

Histochemical Evidence for Reduced Immune Response in Nasal Mucosa of Patients with COVID-19

 and
and
Abstract
:1. Introduction
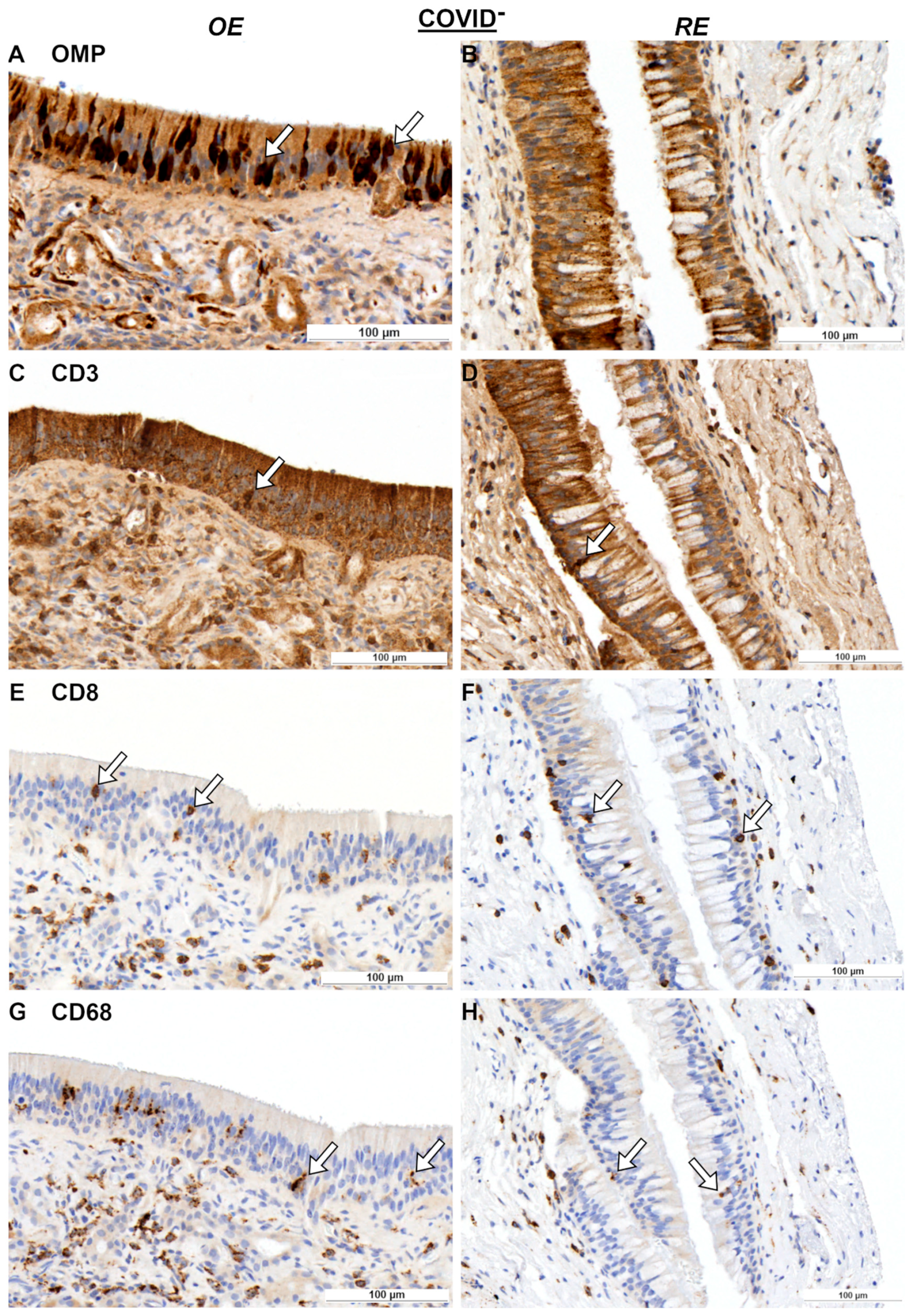
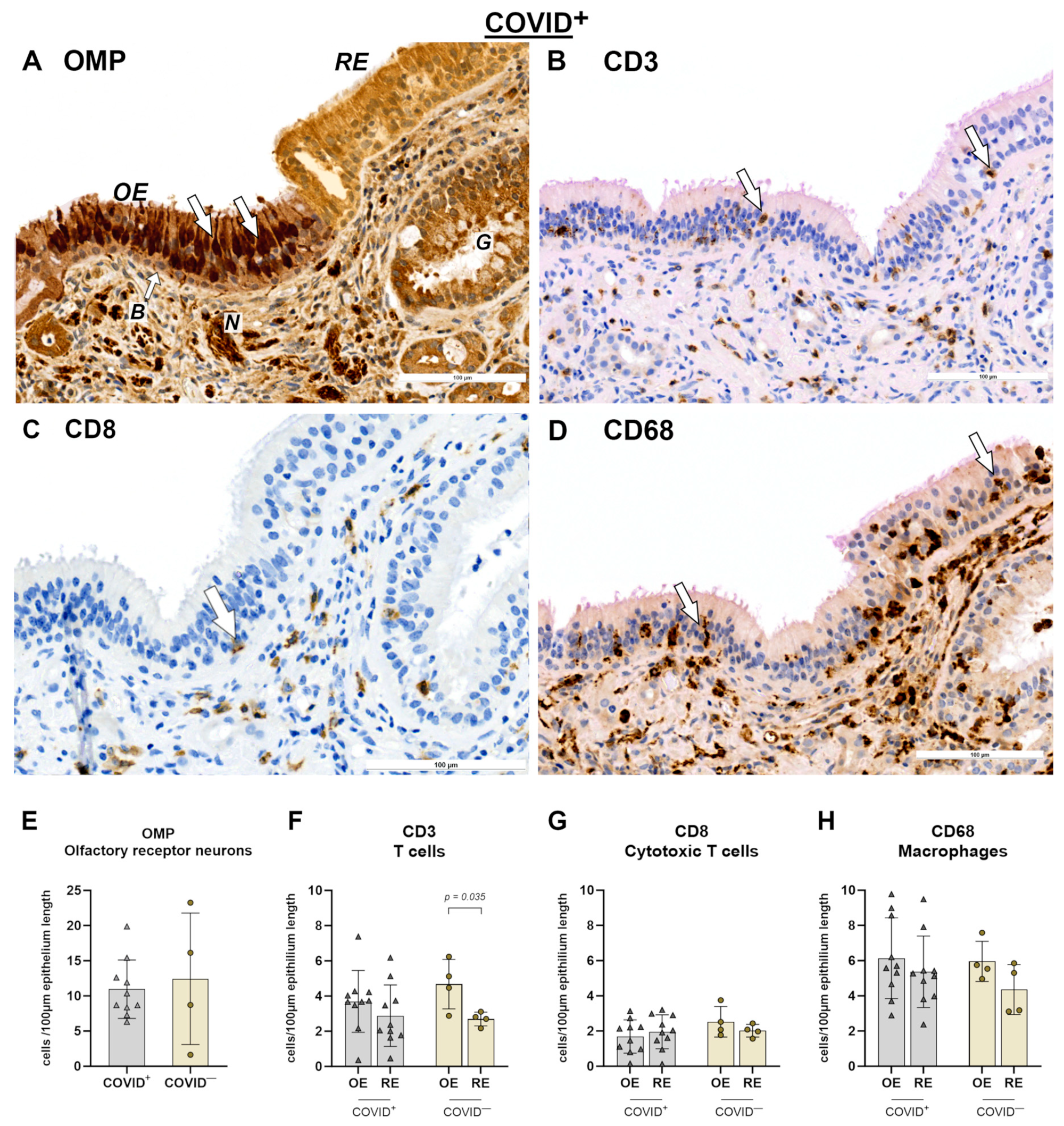
