
Myelin Basic Protein Attenuates Furin-Mediated Bri2 Cleavage and Postpones Its Membrane Trafficking
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
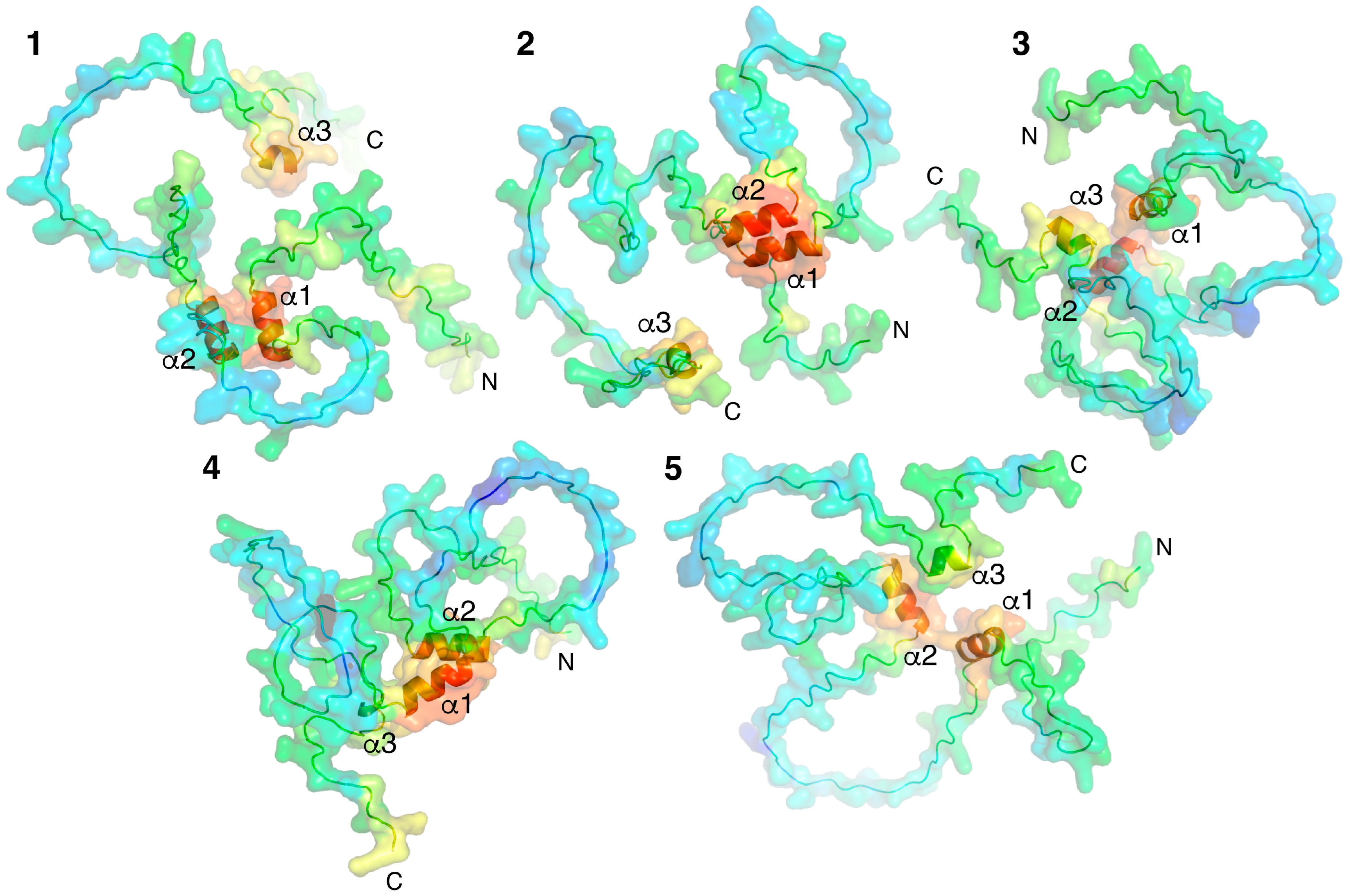
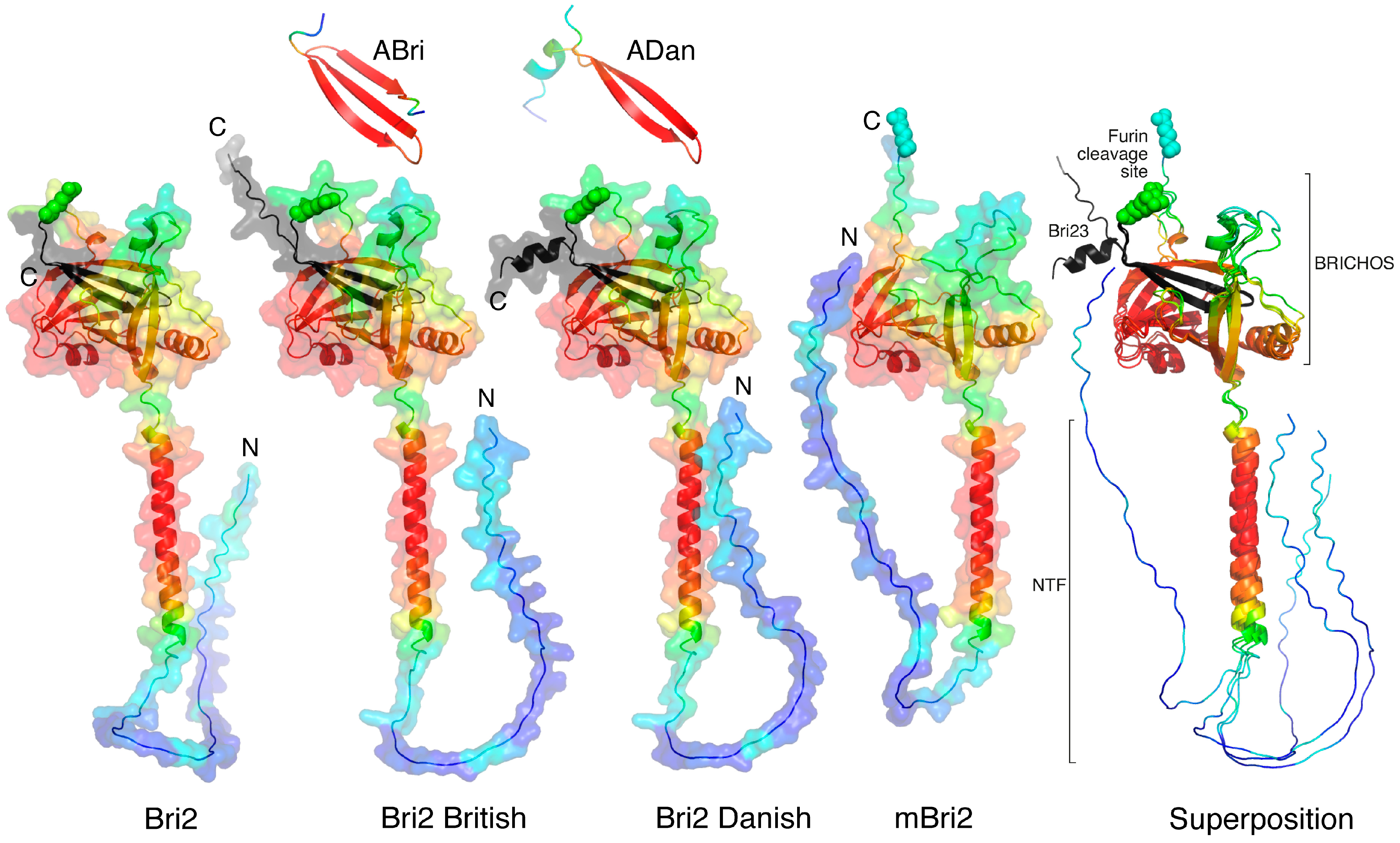
2.1. Molecular Modeling of Individual Proteins Using AlphaFold2
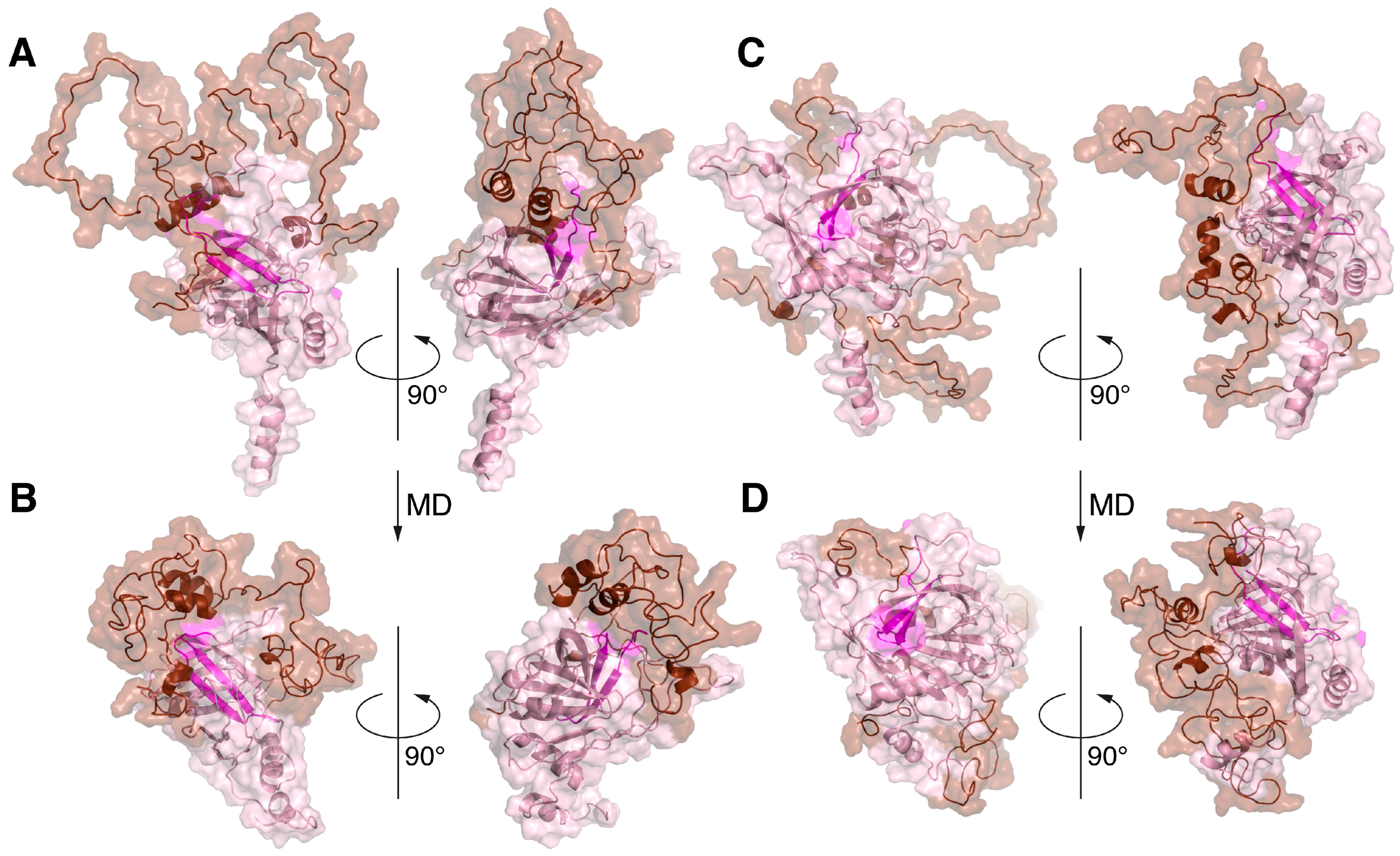
2.2. Simulation of the Intermolecular Interactions during Co-Folding of Two Proteins Using AlphaFold-Multimer
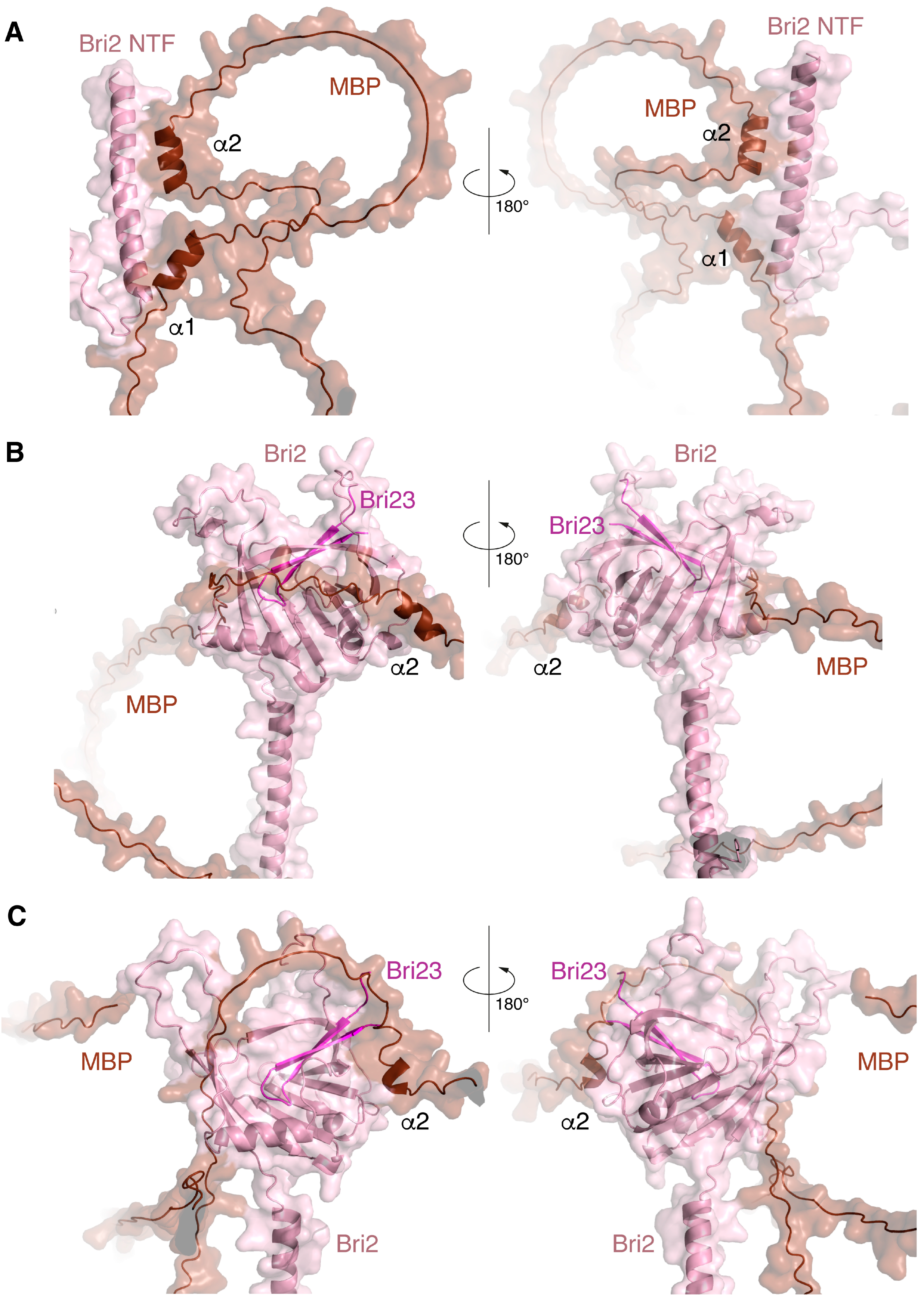
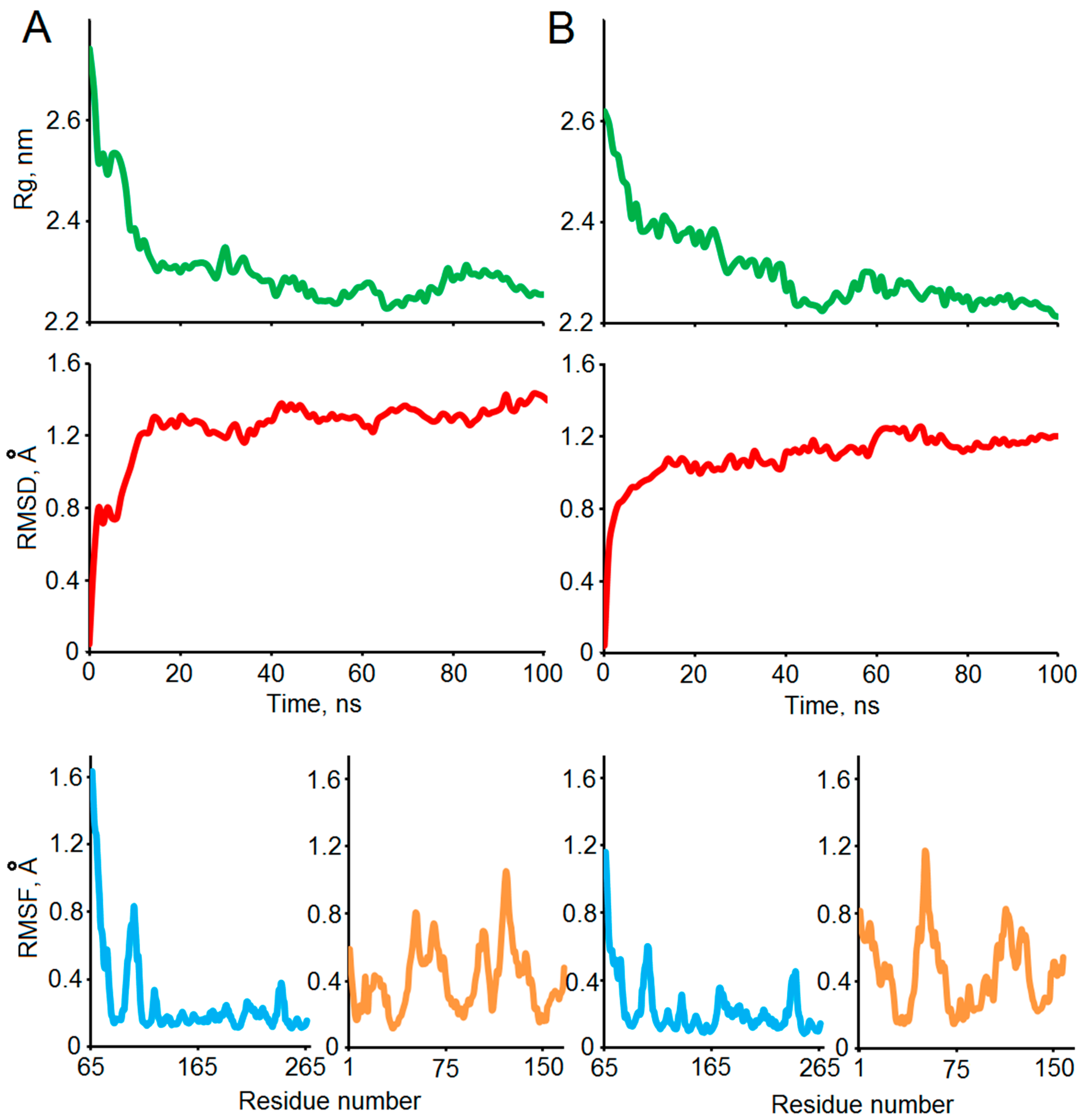
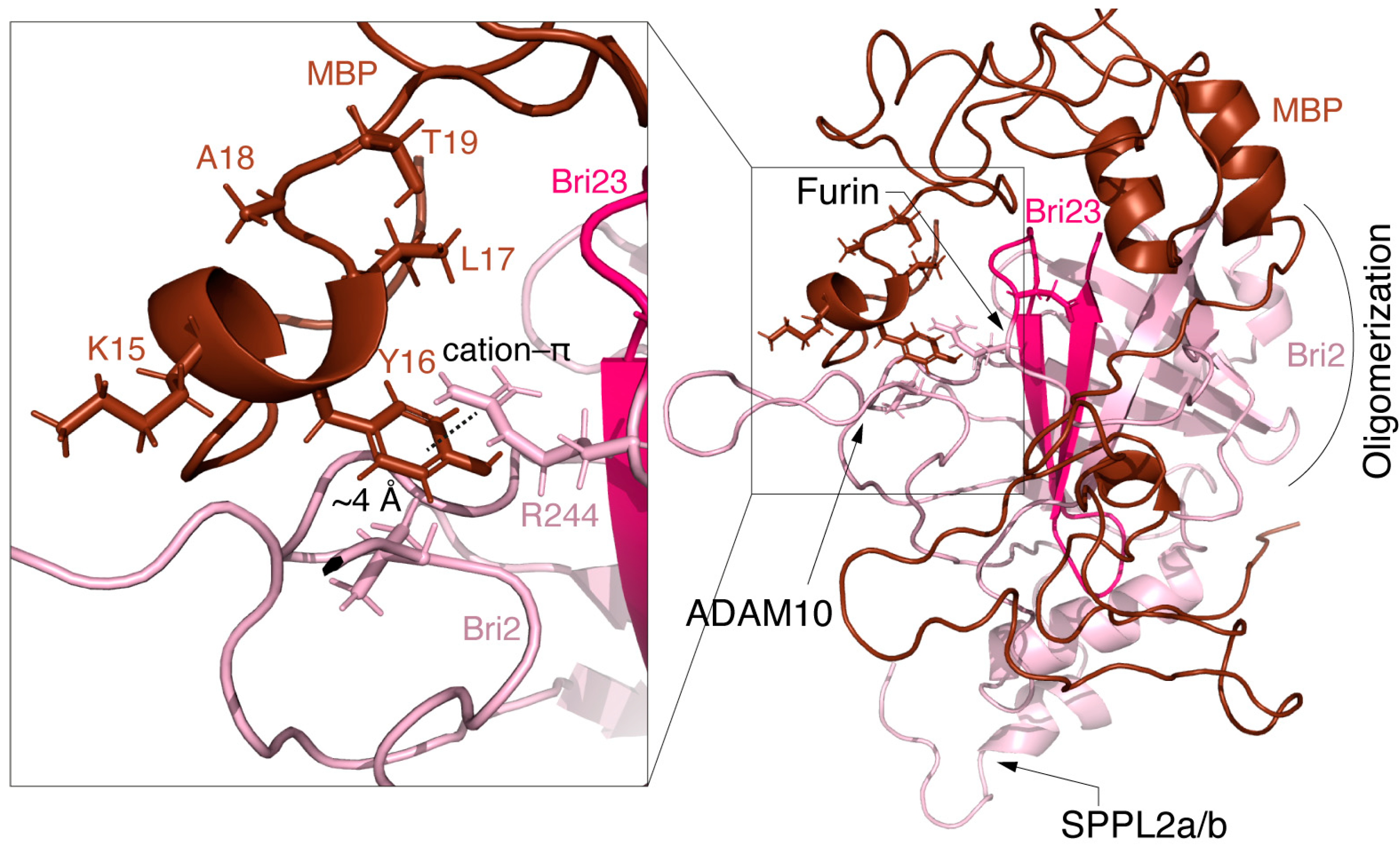
2.3. Protein–Protein Docking and MD Simulation of the MBP-Bri2Δ64 Complexes
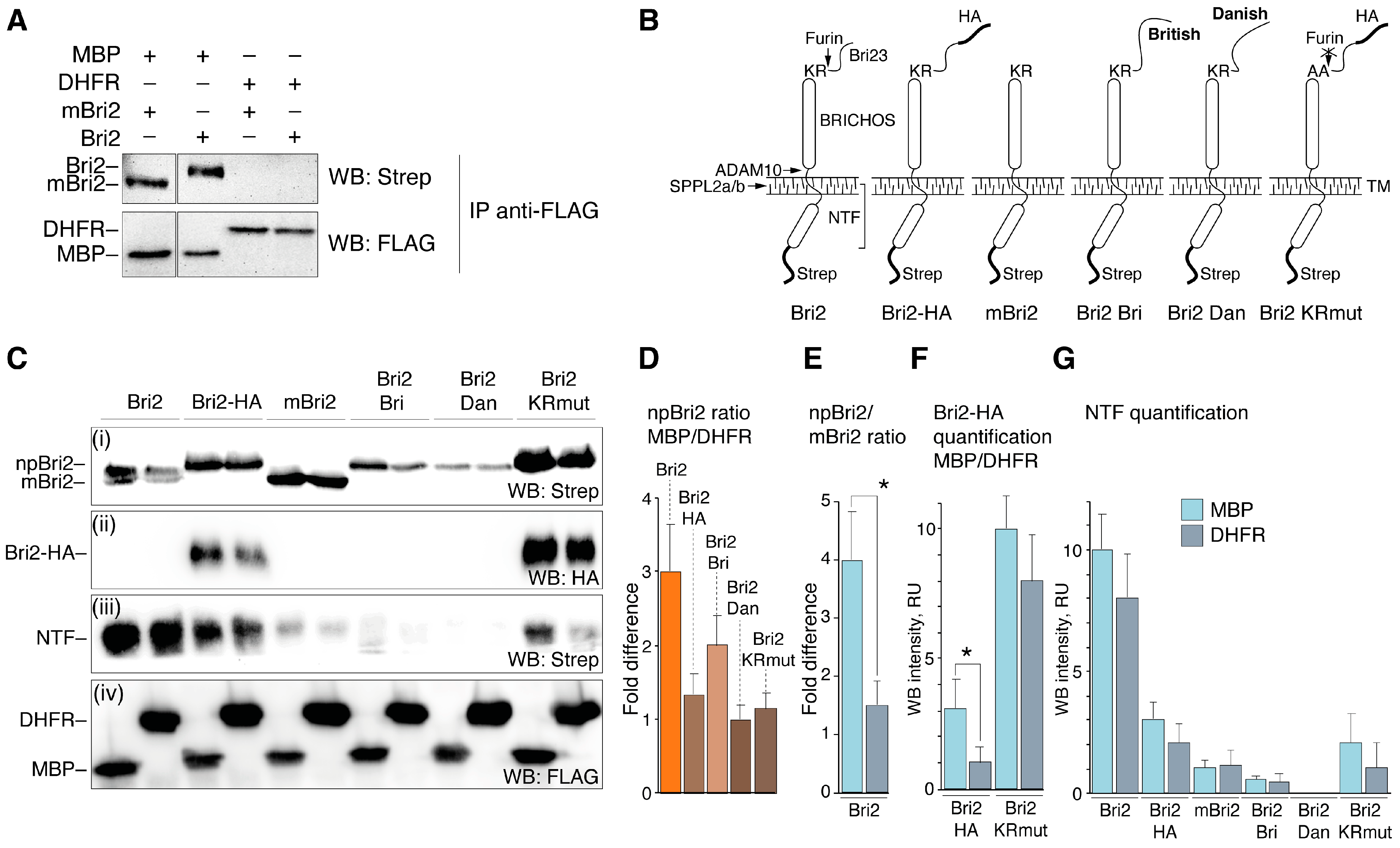
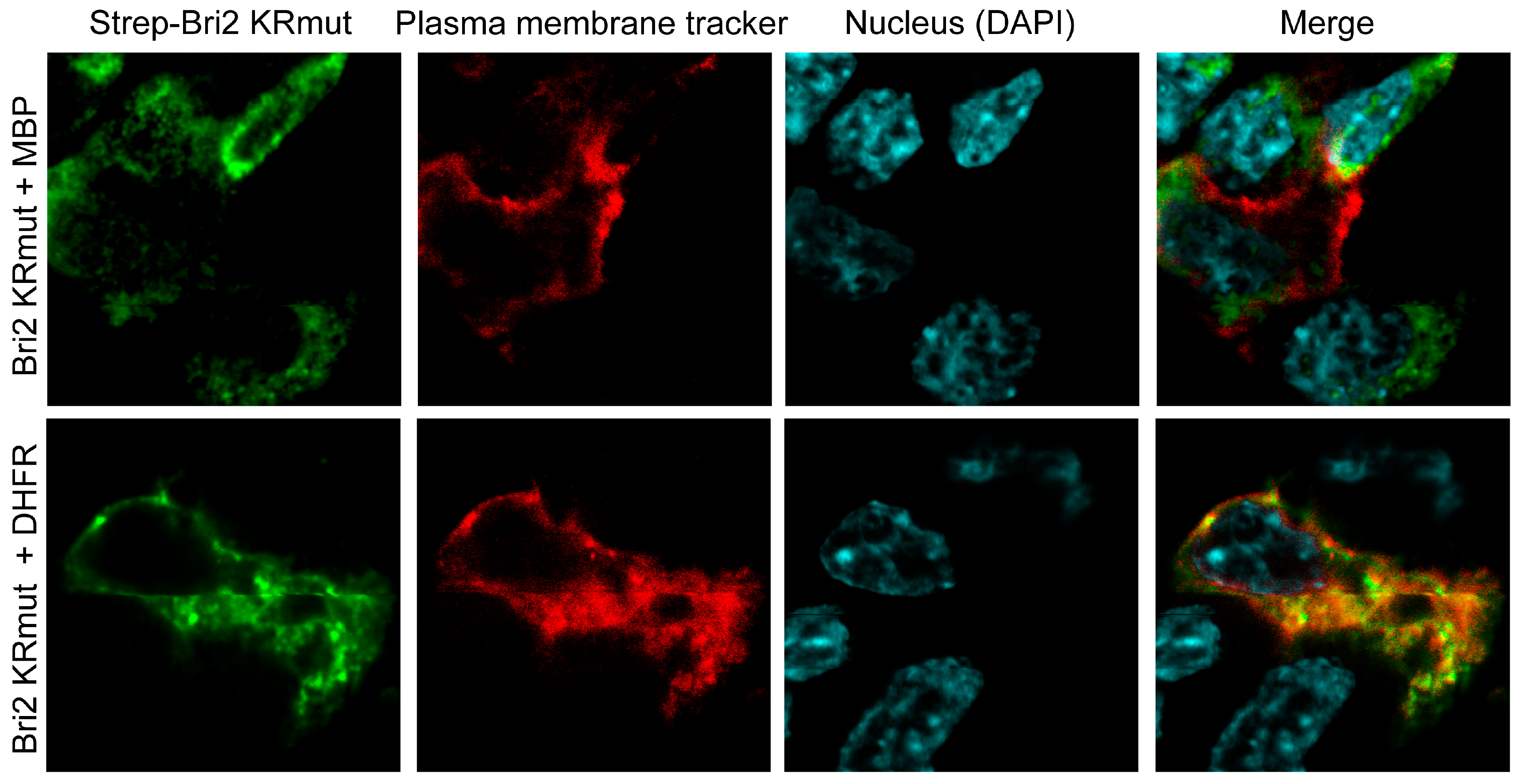
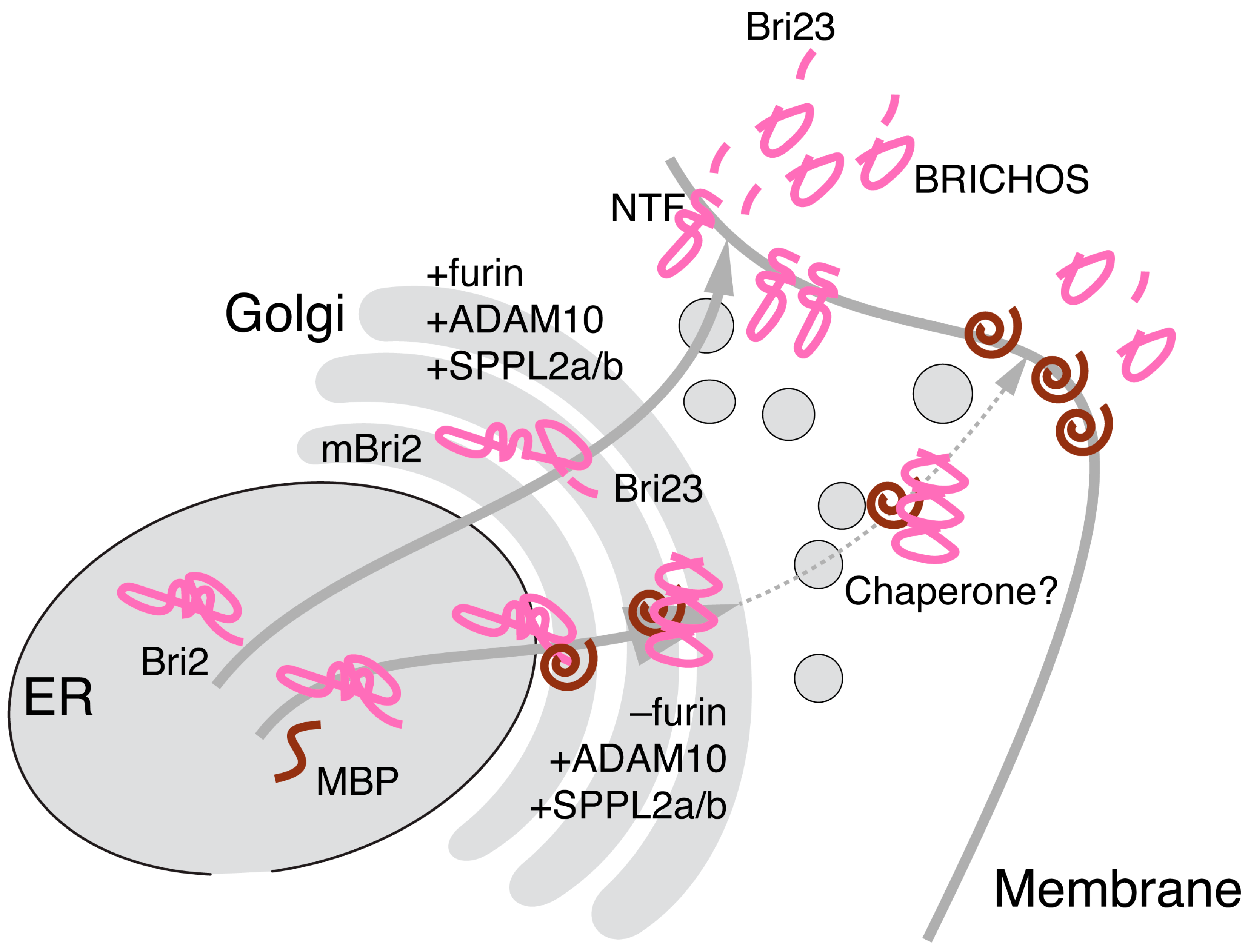
2.4. MBP Modulates Post-Translational Bri2 Processing and Affects Its Intracellular Localization by Restriction of Furin-Catalyzed Release of Its C-Terminal Peptide
3. Materials and Methods
3.1. Plasmids Coding for Bri2 Variants and MBP
3.2. Transient Transfection of Mammalian Cells
3.3. Antibodies
3.4. Immunoprecipitation and Immunoblotting
3.5. Immunofluorescence, Image Acquisition, and Analysis in Cell Culture
3.6. AlphaFold2 and AlphaFold-Multimer Based Modeling
3.7. Molecular Docking
3.8. Molecular Dynamics of Obtained Complexes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, D.; Tsai, L.-H. Bridging Physiology and Pathology in AD. Cell 2009, 137, 997–1000. [Google Scholar] [CrossRef]
- Benson, M.D.; Buxbaum, J.N.; Eisenberg, D.S.; Merlini, G.; Saraiva, M.J.M.; Sekijima, Y.; Sipe, J.D.; Westermark, P. Amyloid Nomenclature 2018: Recommendations by the International Society of Amyloidosis (ISA) Nomenclature Committee. Amyloid 2018, 25, 215–219. [Google Scholar] [CrossRef]
- Hardy, J.A.; Higgins, G.A. Alzheimer’s Disease: The Amyloid Cascade Hypothesis. Science 1992, 256, 184–185. [Google Scholar] [CrossRef]
- Hardy, J. The Discovery of Alzheimer-causing Mutations in the APP Gene and the Formulation of the “Amyloid Cascade Hypothesis”. FEBS J. 2017, 284, 1040–1044. [Google Scholar] [CrossRef]
