
Chicken Interferon-Alpha and -Lambda Exhibit Antiviral Effects against Fowl Adenovirus Serotype 4 in Leghorn Male Hepatocellular Cells

Abstract
:1. Introduction
2. Results
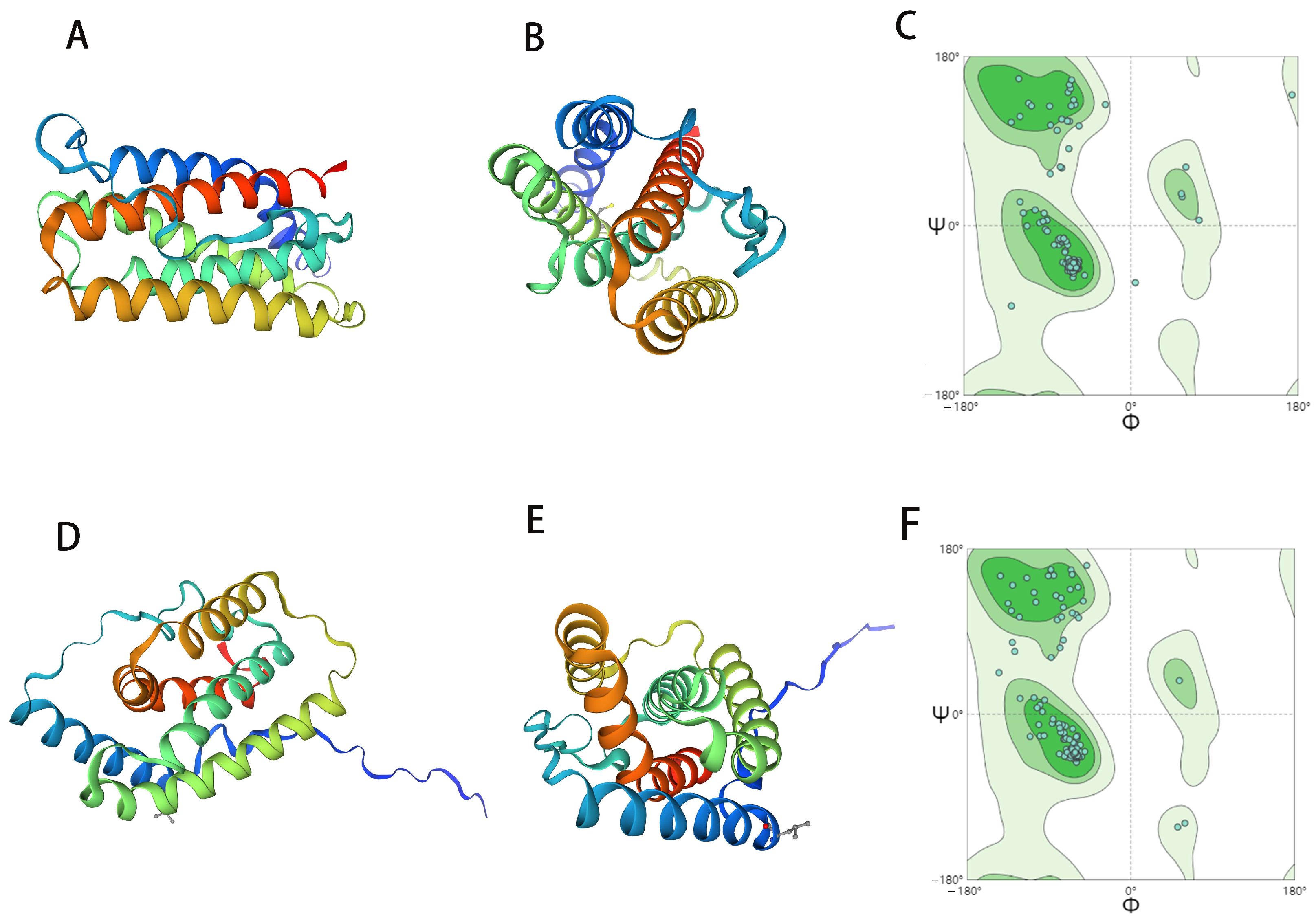
2.1. ChIFN-α and ChIFN-λ Structure Prediction
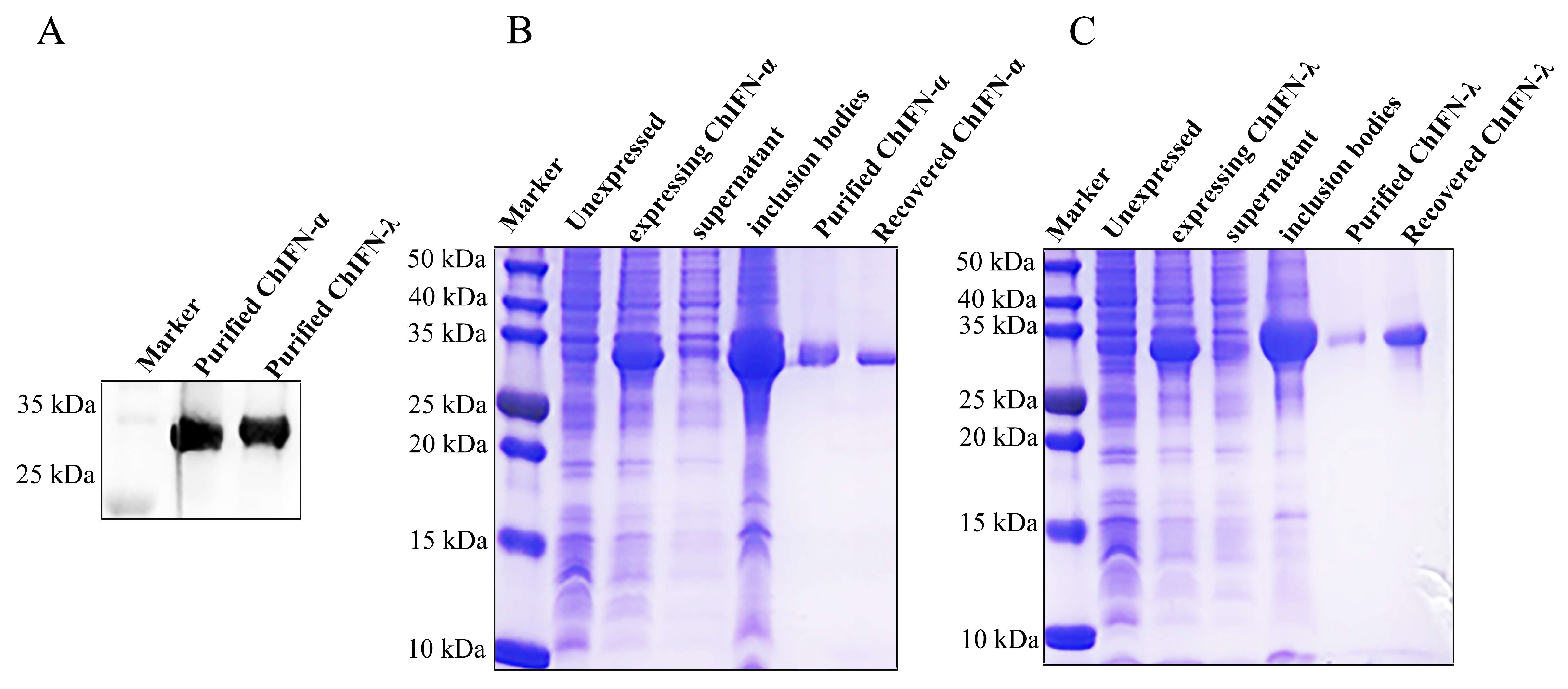
2.2. ChIFN-α and ChIFN-λ Expression and Purification
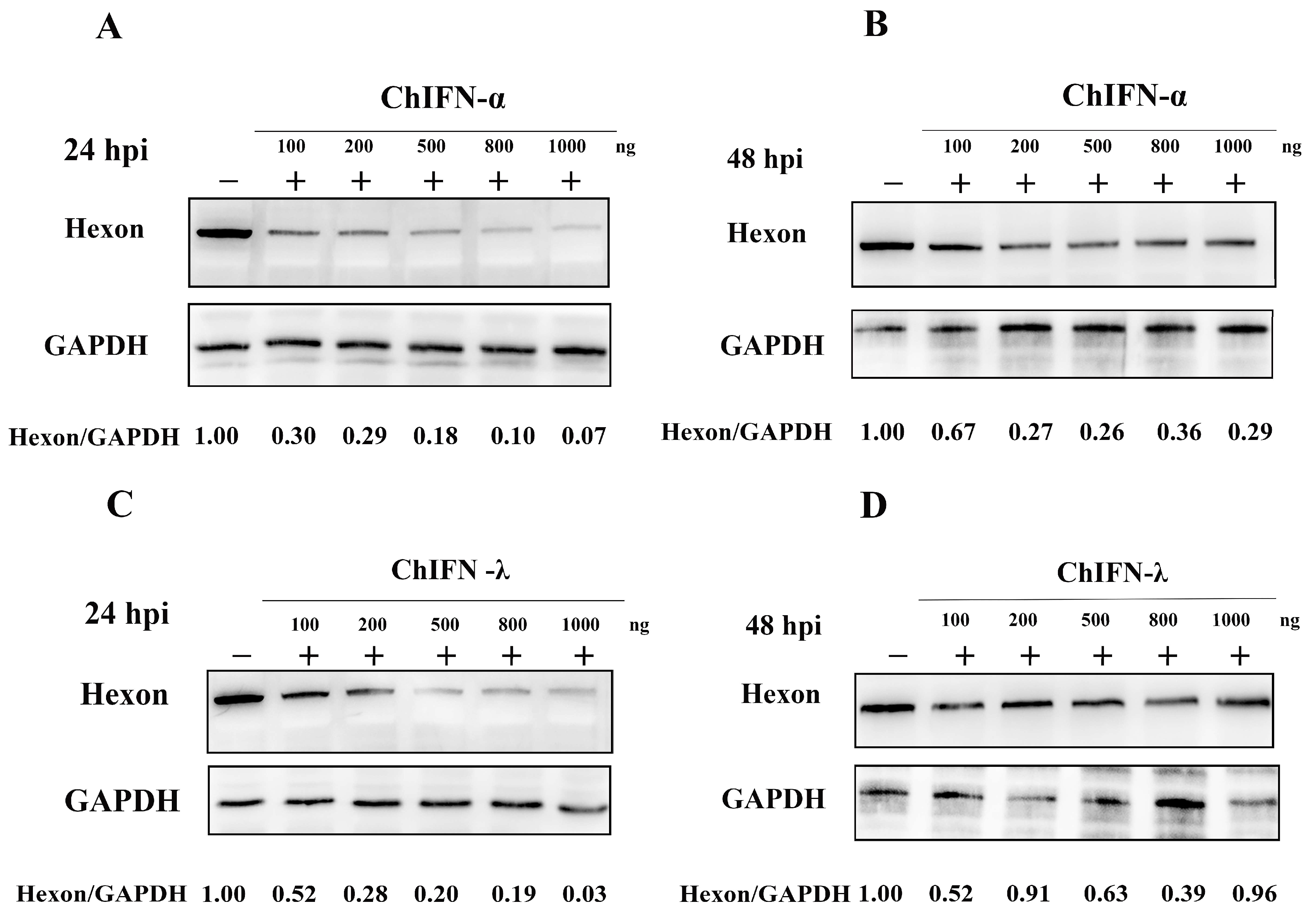
2.3. Assessing the Antiviral Efficacy of ChIFN-α and ChIFN-λ
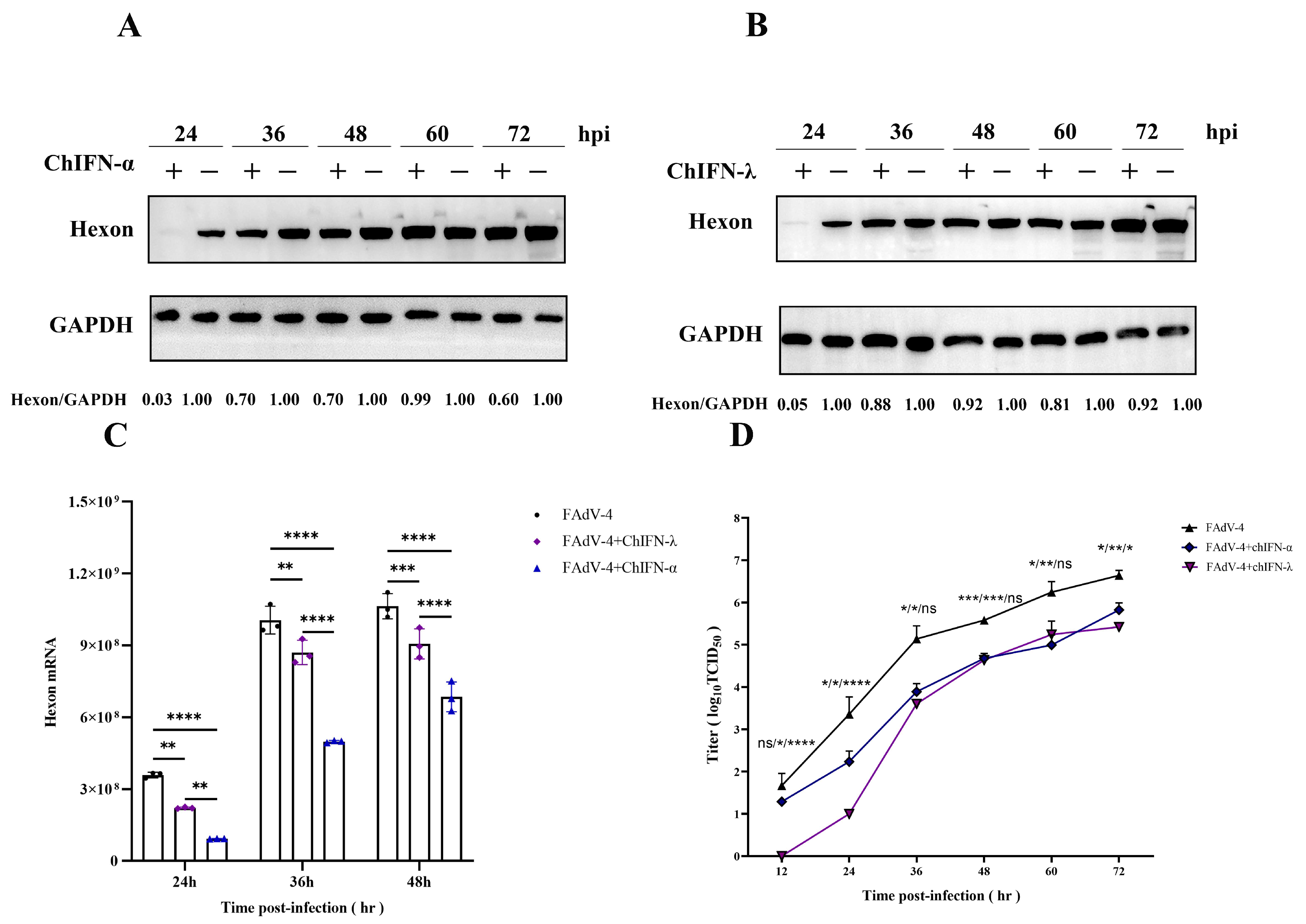
2.4. ChIFN-α and ChIFN-λ Pretreatments Restrict FAdV-4 within a Specific Temporal Horizon
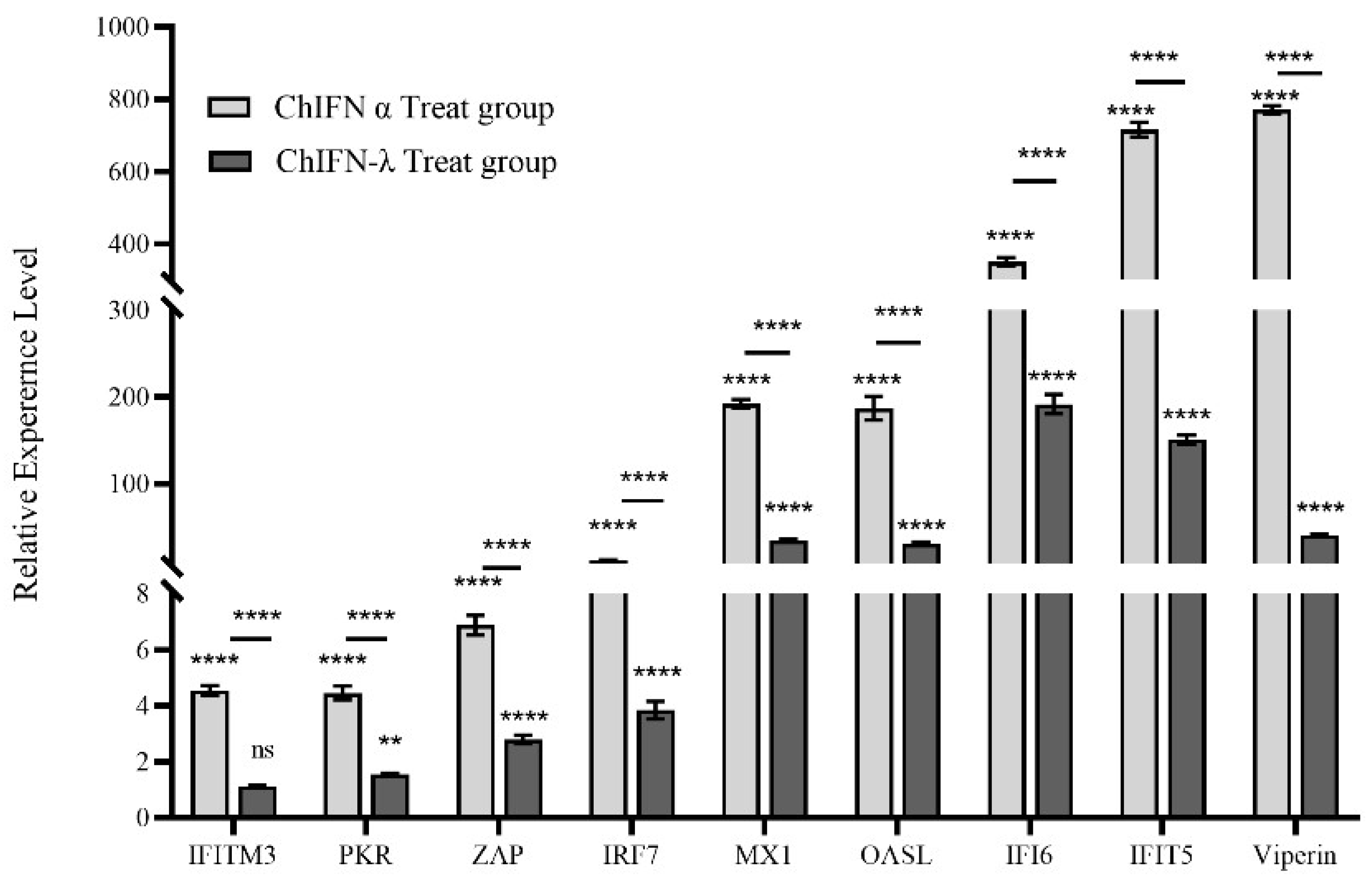
2.5. ChIFN-α and ChIFN-λ Induce ISG Expression in LMH Cells
3. Discussion
4. Materials and Methods
4.1. Cells, Viruses, and Antibodies
4.2. ChIFN-α and ChIFN-λ Structure Prediction and Epression
4.3. IFN Purification and Renaturation
4.4. Antiviral Assay
4.5. Western Blot Analysis
4.6. Virus Titration
4.7. Quantitative Real-Time PCR
4.8. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xia, J.; Yao, K.C.; Liu, Y.Y.; You, G.J.; Li, S.Y.; Liu, P.; Zhao, Q.; Wen Rui Wu, Y.P.; Huang, X.B.; Cao, S.J.; et al. Isolation and molecular characterization of prevalent Fowl adenovirus strains in southwestern China during 2015–2016 for the development of a control strategy. Emerg. Microbes Infect. 2017, 6, e103. [Google Scholar] [CrossRef]
- Ren, G.; Wang, H.; Yan, Y.; Liu, F.; Huang, M.; Chen, R. Pathogenicity of a fowl adenovirus serotype 4 isolated from chickens associated with hydropericardium-hepatitis syndrome in China. Poult. Sci. 2019, 98, 2765–2771. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, J.; Qiu, L.; Han, Z.; Liu, S. Fowl adenovirus species C serotype 4 is attributed to the emergence of hepatitis-hydropericardium syndrome in chickens in China. Infect. Genet. Evol. 2016, 45, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.H.; Sun, L.; Tu, K.H.; Teng, Q.Y.; Xue, J.; Zhang, G.Z. Experimental co-infection of variant infectious bursal disease virus and fowl adenovirus serotype 4 increases mortality and reduces immune response in chickens. Vet. Res. 2021, 52, 61. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Sun, Q.; Shi, Y.; Ding, Y.; Li, Z.; Sun, Y.; Li, M.; Liu, S. Immunosuppressive potential of fowl adenovirus serotype 4. Poult. Sci. 2019, 98, 3514–3522. [Google Scholar] [CrossRef] [PubMed]
