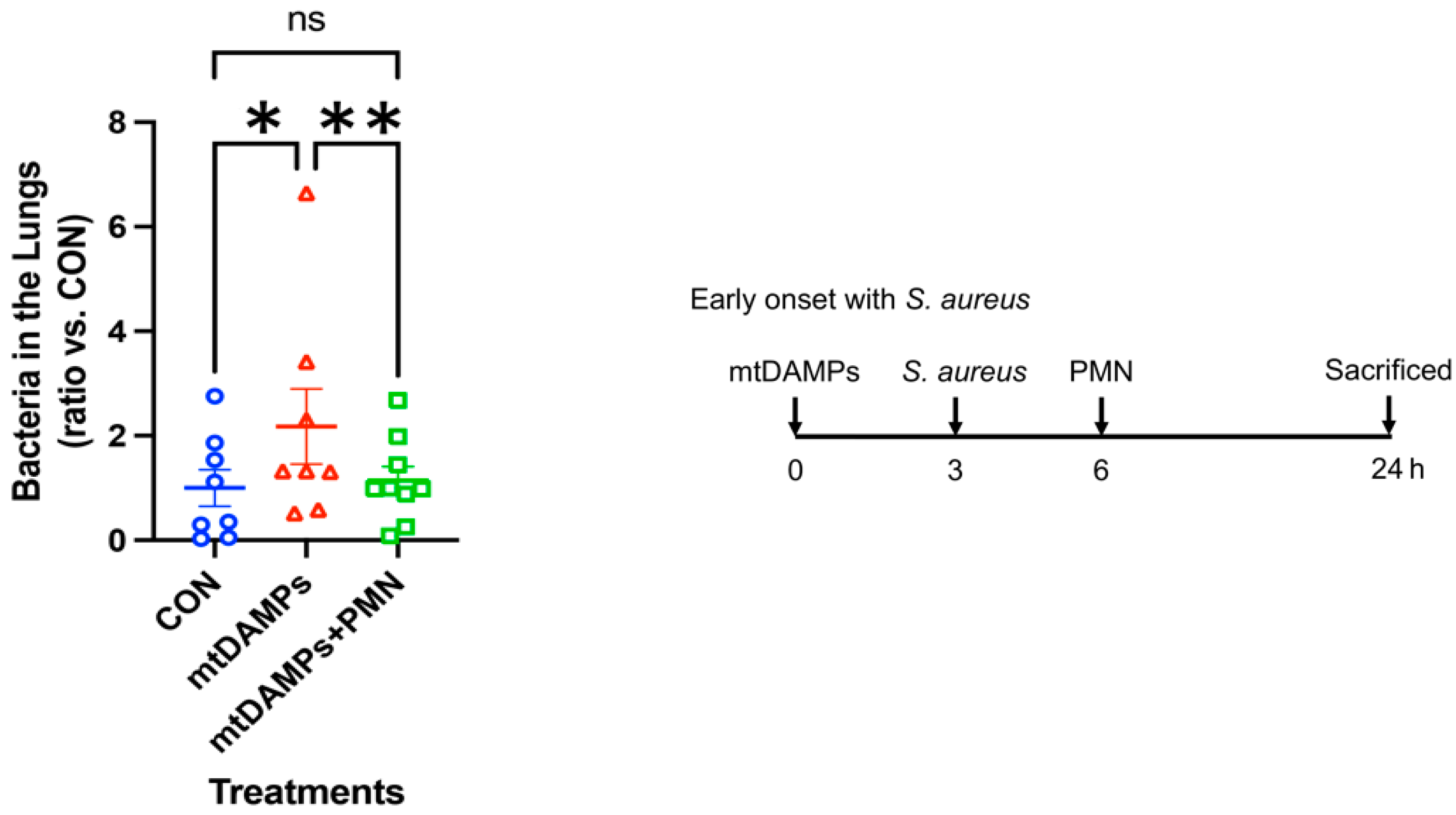
3.2. Prevention of Early Onset of Pneumonia
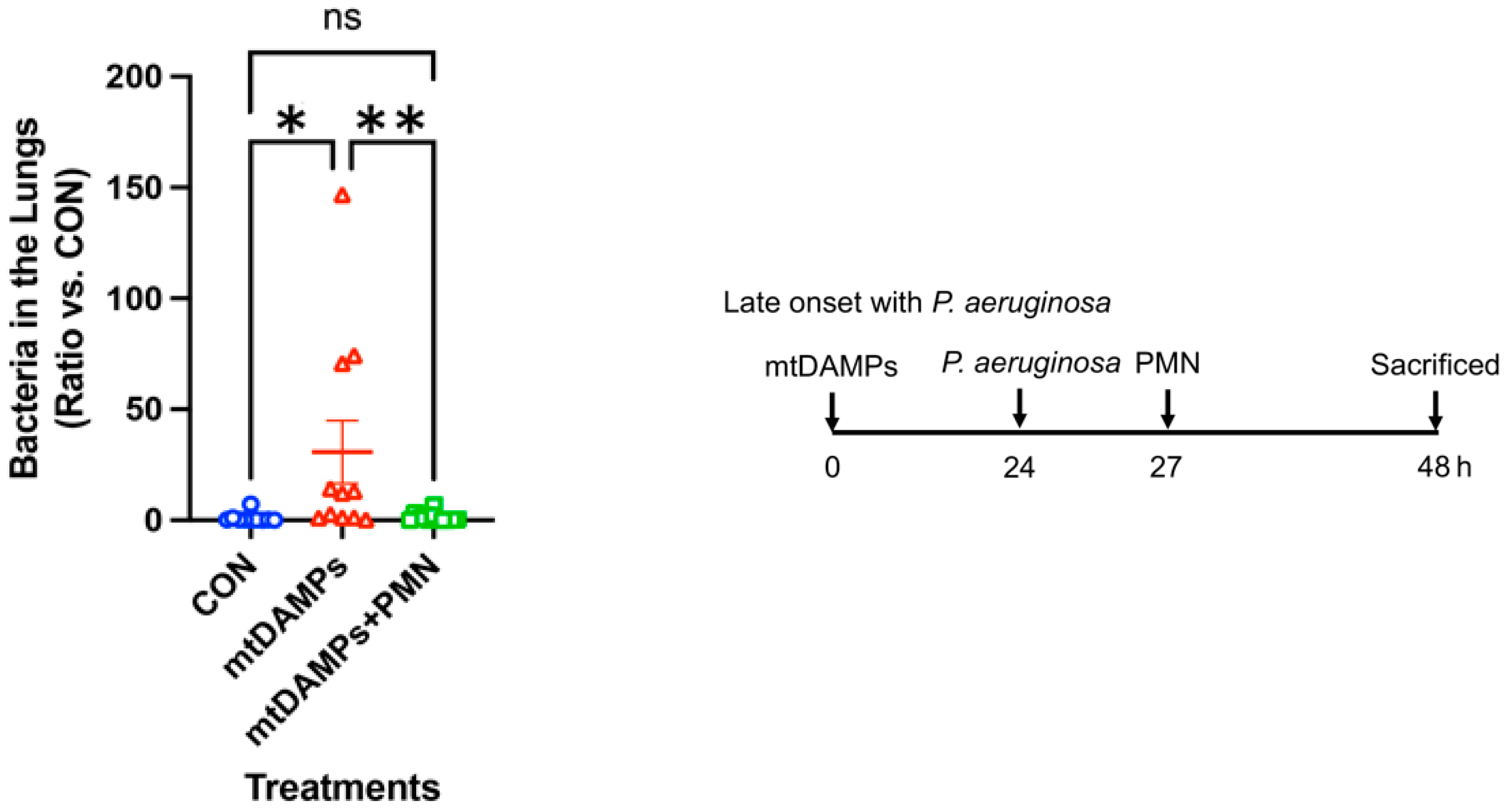
3.3. Prevention of Late-Onset Pneumonia
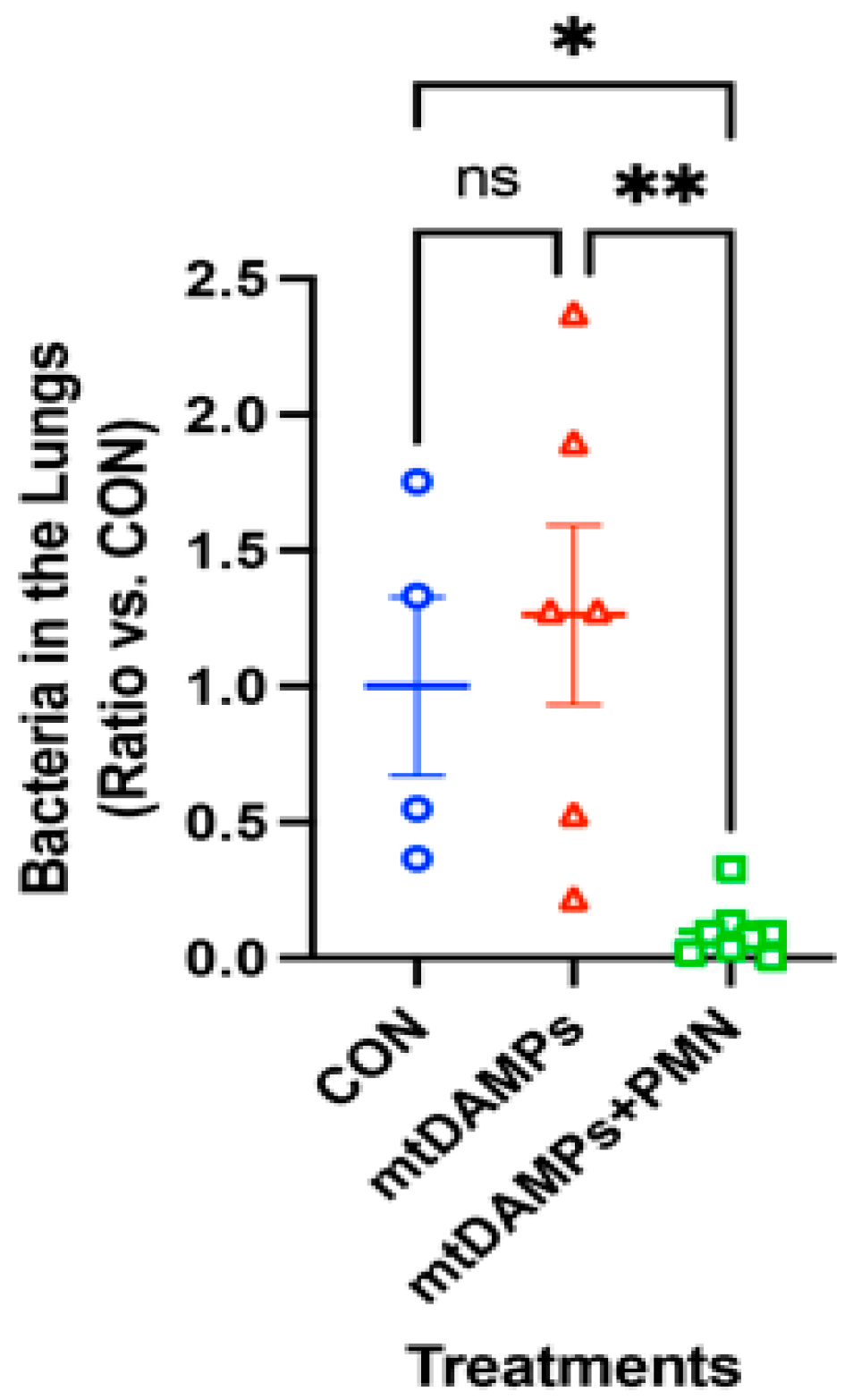
3.4. Prevention of Late Onset of Pneumonia with an Increased Number of P. aeruginosa
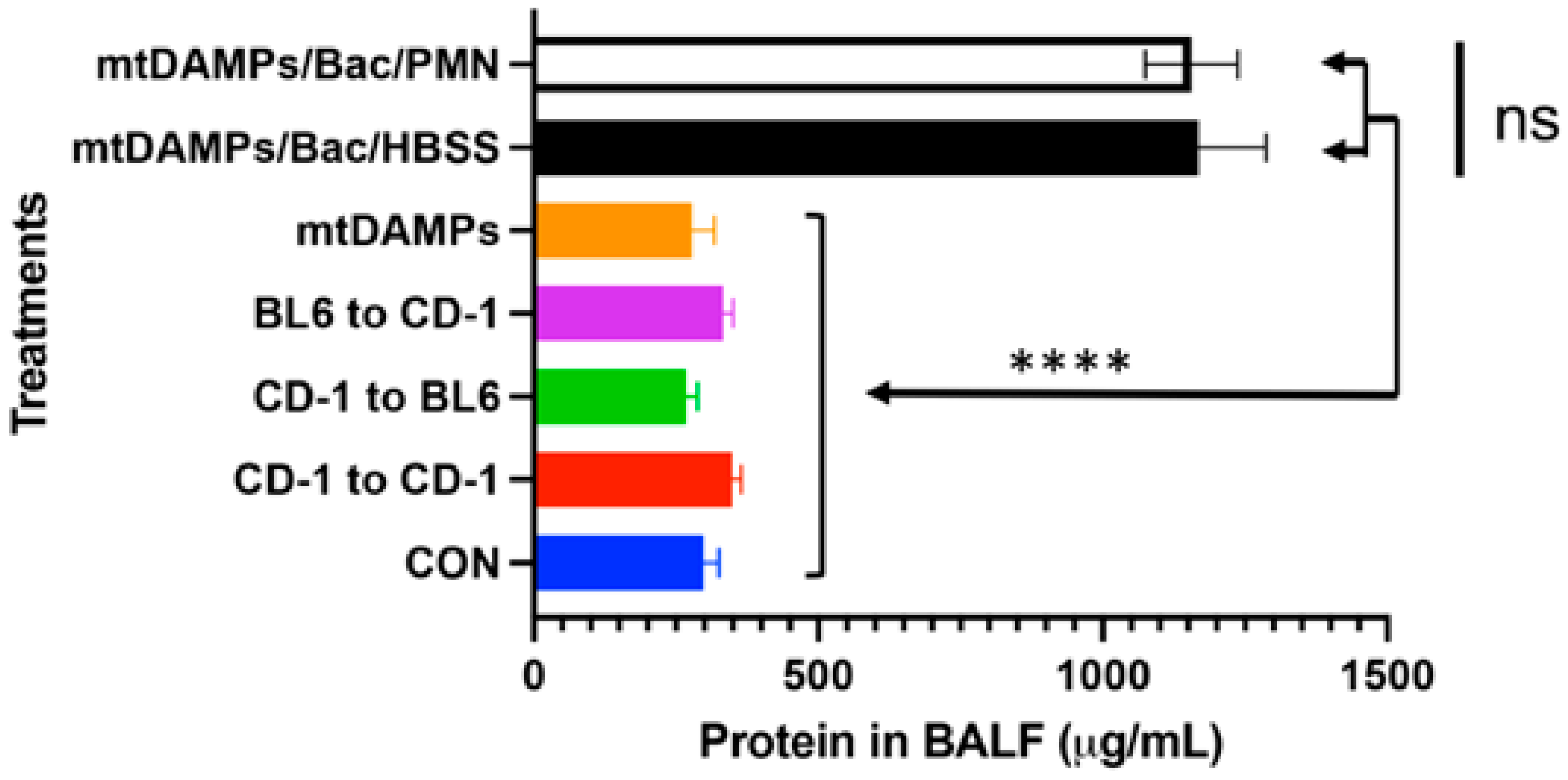
3.5. Evaluation of Potential Lung Injury Due to PMN Instillation
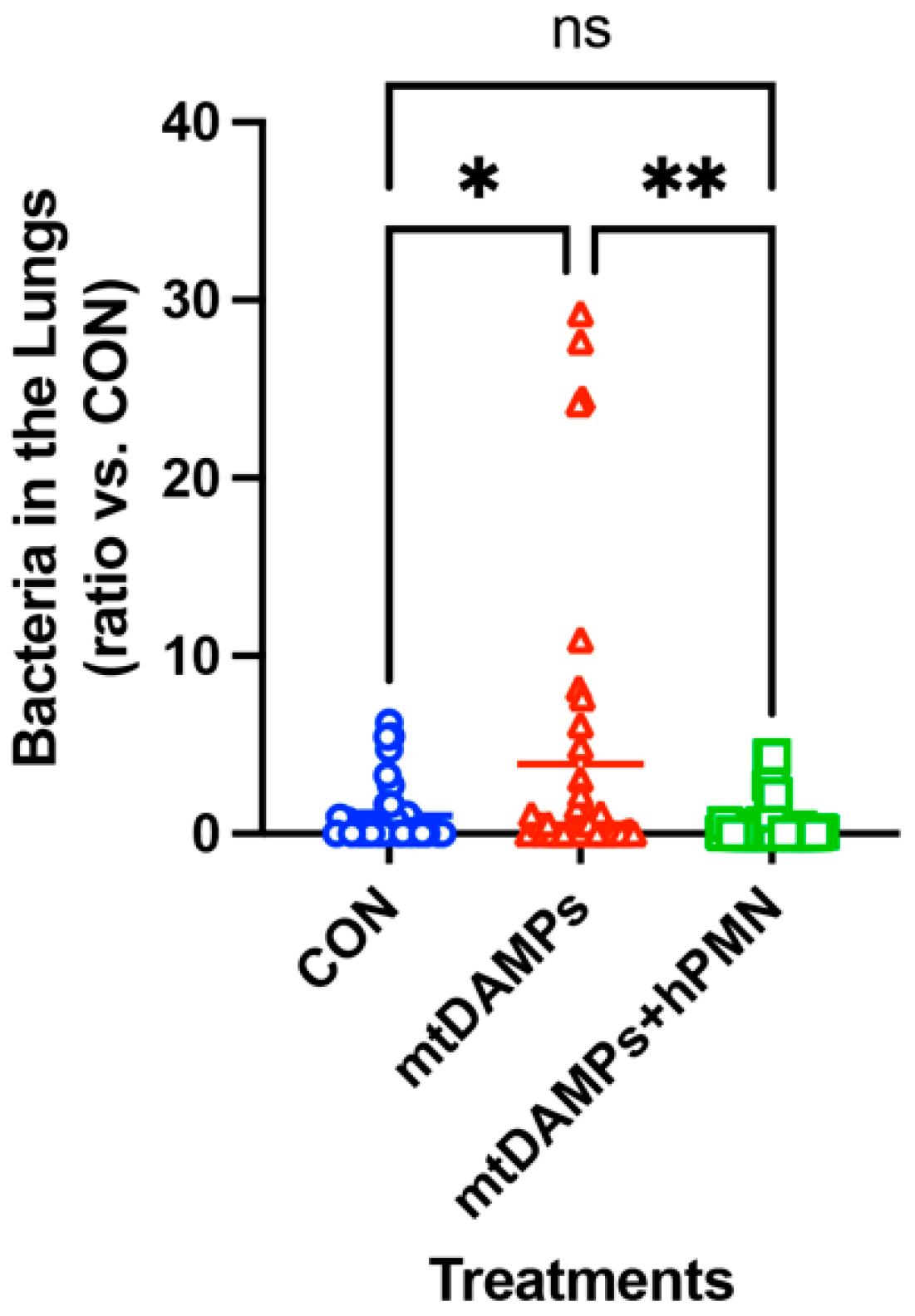
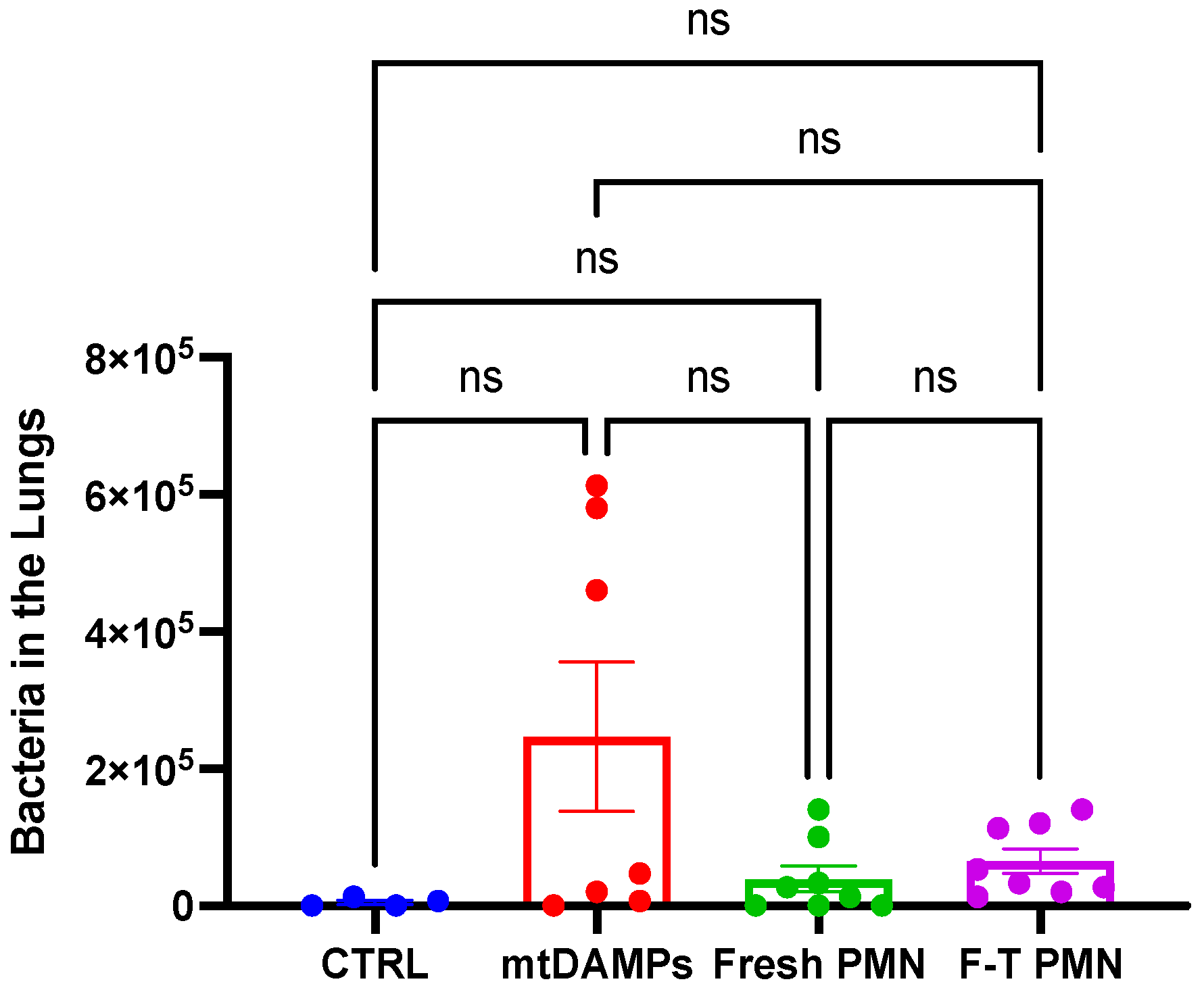
3.6. Human PMN Clears Bacteria in the Lungs without Causing Adverse Effects on the Recipients
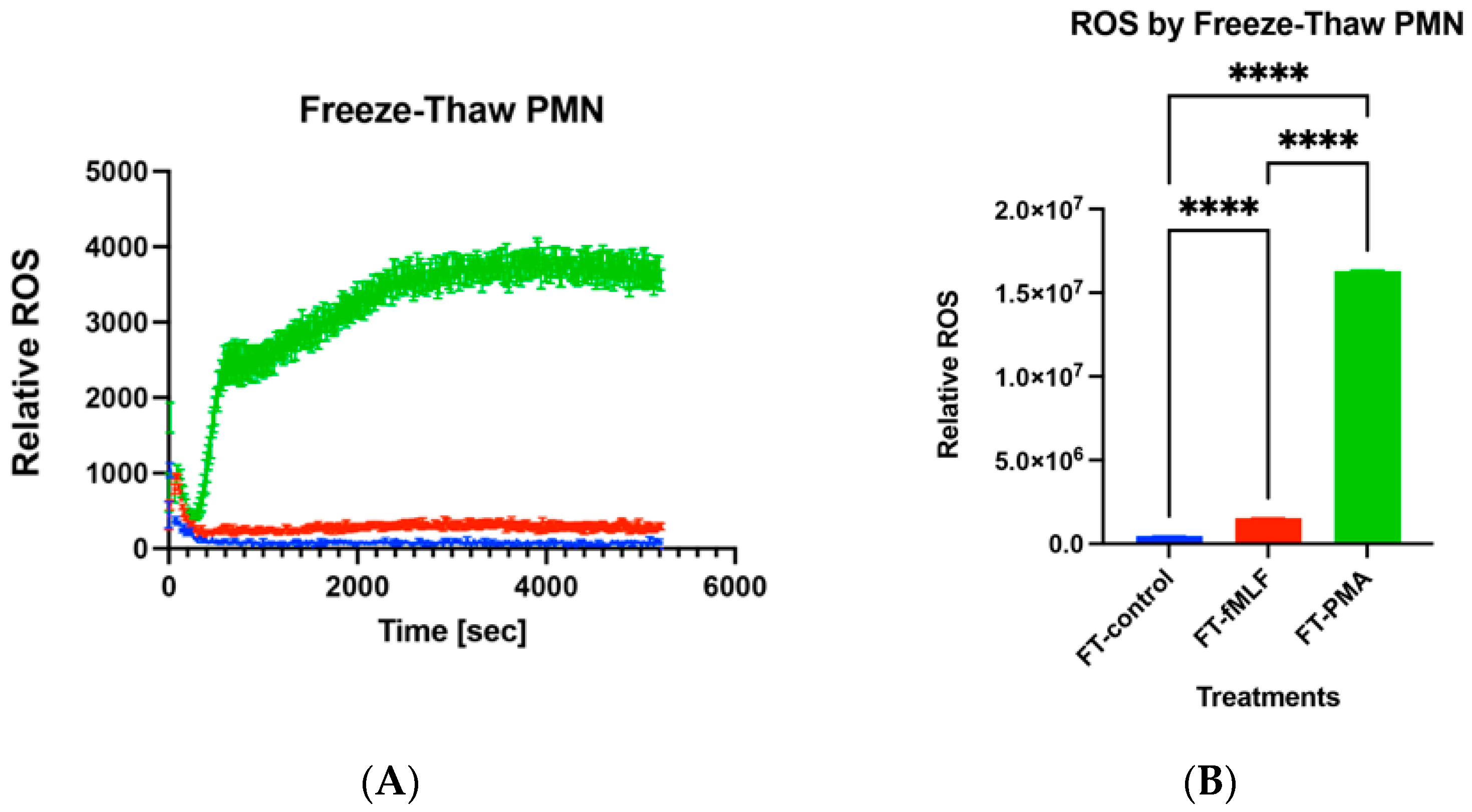
3.7. Functions of Freeze–Thaw PMN
4. Summary
5. More toward Human
6. Detailed Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chevret, S.; Hemmer, M.; Carlet, J.; Langer, M. Incidence and risk factors of pneumonia acquired in intensive care units. Results from a multicenter prospective study on 996 patients. European Cooperative Group on Nosocomial Pneumonia. Intensive Care Med. 1993, 19, 256–264. [Google Scholar]
- Papia, G.; Louie, M.; Tralla, A.; Johnson, C.; Collins, V.; Simor, A.E. Screening high-risk patients for methicillin-resistant Staphylococcus aureus on admission to the hospital: Is it cost effective? Infect. Control Hosp. Epidemiol. 1999, 20, 473–477. [Google Scholar] [CrossRef]
- Papia, G.; McLellan, B.A.; El-Helou, P.; Louie, M.; Rachlis, A.; Szalai, J.P.; Simor, A.E. Infection in hospitalized trauma patients: Incidence, risk factors, and complications. J. Trauma 1999, 47, 923–927. [Google Scholar] [CrossRef]
- Wallace, W.C.; Cinat, M.; Gornick, W.B.; Lekawa, M.E.; Wilson, S.E. Nosocomial infections in the surgical intensive care unit: A difference between trauma and surgical patients. Am. Surg. 1999, 65, 987–990. [Google Scholar] [CrossRef]