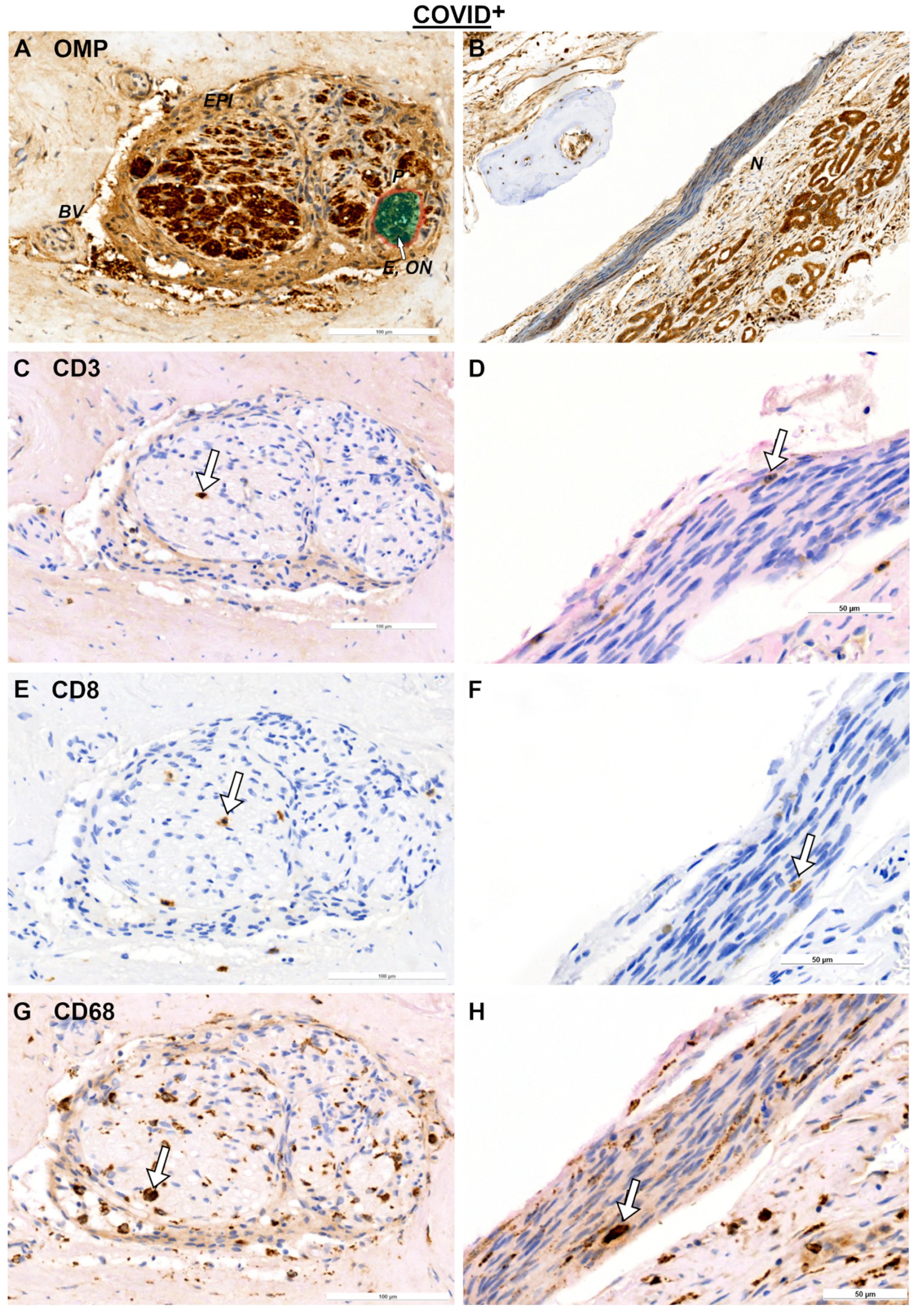
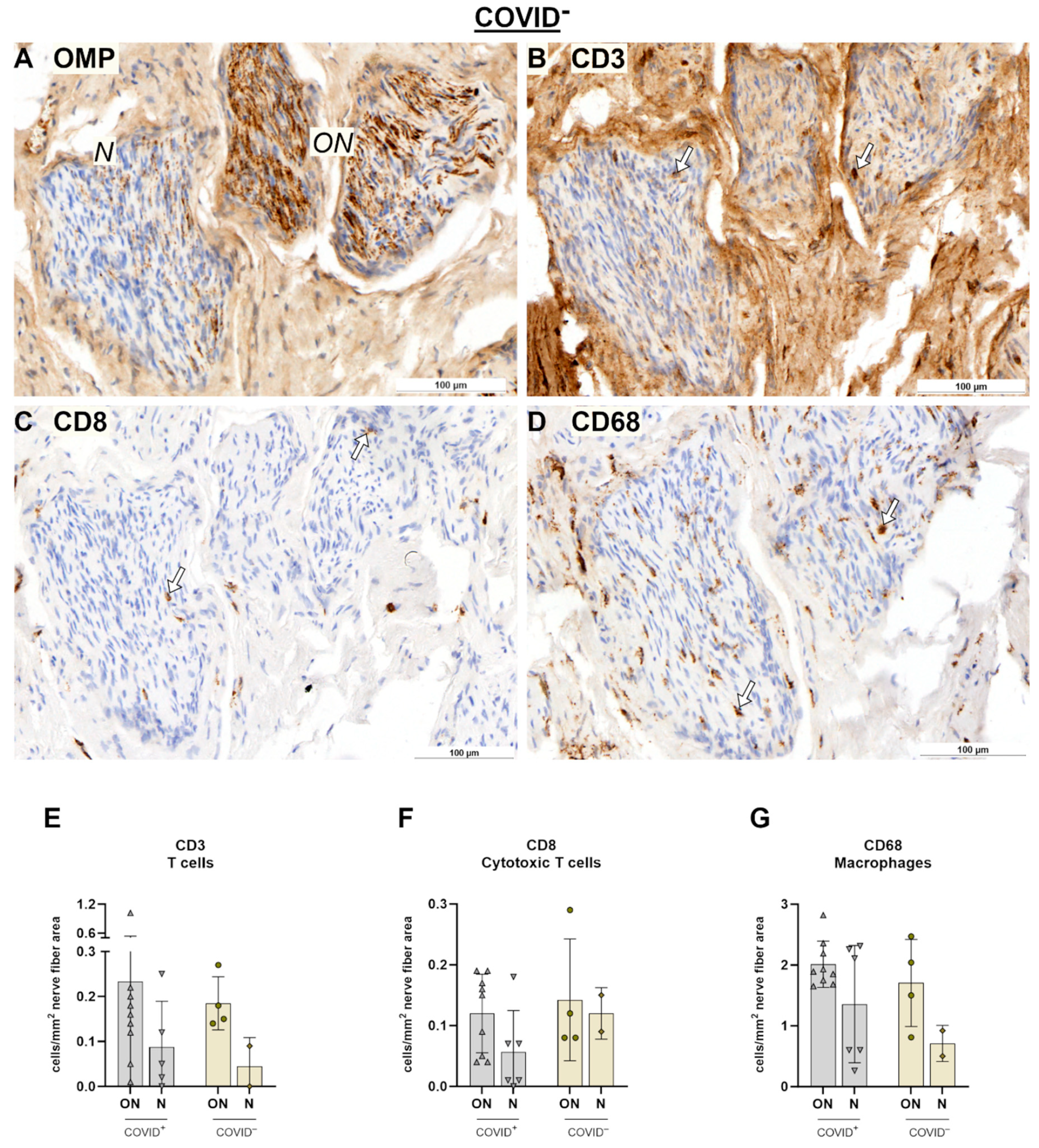
2. Results