- Ganesh, K.; Raghavan, R. Hydropericardium hepatitis syndrome of broiler poultry: Current status of research. Res. Vet. Sci. 2000, 68, 201–206. [Google Scholar] [CrossRef]
- Shen, Z.; Xiang, B.; Li, S.; Ren, X.; Hong, Y.; Liao, J.; Yu, D.; Ren, T.; Liao, M.; Xu, C. Genetic characterization of fowl adenovirus serotype 4 isolates in Southern China reveals potential cross-species transmission. Infect. Genet. Evol. 2019, 75, 103928. [Google Scholar] [CrossRef]
- Jiang, Z.; Liu, M.; Wang, C.; Zhou, X.; Li, F.; Song, J.; Pu, J.; Sun, Y.; Wang, M.; Shahid, M.; et al. Characterization of fowl adenovirus serotype 4 circulating in chickens in China. Vet. Microbiol. 2019, 238, 108427. [Google Scholar] [CrossRef] [PubMed]
- Niczyporuk, J.S.; Kozdrun, W.; Czekaj, H.; Piekarska, K.; Stys-Fijol, N. Isolation and molecular characterization of Fowl adenovirus strains in Black grouse: First reported case in Poland. PLoS ONE 2020, 15, e0234532. [Google Scholar] [CrossRef]
- Li, P.H.; Zheng, P.P.; Zhang, T.F.; Wen, G.Y.; Shao, H.B.; Luo, Q.P. Fowl adenovirus serotype 4: Epidemiology, pathogenesis, diagnostic detection, and vaccine strategies. Poult. Sci. 2017, 96, 2630–2640. [Google Scholar] [CrossRef]
- Meng, F.; Dong, G.; Zhang, Y.; Tian, S.; Cui, Z.; Chang, S.; Zhao, P. Co-infection of fowl adenovirus with different immunosuppressive viruses in a chicken flock. Poult. Sci. 2018, 97, 1699–1705. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.; Yang, L.; Chen, F.; He, X.; Zhang, R.; Zhao, Y.; Gao, G.; Mu, W.; Chen, X.; Luo, S.; et al. Prevalence and Molecular Characteristics of FAdV-4 from Indigenous Chicken Breeds in Yunnan Province, Southwestern China. Microorganisms 2023, 11, 2631. [Google Scholar] [CrossRef] [PubMed]