- Kozka, M.; Sega, A.; Wojnar-Gruszka, K.; Tarnawska, A.; Gniadek, A. Risk Factors of Pneumonia Associated with Mechanical Ventilation. Int. J. Environ. Res. Public Health 2020, 17, 656. [Google Scholar] [CrossRef]
- Margulis, L. Genetic and evolutionary consequences of symbiosis. Exp. Parasitol. 1976, 39, 277–349. [Google Scholar]
- Itagaki, K.; Kaczmarek, E.; Kwon, W.Y.; Chen, L.; Vlkova, B.; Zhang, Q.; Rica, I.; Yaffe, M.B.; Campbell, Y.; Marusich, M.F.; et al. Formyl Peptide Receptor-1 Blockade Prevents Receptor Regulation by Mitochondrial Danger-Associated Molecular Patterns and Preserves Neutrophil Function After Trauma. Crit. Care Med. 2020, 48, e123–e132. [Google Scholar]
- Raoof, M.; Zhang, Q.; Itagaki, K.; Hauser, C.J. Mitochondrial peptides are potent immune activators that activate human neutrophils via FPR-1. J. Trauma 2010, 68, 1328–1332; discussion 1332–1334. [Google Scholar] [CrossRef]
- Sun, S.; Sursal, T.; Adibnia, Y.; Zhao, C.; Zheng, Y.; Li, H.; Otterbein, L.E.; Hauser, C.J.; Itagaki, K. Mitochondrial DAMPs increase endothelial permeability through neutrophil dependent and independent pathways. PLoS ONE 2013, 8, e59989. [Google Scholar] [CrossRef]
- Sursal, T.; Stearns-Kurosawa, D.J.; Itagaki, K.; Oh, S.-Y.; Sun, S.; Kurosawa, S.; Hauser, C.J. Plasma bacterial and mitochondrial DNA distinguish bacterial sepsis from sterile systemic inflammatory response syndrome and quantify inflammatory tissue injury in nonhuman primates. Shock 2013, 39, 55–62. [Google Scholar] [CrossRef]
- Zhang, Q.; Raoof, M.; Chen, Y.; Sumi, Y.; Sursal, T.; Junger, W.; Brohi, K.; Itagaki, K.; Hauser, C.J. Circulating mitochondrial DAMPs cause inflammatory responses to injury. Nature 2010, 464, 104–107. [Google Scholar] [CrossRef]