- Alzheimer Disease Genetics Consortium (ADGC); The European Alzheimer’s Disease Initiative (EADI); Cohorts for Heart and Aging Research in Genomic Epidemiology Consortium (CHARGE); Genetic and Environmental Risk in AD/Defining Genetic, Polygenic and Environmental Risk for Alzheimer’s Disease Consortium (GERAD/PERADES); Kunkle, B.W.; Grenier-Boley, B.; Sims, R.; Bis, J.C.; Damotte, V.; Naj, A.C.; et al. Genetic Meta-Analysis of Diagnosed Alzheimer’s Disease Identifies New Risk Loci and Implicates Aβ, Tau, Immunity and Lipid Processing. Nat. Genet. 2019, 51, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Balchin, D.; Hayer-Hartl, M.; Hartl, F.U. Recent Advances in Understanding Catalysis of Protein Folding by Molecular Chaperones. FEBS Lett. 2020, 594, 2770–2781. [Google Scholar] [CrossRef] [PubMed]
- Deleersnijder, W.; Hong, G.; Cortvrindt, R.; Poirier, C.; Tylzanowski, P.; Pittois, K.; Van Marck, E.; Merregaert, J. Isolation of Markers for Chondro-Osteogenic Differentiation Using cDNA Library Subtraction. J. Biol. Chem. 1996, 271, 19475–19482. [Google Scholar] [CrossRef] [PubMed]
- Fotinopoulou, A.; Tsachaki, M.; Vlavaki, M.; Poulopoulos, A.; Rostagno, A.; Frangione, B.; Ghiso, J.; Efthimiopoulos, S. BRI2 Interacts with Amyloid Precursor Protein (APP) and Regulates Amyloid β (Aβ) Production. J. Biol. Chem. 2005, 280, 30768–30772. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, S.; Giliberto, L.; Matsuda, Y.; Davies, P.; McGowan, E.; Pickford, F.; Ghiso, J.; Frangione, B.; D’Adamio, L. The Familial Dementia BRI2 Gene Binds the Alzheimer Gene Amyloid-β Precursor Protein and Inhibits Amyloid-β Production. J. Biol. Chem. 2005, 280, 28912–28916. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, S.; Giliberto, L.; Matsuda, Y.; McGowan, E.M.; D’Adamio, L. BRI2 Inhibits Amyloid β-Peptide Precursor Protein Processing by Interfering with the Docking of Secretases to the Substrate. J. Neurosci. 2008, 28, 8668–8676. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Wang, R.; Gordon, D.J.; Bass, J.; Steiner, D.F.; Lynn, D.G.; Thinakaran, G.; Meredith, S.C.; Sisodia, S.S. Furin Mediates Enhanced Production of Fibrillogenic ABri Peptides in Familial British Dementia. Nat. Neurosci. 1999, 2, 984–988. [Google Scholar] [CrossRef]
- Kim, S.-H.; Creemers, J.W.M.; Chu, S.; Thinakaran, G.; Sisodia, S.S. Proteolytic Processing of Familial British Dementia-Associated BRI Variants. J. Biol. Chem. 2002, 277, 1872–1877. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Pulido, L.; Devos, D.; Valencia, A. BRICHOS: A Conserved Domain in Proteins Associated with Dementia, Respiratory Distress and Cancer. Trends Biochem. Sci. 2002, 27, 329–332. [Google Scholar] [CrossRef]