- Pestka, S.; Krause, C.D.; Walter, M.R. Interferons, interferon-like cytokines, and their receptors. Immunol. Rev. 2004, 202, 8–32. [Google Scholar] [CrossRef]
- Sick, C.; Schultz, U.; Staeheli, P. A family of genes coding for two serologically distinct chicken interferons. J. Biol. Chem. 1996, 271, 7635–7639. [Google Scholar] [CrossRef]
- Karpala, A.J.; Morris, K.R.; Broadway, M.M.; McWaters, P.G.; O’Neil, T.E.; Goossens, K.E.; Lowenthal, J.W.; Bean, A.G. Molecular cloning, expression, and characterization of chicken IFN -lambda. J. Interferon Cytokine Res. 2008, 28, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Sekellick, M.J.; Ferrandino, A.F.; Hopkins, D.A.; Marcus, P.I. Chicken interferon gene: Cloning, expression, and analysis. J. Interferon Res. 1994, 14, 71–79. [Google Scholar] [CrossRef]
- Zhao, W.; Li, X.; Li, H.; Han, Z.; Wang, F.; Liu, C.; Shao, Y.; Ma, D. Fowl adenoviruse-4 infection induces strong innate immune responses in chicken. Comp. Immunol. Microbiol. Infect. Dis. 2020, 68, 101404. [Google Scholar] [CrossRef]
- Wu, N.; Yang, B.; Wen, B.; Li, W.; Guo, J.; Qi, X.; Wang, J. Pathogenicity and Immune Responses in Specific-Pathogen-Free Chickens During Fowl Adenovirus Serotype 4 Infection. Avian Dis. 2020, 64, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ba, G.; Han, Y.Q.; Ming, S.L.; Wang, M.D.; Fu, P.F.; Zhao, Q.Q.; Zhang, S.; Wu, Y.N.; Yang, G.Y.; et al. Cyclic GMP-AMP synthase is essential for cytosolic double-stranded DNA and fowl adenovirus serotype 4 triggered innate immune responses in chickens. Int. J. Biol. Macromol. 2020, 146, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Raheem, M.A.; Han, C.; Yu, F.; Dai, Y.; Imran, M.; Hong, Q.; Zhang, J.; Tan, Y.; Zha, L.; et al. The fowl adenovirus serotype 4 (FAdV-4) induce cellular pathway in chickens to produce interferon and antigen-presented molecules (MHCI/II). Poult. Sci. 2021, 100, 101406. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Chen, S.; Wang, M.; Cheng, A. Interferons and their receptors in birds: A comparison of gene structure, phylogenetic analysis, and cross modulation. Int. J. Mol. Sci. 2014, 15, 21045–21068. [Google Scholar] [CrossRef] [PubMed]