- Zhao, C.; Itagaki, K.; Gupta, A.; Odom, S.; Sandler, N.; Hauser, C.J. Mitochondrial damage-associated molecular patterns released by abdominal trauma suppress pulmonary immune responses. J. Trauma Acute Care Surg. 2014, 76, 1222–1227. [Google Scholar] [CrossRef]
- Kaczmarek, E.; Hauser, C.J.; Kwon, W.Y.; Rica, I.; Chen, L.; Sandler, N.; Otterbein, L.E.; Campbell, Y.; Cook, C.H.; Yaffe, M.B.; et al. A subset of five human mitochondrial formyl peptides mimics bacterial peptides and functionally deactivates human neutrophils. J. Trauma Acute Care Surg. 2018, 85, 936–943. [Google Scholar] [CrossRef] [PubMed]
- Itagaki, K.; Rica, I.; Zhang, J.; Gallo, D.; DePrato, M.; Otterbein, L.E.; Hauser, C.J. Intratracheal instillation of neutrophils rescues bacterial overgrowth initiated by trauma damage-associated molecular patterns. J. Trauma Acute Care Surg. 2017, 82, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Tarlowe, M.H.; Duffy, A.; Kannan, K.B.; Itagaki, K.; Lavery, R.F.; Livingston, D.H.; Bankey, P.; Hauser, C.J. Prospective study of neutrophil chemokine responses in trauma patients at risk for pneumonia. Am. J. Respir. Crit. Care Med. 2005, 171, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Tarlowe, M.H.; Kannan, K.B.; Itagaki, K.; Adams, J.M.; Livingston, D.H.; Hauser, C.J. Inflammatory chemoreceptor cross-talk suppresses leukotriene B4 receptor 1-mediated neutrophil calcium mobilization and chemotaxis after trauma. J. Immunol. 2003, 171, 2066–2073. [Google Scholar] [CrossRef] [PubMed]