- Martin, L.; Fluhrer, R.; Reiss, K.; Kremmer, E.; Saftig, P.; Haass, C. Regulated Intramembrane Proteolysis of Bri2 (Itm2b) by ADAM10 and SPPL2a/SPPL2b. J. Biol. Chem. 2008, 283, 1644–1652. [Google Scholar] [CrossRef] [PubMed]
- Arosio, P.; Michaels, T.C.T.; Linse, S.; Månsson, C.; Emanuelsson, C.; Presto, J.; Johansson, J.; Vendruscolo, M.; Dobson, C.M.; Knowles, T.P.J. Kinetic Analysis Reveals the Diversity of Microscopic Mechanisms through Which Molecular Chaperones Suppress Amyloid Formation. Nat. Commun. 2016, 7, 10948. [Google Scholar] [CrossRef] [PubMed]
- Poska, H.; Haslbeck, M.; Kurudenkandy, F.R.; Hermansson, E.; Chen, G.; Kostallas, G.; Abelein, A.; Biverstål, H.; Crux, S.; Fisahn, A.; et al. Dementia-Related Bri2 BRICHOS Is a Versatile Molecular Chaperone That Efficiently Inhibits Aβ42 Toxicity in Drosophila. Biochem. J. 2016, 473, 3683–3704. [Google Scholar] [CrossRef]
- Chen, G.; Abelein, A.; Nilsson, H.E.; Leppert, A.; Andrade-Talavera, Y.; Tambaro, S.; Hemmingsson, L.; Roshan, F.; Landreh, M.; Biverstål, H.; et al. Bri2 BRICHOS Client Specificity and Chaperone Activity Are Governed by Assembly State. Nat. Commun. 2017, 8, 2081. [Google Scholar] [CrossRef]
- Manchanda, S.; Galan-Acosta, L.; Abelein, A.; Tambaro, S.; Chen, G.; Nilsson, P.; Johansson, J. Intravenous Treatment with a Molecular Chaperone Designed against β-Amyloid Toxicity Improves Alzheimer’s Disease Pathology in Mouse Models. Mol. Ther. 2023, 31, 487–502. [Google Scholar] [CrossRef]
- Kim, J.; Miller, V.M.; Levites, Y.; West, K.J.; Zwizinski, C.W.; Moore, B.D.; Troendle, F.J.; Bann, M.; Verbeeck, C.; Price, R.W.; et al. BRI2 (ITM2b) Inhibits A Deposition In Vivo. J. Neurosci. 2008, 28, 6030–6036. [Google Scholar] [CrossRef]
- Vidal, R.; Frangione, B.; Rostagno, A.; Mead, S.; Révész, T.; Plant, G.; Ghiso, J. A Stop-Codon Mutation in the BRI Gene Associated with Familial British Dementia. Nature 1999, 399, 776–781. [Google Scholar] [CrossRef]
- Vidal, R.; Révész, T.; Rostagno, A.; Kim, E.; Holton, J.L.; Bek, T.; Bojsen-Møller, M.; Braendgaard, H.; Plant, G.; Ghiso, J.; et al. A Decamer Duplication in the 3′ Region of the BRI Gene Originates an Amyloid Peptide That Is Associated with Dementia in a Danish Kindred. Proc. Natl. Acad. Sci. USA 2000, 97, 4920–4925. [Google Scholar] [CrossRef] [PubMed]