- Sick, C.; Schultz, U.; Munster, U.; Meier, J.; Kaspers, B.; Staeheli, P. Promoter structures and differential responses to viral and nonviral inducers of chicken type I interferon genes. J. Biol. Chem. 1998, 273, 9749–9754. [Google Scholar] [CrossRef]
- Ye, L.; Schnepf, D.; Staeheli, P. Interferon-lambda orchestrates innate and adaptive mucosal immune responses. Nat. Rev. Immunol. 2019, 19, 614–625. [Google Scholar] [CrossRef]
- Ank, N.; West, H.; Bartholdy, C.; Eriksson, K.; Thomsen, A.R.; Paludan, S.R. Lambda interferon (IFN-lambda), a type III IFN, is induced by viruses and IFNs and displays potent antiviral activity against select virus infections in vivo. J. Virol. 2006, 80, 4501–4509. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Sun, B.; Zhuo, Y.; Jiang, Z.; Li, R.; Lin, C.; Jin, Y.; Gao, Y.; Wang, D. Interferon and interferon-stimulated genes in HBV treatment. Front. Immunol. 2022, 13, 1034968. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Wu, S.; Feng, M.; Feng, S.; Sun, C.; Bai, D.; Gu, M.; Liao, M.; Cao, W. Recombinant chicken interferon-alpha inhibits the replication of exogenous avian leukosis virus (ALV) in DF-1 cells. Mol. Immunol. 2016, 76, 62–69. [Google Scholar] [CrossRef]
- Xia, C.; Liu, J.; Wu, Z.G.; Lin, C.Y.; Wang, M. The interferon-alpha genes from three chicken lines and its effects on H9N2 influenza viruses. Anim. Biotechnol. 2004, 15, 77–88. [Google Scholar] [CrossRef]
- Mo, C.W.; Cao, Y.C.; Lim, B.L. The in vivo and in vitro effects of chicken interferon alpha on infectious bursal disease virus and Newcastle disease virus infection. Avian Dis. 2001, 45, 389–399. [Google Scholar] [CrossRef]
- Levy, A.M.; Heller, E.D.; Leitner, G.; Davidson, I. Effect of native chicken interferon on MDV replication. Acta Virol. 1999, 43, 121–127. [Google Scholar]
- Li, L.; Fu, F.; Xue, M.; Chen, W.; Liu, J.; Shi, H.; Chen, J.; Bu, Z.; Feng, L.; Liu, P. IFN-lambda preferably inhibits PEDV infection of porcine intestinal epithelial cells compared with IFN-alpha. Antivir. Res. 2017, 140, 76–82. [Google Scholar] [CrossRef]