- Bronchard, R.; Albaladejo, P.; Brezac, G.; Geffroy, A.; Seince, P.F.; Morris, W.; Branger, C.; Marty, J. Early onset pneumonia: Risk factors and consequences in head trauma patients. Anesthesiology 2004, 100, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Maze, M.; Koch, L.G.; Britton, S.L.; Hellman, J. Exaggerated Acute Lung Injury and Impaired Antibacterial Defenses During Staphylococcus aureus Infection in Rats with the Metabolic Syndrome. PLoS ONE 2015, 10, e0126906. [Google Scholar] [CrossRef] [PubMed]
- Launey, Y.; Asehnoune, K.; Lasocki, S.; Dahyot-Fizelier, C.; Huet, O.; Le Pabic, E.; Malejac, B.; Seguin, P.; AtlanRea, G. Risk factors for ventilator-associated pneumonia due to Staphylococcus aureus in patients with severe brain injury: A multicentre retrospective cohort study. Anaesth. Crit. Care Pain Med. 2021, 40, 100785. [Google Scholar] [CrossRef]
- Rello, J.; Ausina, V.; Castella, J.; Net, A.; Prats, G. Nosocomial respiratory tract infections in multiple trauma patients. Influence of level of consciousness with implications for therapy. Chest 1992, 102, 525–529. [Google Scholar] [CrossRef]
- Kelly, E.; Bailey, C.P.; Henderson, G. Agonist-selective mechanisms of GPCR desensitization. Br. J. Pharmacol. 2008, 153 (Suppl. S1), S379–S388. [Google Scholar] [CrossRef]
- Chambers, H.F.; Deleo, F.R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef]
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef]
- Gea-Banacloche, J. Granulocyte transfusions: A concise review for practitioners. Cytotherapy 2017, 19, 1256–1269. [Google Scholar] [CrossRef] [PubMed]
- Hazeldine, J.; Hampson, P.; Lord, J.M. The impact of trauma on neutrophil function. Injury 2014, 45, 1824–1833. [Google Scholar] [CrossRef]