- Rhyu, J.-M.; Park, J.; Shin, B.-S.; Kim, Y.-E.; Kim, E.-J.; Kim, K.W.; Cho, Y.G. A Novel c.800G>C Variant of the ITM2B Gene in Familial Korean Dementia. JAD 2023, 93, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, K.-L.; Wang, Y.; Huang, Y.-Y.; Chen, S.-D.; Dong, Q.; Cui, M.; Yu, J.-T. A Novel ITM2B Mutation Associated with Familial Chinese Dementia. JAD 2021, 81, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Leppert, A.; Poska, H.; Landreh, M.; Abelein, A.; Chen, G.; Johansson, J. A New Kid in the Folding Funnel: Molecular Chaperone Activities of the BRICHOS Domain. Protein Sci. 2023, 32, e4645. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, E.V.; Rakitina, T.V.; Saratov, G.A.; Kudriaeva, A.A.; Belogurov, A.A. Deconvolution of the MBP-Bri2 Interaction by a Yeast Two Hybrid System and Synergy of the AlphaFold2 and High Ambiguity Driven Protein-Protein Docking. Crystals 2022, 12, 197. [Google Scholar] [CrossRef]
- Eylar, E.H.; Salk, J.; Beveridge, G.C.; Brown, L.V. Experimental Allergic Encephalomyelitis. Arch. Biochem. Biophys. 1969, 132, 34–48. [Google Scholar] [CrossRef]
- Ryberg, B. Multiple Specificities of Antibrain Antibodies in Multiple Sclerosis and Chronic Myelopathy. J. Neurol. Sci. 1978, 38, 357–382. [Google Scholar] [CrossRef]
- Panitch, H.S. CSF Antibody to Myelin Basic Protein: Measurement in Patients with Multiple Sclerosis and Subacute Sclerosing Panencephalitis. Arch. Neurol 1980, 37, 206. [Google Scholar] [CrossRef]
- Górny, M.K.; Wróblewska, Z.; Pleasure, D.; Miller, S.L.; Wajgt, A.; Koprowski, H. CSF Antibodies to Myelin Basic Protein and Oligodendrocytes in Multiple Sclerosis and Other Neurological Diseases. Acta Neurol. Scand. 2009, 67, 338–347. [Google Scholar] [CrossRef]
- Doolittle, D.P.; Schweikart, K.M. Myelin Deficient, a New Neurological Mutant in the Mouse. J. Hered. 1977, 68, 331–332. [Google Scholar] [CrossRef]
- Popko, B.; Puckett, C.; Hood, L. A Novel Mutation in Myelin-Deficient Mice Results in Unstable Myelin Basic Protein Gene Transcripts. Neuron 1988, 1, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Omlin, F.X. Immunocytochemical Localization of Basic Protein in Major Dense Line Regions of Central and Peripheral Myelin. J. Cell Biol. 1982, 95, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Readhead, C.; Takasashi, N.; Shine, H.D.; Saavedra, R.; Sidman, R.; Hood, L. Role of Myelin Basic Protein in the Formation of Central Nervous System Myelin. Ann. N. Y. Acad. Sci. 1990, 605, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Harauz, G.; Ladizhansky, V.; Boggs, J.M. Structural Polymorphism and Multifunctionality of Myelin Basic Protein. Biochemistry 2009, 48, 8094–8104. [Google Scholar] [CrossRef] [PubMed]