- Reboul, J.; Gardiner, K.; Monneron, D.; Uze, G.; Lutfalla, G. Comparative genomic analysis of the interferon/interleukin-10 receptor gene cluster. Genome Res. 1999, 9, 242–250. [Google Scholar] [CrossRef] [PubMed]
- International Chicken Genome Sequencing, C. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 2004, 432, 695–716. [Google Scholar] [CrossRef] [PubMed]
- Santhakumar, D.; Rubbenstroth, D.; Martinez-Sobrido, L.; Munir, M. Avian Interferons and Their Antiviral Effectors. Front. Immunol. 2017, 8, 49. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, T.; Nomura, K.; Hirayama, Y.; Kitagawa, T. Establishment and characterization of a chicken hepatocellular carcinoma cell line, LMH. Cancer Res. 1987, 47, 4460–4464. [Google Scholar] [PubMed]
- Zhang, J.; Zou, Z.; Huang, K.; Lin, X.; Chen, H.; Jin, M. Insights into leghorn male hepatocellular cells response to fowl adenovirus serotype 4 infection by transcriptome analysis. Vet. Microbiol. 2018, 214, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Darnell, J.E., Jr.; Kerr, I.M.; Stark, G.R. Jak-STAT pathways and transcriptional activation in response to IFNs and other extracellular signaling proteins. Science 1994, 264, 1415–1421. [Google Scholar] [CrossRef]
- Hornung, V.; Hartmann, R.; Ablasser, A.; Hopfner, K.P. OAS proteins and cGAS: Unifying concepts in sensing and responding to cytosolic nucleic acids. Nat. Rev. Immunol. 2014, 14, 521–528. [Google Scholar] [CrossRef]
- Seo, J.Y.; Yaneva, R.; Cresswell, P. Viperin: A multifunctional, interferon-inducible protein that regulates virus replication. Cell Host Microbe 2011, 10, 534–539. [Google Scholar] [CrossRef]
- Haller, O.; Staeheli, P.; Schwemmle, M.; Kochs, G. Mx GTPases: Dynamin-like antiviral machines of innate immunity. Trends Microbiol. 2015, 23, 154–163. [Google Scholar] [CrossRef]