- Estcourt, L.J.; Stanworth, S.J.; Hopewell, S.; Doree, C.; Trivella, M.; Massey, E. Granulocyte transfusions for treating infections in people with neutropenia or neutrophil dysfunction. Cochrane Database Syst. Rev. 2016, 4, CD005339. [Google Scholar] [CrossRef]
- Malkin, M.; Nevo, A.; Brundage, S.I.; Schreiber, M. Effectiveness and safety of whole blood compared to balanced blood components in resuscitation of hemorrhaging trauma patients-A systematic review. Injury 2021, 52, 182–188. [Google Scholar] [CrossRef]
- Xiao, W.; Mindrinos, M.N.; Seok, J.; Cuschieri, J.; Cuenca, A.G.; Gao, H.; Hayden, D.L.; Hennessy, L.; Moore, E.E.; Minei, J.P.; et al. A genomic storm in critically injured humans. J. Exp. Med. 2011, 208, 2581–2590. [Google Scholar] [CrossRef]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef]
- Abraham, E. Neutrophils and acute lung injury. Crit. Care Med. 2003, 31 (Suppl. S4), S195–S199. [Google Scholar] [CrossRef]
- Paladino, L.; Subramanian, R.A.; Bonilla, E.; Sinert, R.H. Leukocytosis as prognostic indicator of major injury. West. J. Emerg. Med. 2010, 11, 450–455. [Google Scholar] [PubMed]
- Junger, W.G.; Rhind, S.G.; Rizoli, S.B.; Cuschieri, J.; Baker, A.J.; Shek, P.N.; Hoyt, D.B.; Bulger, E.M. Prehospital hypertonic saline resuscitation attenuates the activation and promotes apoptosis of neutrophils in patients with severe traumatic brain injury. Shock 2013, 40, 366–374. [Google Scholar] [PubMed]
- Junger, W.G.; Rhind, S.G.; Rizoli, S.B.; Cuschieri, J.; Shiu, M.Y.; Baker, A.J.; Li, L.; Shek, P.N.; Hoyt, D.B.; Bulger, E.M. Resuscitation of traumatic hemorrhagic shock patients with hypertonic saline-without dextran-inhibits neutrophil and endothelial cell activation. Shock 2012, 38, 341–350. [Google Scholar] [PubMed]