- Hoos, M.D.; Ahmed, M.; Smith, S.O.; Van Nostrand, W.E. Inhibition of Familial Cerebral Amyloid Angiopathy Mutant Amyloid β-Protein Fibril Assembly by Myelin Basic Protein. J. Biol. Chem. 2007, 282, 9952–9961. [Google Scholar] [CrossRef] [PubMed]
- Hoos, M.D.; Ahmed, M.; Smith, S.O.; Van Nostrand, W.E. Myelin Basic Protein Binds to and Inhibits the Fibrillar Assembly of Aβ42 in Vitro. Biochemistry 2009, 48, 4720–4727. [Google Scholar] [CrossRef]
- Liao, M.-C.; Hoos, M.D.; Aucoin, D.; Ahmed, M.; Davis, J.; Smith, S.O.; Van Nostrand, W.E. N-Terminal Domain of Myelin Basic Protein Inhibits Amyloid β-Protein Fibril Assembly. J. Biol. Chem. 2010, 285, 35590–35598. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Martins, F.; Marafona, A.M.; Pereira, C.D.; Müller, T.; Loosse, C.; Kolbe, K.; da Cruz e Silva, O.A.B.; Rebelo, S. Identification and Characterization of the BRI2 Interactome in the Brain. Sci. Rep. 2018, 8, 3548. [Google Scholar] [CrossRef]
- Pool, M.R. Targeting of Proteins for Translocation at the Endoplasmic Reticulum. Int. J. Mol. Sci. 2022, 23, 3773. [Google Scholar] [CrossRef]
- Shao, S.; Rodrigo-Brenni, M.C.; Kivlen, M.H.; Hegde, R.S. Mechanistic Basis for a Molecular Triage Reaction. Science 2017, 355, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.; Bauer, N.M.; Schäfer, I.; White, R. Making Myelin Basic Protein -from mRNA Transport to Localized Translation. Front. Cell. Neurosci. 2013, 7, 169. [Google Scholar] [CrossRef]
- Shao, S.; Hegde, R.S. A Calmodulin-Dependent Translocation Pathway for Small Secretory Proteins. Cell 2011, 147, 1576–1588. [Google Scholar] [CrossRef]
- Smirnova, E.V.; Rakitina, T.V.; Ziganshin, R.H.; Arapidi, G.P.; Saratov, G.A.; Kudriaeva, A.A.; Belogurov, A.A. Comprehensive Atlas of the Myelin Basic Protein Interaction Landscape. Biomolecules 2021, 11, 1628. [Google Scholar] [CrossRef] [PubMed]
- Senior, A.W.; Evans, R.; Jumper, J.; Kirkpatrick, J.; Sifre, L.; Green, T.; Qin, C.; Žídek, A.; Nelson, A.W.R.; Bridgland, A.; et al. Protein Structure Prediction Using Multiple Deep Neural Networks in the 13th Critical Assessment of Protein Structure Prediction (CASP13). Proteins 2019, 87, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- de Vries, S.J.; van Dijk, A.D.J.; Krzeminski, M.; van Dijk, M.; Thureau, A.; Hsu, V.; Wassenaar, T.; Bonvin, A.M.J.J. HADDOCK versus HADDOCK: New Features and Performance of HADDOCK2.0 on the CAPRI Targets. Proteins 2007, 69, 726–733. [Google Scholar] [CrossRef]