- Richardson, R.B.; Ohlson, M.B.; Eitson, J.L.; Kumar, A.; McDougal, M.B.; Boys, I.N.; Mar, K.B.; De La Cruz-Rivera, P.C.; Douglas, C.; Konopka, G.; et al. A CRISPR screen identifies IFI6 as an ER-resident interferon effector that blocks flavivirus replication. Nat. Microbiol. 2018, 3, 1214–1223. [Google Scholar] [CrossRef]
- Zhou, X.; Michal, J.J.; Zhang, L.; Ding, B.; Lunney, J.K.; Liu, B.; Jiang, Z. Interferon induced IFIT family genes in host antiviral defense. Int. J. Biol. Sci. 2013, 9, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Schachner, A.; Matos, M.; Grafl, B.; Hess, M. Fowl adenovirus-induced diseases and strategies for their control—A review on the current global situation. Avian Pathol. 2018, 47, 111–126. [Google Scholar] [CrossRef]
- Bordi, L.; Lalle, E.; Caglioti, C.; Travaglini, D.; Lapa, D.; Marsella, P.; Quartu, S.; Kis, Z.; Arien, K.K.; Huemer, H.P.; et al. Antagonistic antiviral activity between IFN-lambda and IFN-alpha against lethal Crimean-Congo hemorrhagic fever virus in vitro. PLoS ONE 2015, 10, e0116816. [Google Scholar] [CrossRef]
- Felgenhauer, U.; Schoen, A.; Gad, H.H.; Hartmann, R.; Schaubmar, A.R.; Failing, K.; Drosten, C.; Weber, F. Inhibition of SARS-CoV-2 by type I and type III interferons. J. Biol. Chem. 2020, 295, 13958–13964. [Google Scholar] [CrossRef] [PubMed]
- Kumaran, J.; Colamonici, O.R.; Fish, E.N. Structure-function study of the extracellular domain of the human type I interferon receptor (IFNAR)-1 subunit. J. Interferon Cytokine Res. 2000, 20, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Reuter, A.; Soubies, S.; Hartle, S.; Schusser, B.; Kaspers, B.; Staeheli, P.; Rubbenstroth, D. Antiviral activity of lambda interferon in chickens. J. Virol. 2014, 88, 2835–2843. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Xie, T.; Liao, M.; Zhang, X.; Feng, M. Systematic identification of chicken type I, II and III interferon-stimulated genes. Vet. Res. 2020, 51, 70. [Google Scholar] [CrossRef]
- Santhakumar, D.; Rohaim, M.A.; Munir, M. Genome-Wide Classification of Type I, Type II and Type III Interferon-Stimulated Genes in Chicken Fibroblasts. Vaccines 2019, 7, 160. [Google Scholar] [CrossRef]