- Kanyilmaz, S.; Hepguler, S.; Atamaz, F.C.; Gokmen, N.M.; Ardeniz, O.; Sin, A. Phagocytic and oxidative burst activity of neutrophils in patients with spinal cord injury. Arch. Phys. Med. Rehabil. 2013, 94, 369–374. [Google Scholar] [CrossRef]
- Kinoshita, M.; Miyazaki, H.; Ono, S.; Inatsu, A.; Nakashima, H.; Tsujimoto, H.; Shinomiya, N.; Saitoh, D.; Seki, S. Enhancement of neutrophil function by interleukin-18 therapy protects burn-injured mice from methicillin-resistant Staphylococcus aureus. Infect. Immun. 2011, 79, 2670–2680. [Google Scholar] [CrossRef]
- Kurihara, T.; Jones, C.N.; Yu, Y.M.; Fischman, A.J.; Watada, S.; Tompkins, R.G.; Fagan, S.P.; Irimia, D. Resolvin D2 restores neutrophil directionality and improves survival after burns. FASEB J. 2013, 27, 2270–2281. [Google Scholar]
- Liao, Y.; Liu, P.; Guo, F.; Zhang, Z.Y.; Zhang, Z. Oxidative burst of circulating neutrophils following traumatic brain injury in human. PLoS ONE 2013, 8, e68963. [Google Scholar] [CrossRef]
- Xu, H.; Wang, B.; Ono, M.; Kagita, A.; Fujii, K.; Sasakawa, N.; Ueda, T.; Gee, P.; Nishikawa, M.; Nomura, M.; et al. Targeted Disruption of HLA Genes via CRISPR-Cas9 Generates iPSCs with Enhanced Immune Compatibility. Cell Stem Cell 2019, 24, 566–578.e7. [Google Scholar] [CrossRef]
- Takahashi, T.; Inada, S.; Pommier, C.G.; O’Shea, J.J.; Brown, E.J. Osmotic stress and the freeze-thaw cycle cause shedding of Fc and C3b receptors by human polymorphonuclear leukocytes. J. Immunol. 1985, 134, 4062–4068. [Google Scholar] [CrossRef]
- Lima, S.S.; Franca, M.S.; Godoi, C.C.; Martinho, G.H.; de Jesus, L.A.; Romanelli, R.M.; Clemente, W.T. Neutropenic patients and their infectious complications at a University Hospital. Rev. Bras. Hematol. Hemoter. 2013, 35, 18–22. [Google Scholar] [CrossRef]