- Bates, I.R.; Libich, D.S.; Wood, D.D.; Moscarello, M.A.; Harauz, G. An Arg/Lys→Gln Mutant of Recombinant Murine Myelin Basic Protein as a Mimic of the Deiminated Form Implicated in Multiple Sclerosis. Protein Expr. Purif. 2002, 25, 330–341. [Google Scholar] [CrossRef]
- Fernández-Recio, J.; Totrov, M.; Abagyan, R. Identification of Protein–Protein Interaction Sites from Docking Energy Landscapes. J. Mol. Biol. 2004, 335, 843–865. [Google Scholar] [CrossRef]
- Barbarese, E.; Pfeiffer, S.E. Developmental Regulation of Myelin Basic Protein in Dispersed Cultures. Proc. Natl. Acad. Sci. USA 1981, 78, 1953–1957. [Google Scholar] [CrossRef]
- Happel, P.; Möller, K.; Schwering, N.K.; Dietzel, I.D. Migrating Oligodendrocyte Progenitor Cells Swell Prior to Soma Dislocation. Sci. Rep. 2013, 3, 1806. [Google Scholar] [CrossRef]
- Kudriaeva, A.; Kuzina, E.S.; Zubenko, O.; Smirnov, I.V.; Belogurov, A. Charge-Mediated Proteasome Targeting. FASEB J. 2019, 33, 6852–6866. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, C.; Boelens, R.; Bonvin, A.M.J.J. HADDOCK: A Protein−Protein Docking Approach Based on Biochemical or Biophysical Information. J. Am. Chem. Soc. 2003, 125, 1731–1737. [Google Scholar] [CrossRef] [PubMed]
- Mitternacht, S. FreeSASA: An Open Source C Library for Solvent Accessible Surface Area Calculations. F1000Research 2016, 5, 189. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of Simple Potential Functions for Simulating Liquid Water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, Flexible, and Free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef] [PubMed]
- Lindorff-Larsen, K.; Piana, S.; Palmo, K.; Maragakis, P.; Klepeis, J.L.; Dror, R.O.; Shaw, D.E. Improved Side-Chain Torsion Potentials for the Amber ff99SB Protein Force Field: Improved Protein Side-Chain Potentials. Proteins 2010, 78, 1950–1958. [Google Scholar] [CrossRef] [PubMed]
- Horn, H.W.; Swope, W.C.; Pitera, J.W.; Madura, J.D.; Dick, T.J.; Hura, G.L.; Head-Gordon, T. Development of an Improved Four-Site Water Model for Biomolecular Simulations: TIP4P-Ew. J. Chem. Phys. 2004, 120, 9665–9678. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; Van Gunsteren, W.F.; DiNola, A.; Haak, J.R. Molecular Dynamics with Coupling to an External Bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef]