- Pei, J.; Sekellick, M.J.; Marcus, P.I.; Choi, I.S.; Collisson, E.W. Chicken interferon type I inhibits infectious bronchitis virus replication and associated respiratory illness. J. Interferon Cytokine Res. 2001, 21, 1071–1077. [Google Scholar] [CrossRef]
- Yount, J.S.; Moltedo, B.; Yang, Y.Y.; Charron, G.; Moran, T.M.; Lopez, C.B.; Hang, H.C. Palmitoylome profiling reveals S-palmitoylation-dependent antiviral activity of IFITM3. Nat. Chem. Biol. 2010, 6, 610–614. [Google Scholar] [CrossRef]
- Gao, G.; Guo, X.; Goff, S.P. Inhibition of retroviral RNA production by ZAP, a CCCH-type zinc finger protein. Science 2002, 297, 1703–1706. [Google Scholar] [CrossRef]
- Munir, M.; Berg, M. The multiple faces of proteinkinase R in antiviral defense. Virulence 2013, 4, 85–89. [Google Scholar] [CrossRef]
- Tan, K.S.; Olfat, F.; Phoon, M.C.; Hsu, J.P.; Howe, J.L.C.; Seet, J.E.; Chin, K.C.; Chow, V.T.K. In vivo and in vitro studies on the antiviral activities of viperin against influenza H1N1 virus infection. J. Gen. Virol. 2012, 93, 1269–1277. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; You, G.; Haiyilati, A.; Wang, H.; Jiao, H.; Wang, Y.; Gao, L.; Cao, H.; Li, X.; Zheng, S.J. Critical Role of Viral Protein Hexon in Hypervirulent Fowl Adenovirus Serotype-4-Induced Autophagy by Interaction with BAG3 and Promotion of Viral Replication in LMH Cells. J. Virol. 2023, 97, e0028423. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Zhu, F.; Shen, P. Expression and purification of chicken beta interferon and its antivirus immunological activity. Protein Expr. Purif. 2012, 84, 123–129. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence(5′-3′) |
|---|---|
| Primers for gene cloning | |
| ChIFN-α-F | CCGGAATTCTGCAACCACCTTC |
| ChIFN-α-R | CCCAAGCTTCTAAGTGCGCGTGTTGCC |
| ChIFN-λ-F | CCGGAATTCCAGGTCACCCCGAAGAA |
| ChIFN-λ-R | CCCAAGCTTCTAAGTGCAATCCTCGCGCTGGGC |
| Primers for RT-qPCR | |
| Q-Hexon-F | CGAGGACTACGACGATTA |
| Q-Hexon-R | CGTGATACAGCAGGTTAATG |
| Q-MX1-F | AAGCCTGAGCATGAGCAGAA |
| Q-MX1-R | TCTCAGGCTGTCAACAAGATCAA |