- Lustberg, M.B. Management of neutropenia in cancer patients. Clin. Adv. Hematol. Oncol. 2012, 10, 825–826. [Google Scholar]
- Boxer, L.A. Severe congenital neutropenia: Genetics and pathogenesis. Trans. Am. Clin. Climatol. Assoc. 2006, 117, 13–31; discussion 31-2. [Google Scholar] [PubMed]
- Sapey, E.; Greenwood, H.; Walton, G.; Mann, E.; Love, A.; Aaronson, N.; Insall, R.H.; Stockley, R.A.; Lord, J.M. Phosphoinositide 3-kinase inhibition restores neutrophil accuracy in the elderly: Toward targeted treatments for immunosenescence. Blood 2014, 123, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Kwon, W.Y.; Vlkova, B.; Riça, I.; Kaczmarek, E.; Park, J.; Kim, H.I.; Konecna, B.; Jung, F.; Douglas, G.; et al. Direct airway instillation of neutrophils overcomes chemotactic deficits induced by injury. Shock 2021, 56, 119–124. [Google Scholar] [CrossRef]
- Kim, H.I.; Park, J.; Konecna, B.; Huang, W.; Rica, I.; Gallo, D.; Otterbein, L.E.; Itagaki, K.; Hauser, C.J. Plasma and wound fluids from trauma patients suppress neutrophil extracellular respiratory burst. J. Trauma Acute Care Surg. 2022, 92, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Meurens, F.; Summerfield, A.; Nauwynck, H.; Saif, L.; Gerdts, V. The pig: A model for human infectious diseases. Trends Microbiol. 2012, 20, 50–57. [Google Scholar] [CrossRef]
- Judge, E.P.; Hughes, J.M.; Egan, J.J.; Maguire, M.; Molloy, E.L.; O’Dea, S. Anatomy and bronchoscopy of the porcine lung. A model for translational respiratory medicine. Am. J. Respir. Cell Mol. Biol. 2014, 51, 334–343. [Google Scholar] [CrossRef]
- Wang, J.; Hossain, M.; Thanabalasuriar, A.; Gunzer, M.; Meininger, C.; Kubes, P. Visualizing the function and fate of neutrophils in sterile injury and repair. Science 2017, 358, 111–116. [Google Scholar] [CrossRef]