- Parrinello, M.; Rahman, A. Polymorphic Transitions in Single Crystals: A New Molecular Dynamics Method. J. Appl. Phys. 1981, 52, 7182–7190. [Google Scholar] [CrossRef]
- Van Gunsteren, W.F.; Berendsen, H.J.C. A Leap-Frog Algorithm for Stochastic Dynamics. Mol. Simul. 1988, 1, 173–185. [Google Scholar] [CrossRef]
- Chen, G.; Andrade-Talavera, Y.; Tambaro, S.; Leppert, A.; Nilsson, H.E.; Zhong, X.; Landreh, M.; Nilsson, P.; Hebert, H.; Biverstål, H.; et al. Augmentation of Bri2 Molecular Chaperone Activity against Amyloid-β Reduces Neurotoxicity in Mouse Hippocampus in Vitro. Commun. Biol. 2020, 3, 32. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MBP-Bri2 ICD Complexes | |||||
| 1 | 2 | 3 | 4 | 5 | |
| Interface area *, Å2 | 552 | 342.7 | 484.3 | - | - |
| ΔG **, kcal/mol | −11.0 | 0.7 | 1.1 | - | - |
| ΔG, p-value | 0.235 | 0.798 | 0.867 | - | - |
| Hydrogen bonds | 0 | 4 | 4 | - | - |
| Salt Bridges | 0 | 0 | 5 | - | - |
| MBP-Bri2 NTF Complexes | |||||
| 1 | 2 | 3 | 4 | 5 | |
| Interface area *, Å2 | 456.8 | 906.7 | 531.3 | 528.7 | 62.1 |
| ΔG **, kcal/mol | −10.8 | −15.0 | −7.2 | −10.4 | −1.8 |
| ΔG, p-value | 0.299 | 0.481 | 0.503 | 0.394 | 0.289 |
| Hydrogen bonds | 0 | 3 | 4 | 0 | 0 |
| Salt Bridges | 0 | 7 | 1 | 0 | 0 |
| MBP-Bri2 Complexes | |||||
| 1 | 2 | 3 | 4 | 5 | |
| Interface area * Å2 | 820.9 | 979.6 | 1718.9 | 576.3 | 327.9 |
| ΔG **, kcal/mol | −10.2 | −15.0 | −22.5 | −8.8 | −4.0 |
| ΔG, p-value | 0.372 | 0.263 | 0.129 | 0.288 | 0.293 |
| Hydrogen bonds | 7 | 2 | 14 | 0 | 0 |
| Salt Bridges | 0 | 3 | 0 | 0 | 0 |
| MBP | Bri2 | |||||
|---|---|---|---|---|---|---|
| N | AA | Atom | Distance Å | AA | Atom | Contact Type * |
| 2 | Ser116 | N | 3.32 | Gly90 | O | HB |
| 2 | Ser116 | O | 3.08 | Gly90 | N | HB |
| 2 | Arg98 | NE | 3.27 | Glu235 | OE1 | SB |
| 2 | Arg98 | NH1 | 2.53 | Glu235 | OE1 | SB |
| 2 | Arg98 | NH2 | 3.28 | Glu235 | OE2 | SB |
| 3 | Arg108 | NH2 | 3.15 | Asn101 | OD1 | HB |
| 3 | Leu110 | N | 3.81 | Asp96 | OD1 | HB |
| 3 | Leu110 | N | 3.62 | Asp96 | OD2 | HB |
| 3 | Ser111 | N | 3.43 | Asp96 | OD1 | HB |
| 3 | Ser113 | N | 3.45 | Pro194 | O | HB |
| 3 | Ser113 | OG | 2.26 | Ser196 | OG | HB |
| 3 | Ser113 | OG | 3.27 | Ser196 | O | HB |