| Q-OASL-F | ACATCCTCGCCATCATCGA |
| Q-OASL-R | GCGGACTGGTGATGCTGACT |
| Q-IFIT5-F | TGCTCTGAGGGAAGAACCCAACA |
| Q-IFIT5-R | AGGCTCCAGGGATGAGTCCACTT |
| Q-Viperin-F | AACGGTGGTTCAAGAAGTATGG |
| Q-Viperin-R | ACAGCATAATCTCGGCACCA |
| Q-IFITM3-F | TGGTGACGGTGGAGACG |
| Q-IFITM3-R | GGCAACCAGGGCGATGA |
| Q-ZAP-F | TTCCAAGTCAAGCCTGTCCC |
| Q-ZAP-R | CTCCGCTCTGCCTCTTCATC |
| Q-PKR-F | TGACTTCTGTGACATACAACCCTC |
| Q-PKR-R | TTTCAAACCAAATCAATCCC |
| Q-IRF7-F | AACGACGACCCGCACAAG |
| Q-IRF7-R | GCAGCAGGTCCAAATCCA |
| Q-IFI6-F | TCAACACACTCCTCAGGCTTTACC |
| Q-IFI6-R | GAACTCCGCCTCCGCAAGAG |
| Q-β-actin-F | CAACACAGTGCTGTCTGGTGGTA |
| Q-β-actin-R | ATCGTACTCCTGCTTGCTGATCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, J.; He, X.; Zhang, R.; Zhang, L.; Chen, L.; He, F.; Li, L.; Yang, L.; Ren, T.; Xiang, B. Chicken Interferon-Alpha and -Lambda Exhibit Antiviral Effects against Fowl Adenovirus Serotype 4 in Leghorn Male Hepatocellular Cells. Int. J. Mol. Sci. 2024, 25, 1681. https://doi.org/10.3390/ijms25031681
Lai J, He X, Zhang R, Zhang L, Chen L, He F, Li L, Yang L, Ren T, Xiang B. Chicken Interferon-Alpha and -Lambda Exhibit Antiviral Effects against Fowl Adenovirus Serotype 4 in Leghorn Male Hepatocellular Cells. International Journal of Molecular Sciences. 2024; 25(3):1681. https://doi.org/10.3390/ijms25031681
Chicago/Turabian StyleLai, Jinyu, Xingchen He, Rongjie Zhang, Limei Zhang, Libin Chen, Fengping He, Lei Li, Liangyu Yang, Tao Ren, and Bin Xiang. 2024. "Chicken Interferon-Alpha and -Lambda Exhibit Antiviral Effects against Fowl Adenovirus Serotype 4 in Leghorn Male Hepatocellular Cells" International Journal of Molecular Sciences 25, no. 3: 1681. https://doi.org/10.3390/ijms25031681
APA StyleLai, J., He, X., Zhang, R., Zhang, L., Chen, L., He, F., Li, L., Yang, L., Ren, T., & Xiang, B. (2024). Chicken Interferon-Alpha and -Lambda Exhibit Antiviral Effects against Fowl Adenovirus Serotype 4 in Leghorn Male Hepatocellular Cells. International Journal of Molecular Sciences, 25(3), 1681. https://doi.org/10.3390/ijms25031681