- Itagaki, K.; Kannan, K.B.; Livingston, D.H.; Deitch, E.A.; Fekete, Z.; Hauser, C.J. Store-operated calcium entry in human neutrophils reflects multiple contributions from independently regulated pathways. J. Immunol. 2002, 168, 4063–4069. [Google Scholar] [CrossRef]
- Swamydas, M.; Lionakis, M.S. Isolation, purification and labeling of mouse bone marrow neutrophils for functional studies and adoptive transfer experiments. J. Vis. Exp. 2013, 77, e50586. [Google Scholar]
- Itagaki, K.; Rica, I.; Konecna, B.; Kim, H.I.; Park, J.; Kaczmarek, E.; Hauser, C.J. Role of Mitochondria-Derived Danger Signals Released after Injury in Systemic Inflammation and Sepsis. Antioxid. Redox Signal. 2021, 35, 1273–1290. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macáková, K.; Kaczmarek, E.; Itagaki, K. Can Neutrophils Prevent Nosocomial Pneumonia after Serious Injury? Int. J. Mol. Sci. 2023, 24, 7627. https://doi.org/10.3390/ijms24087627
Macáková K, Kaczmarek E, Itagaki K. Can Neutrophils Prevent Nosocomial Pneumonia after Serious Injury? International Journal of Molecular Sciences. 2023; 24(8):7627. https://doi.org/10.3390/ijms24087627
Chicago/Turabian StyleMacáková, Kristína, Elzbieta Kaczmarek, and Kiyoshi Itagaki. 2023. "Can Neutrophils Prevent Nosocomial Pneumonia after Serious Injury?" International Journal of Molecular Sciences 24, no. 8: 7627. https://doi.org/10.3390/ijms24087627
APA StyleMacáková, K., Kaczmarek, E., & Itagaki, K. (2023). Can Neutrophils Prevent Nosocomial Pneumonia after Serious Injury? International Journal of Molecular Sciences, 24(8), 7627. https://doi.org/10.3390/ijms24087627