| 3 | Gly118 | N | 3.77 | Tyr88 | O | HB |
| 3 | Phe90 | O | 3.46 | Lys230 | NZ | HB |
| 3 | Ser111 | O | 3.43 | Ser196 | N | HB |
| 3 | Ser111 | O | 3.46 | Tyr197 | N | HB |
| 3 | Leu112 | O | 3.84 | Ile94 | N | HB |
| 3 | Ser113 | OG | 2.96 | Ser196 | N | HB |
| 3 | Glu120 | OE2 | 3.68 | Tyr88 | OH | HB |
| Intermolecular Energy, kcal/mol | Buried Surface, Å | |||||
|---|---|---|---|---|---|---|
| Model | Total | Vdw 1 | Elec 1 | dH 2 | Edesol 3 | |
| 1–25 | −414.6 | −97.8 | −316.8 | −344.4 | −37.3 | 4042.2 |
| 5–6 | −473.6 | −86.8 | −386.8 | −352.5 | −24.2 | 3802.7 |
| Complex | 1–25 (Figure 5A,B) | 5–6 (Figure 5C,D) | ||
|---|---|---|---|---|
| Characteristics/Structures | HADDOCK | MD | HADDOCK | MD |
| interface area, Å2 | 1921.3 | 2587.0 | 1823 | 3056 |
| ΔG, kcal/mol | −18.9 | −15.9 | −12.0 | −10.2 |
| ΔG, p-value | 0.263 | 0.504 | 0.635 | 0.585 |
| Hydrogen bonds | 15 | 27 | 14 | 44 |
| Salt Bridges | 3 | 10 | 8 | 29 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smirnova, E.V.; Timofeev, V.I.; Rakitina, T.V.; Petrenko, D.E.; Elmeeva, O.S.; Saratov, G.A.; Kudriaeva, A.A.; Bocharov, E.V.; Belogurov, A.A., Jr. Myelin Basic Protein Attenuates Furin-Mediated Bri2 Cleavage and Postpones Its Membrane Trafficking. Int. J. Mol. Sci. 2024, 25, 2608. https://doi.org/10.3390/ijms25052608
Smirnova EV, Timofeev VI, Rakitina TV, Petrenko DE, Elmeeva OS, Saratov GA, Kudriaeva AA, Bocharov EV, Belogurov AA Jr. Myelin Basic Protein Attenuates Furin-Mediated Bri2 Cleavage and Postpones Its Membrane Trafficking. International Journal of Molecular Sciences. 2024; 25(5):2608. https://doi.org/10.3390/ijms25052608
Chicago/Turabian StyleSmirnova, Evgeniya V., Vladimir I. Timofeev, Tatiana V. Rakitina, Dmitry E. Petrenko, Olga S. Elmeeva, George A. Saratov, Anna A. Kudriaeva, Eduard V. Bocharov, and Alexey A. Belogurov, Jr. 2024. "Myelin Basic Protein Attenuates Furin-Mediated Bri2 Cleavage and Postpones Its Membrane Trafficking" International Journal of Molecular Sciences 25, no. 5: 2608. https://doi.org/10.3390/ijms25052608
APA StyleSmirnova, E. V., Timofeev, V. I., Rakitina, T. V., Petrenko, D. E., Elmeeva, O. S., Saratov, G. A., Kudriaeva, A. A., Bocharov, E. V., & Belogurov, A. A., Jr. (2024). Myelin Basic Protein Attenuates Furin-Mediated Bri2 Cleavage and Postpones Its Membrane Trafficking. International Journal of Molecular Sciences, 25(5), 2608. https://doi.org/10.3390/ijms25052608