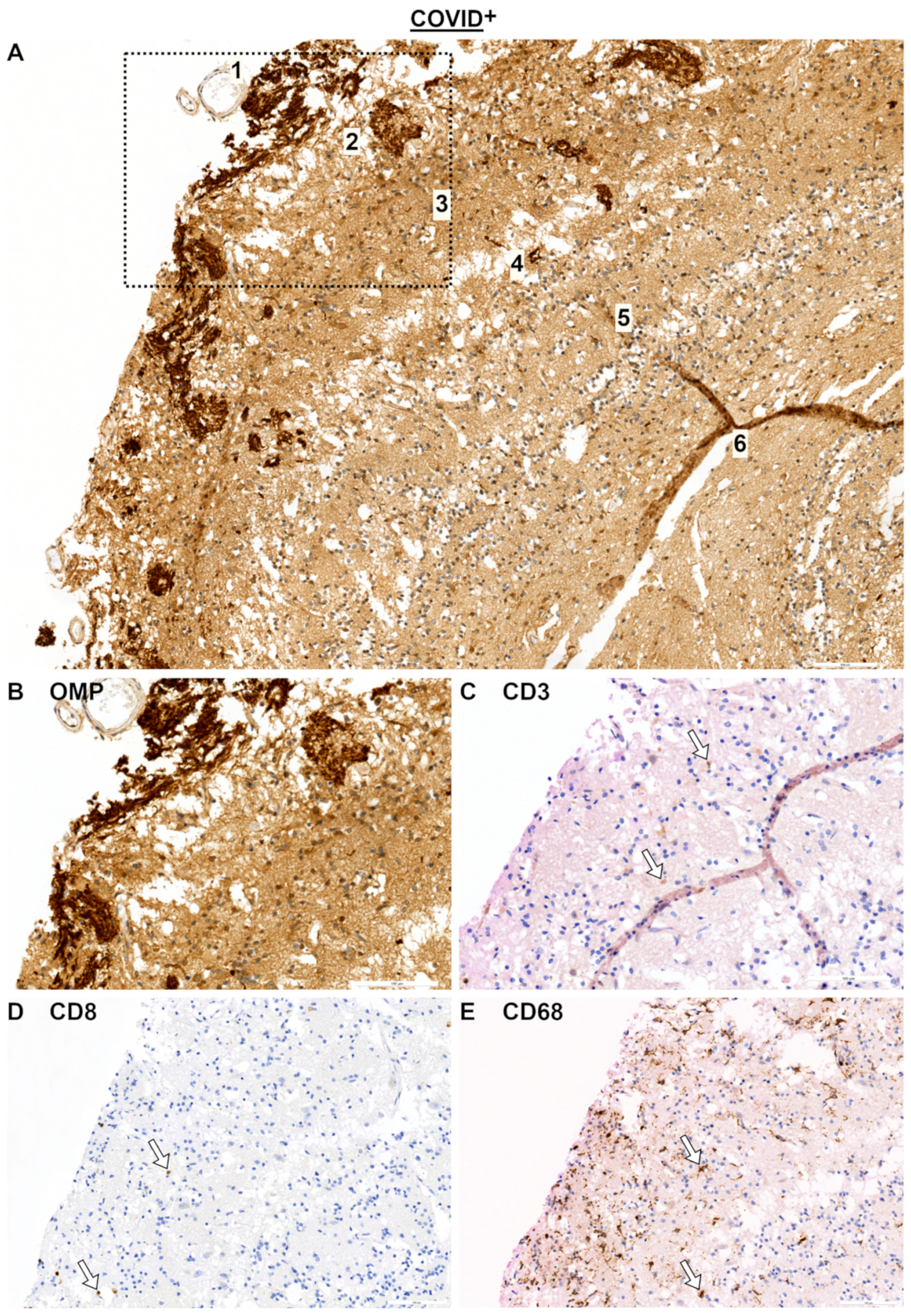
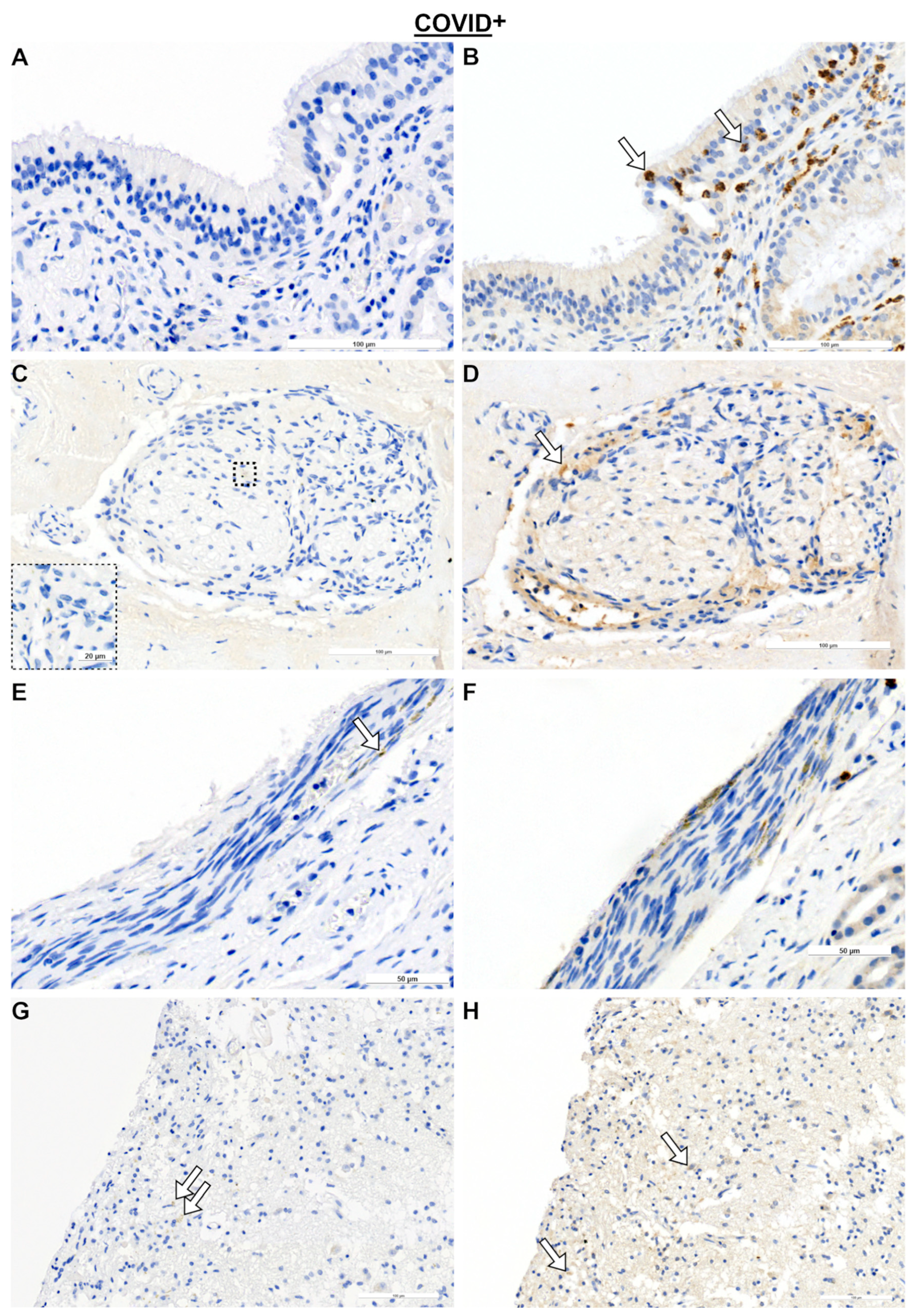
2.1. Olfactory Mucosa and Neural Tissue
2.2. Olfactory Bulb (OB) and SARS Spike Protein
3. Discussion
3.1. Immune Response in Nasal Mucosa
3.2. Immune Responses in Nerve Fibers and Central Nervous System
3.3. Limitations
4. Materials and Methods
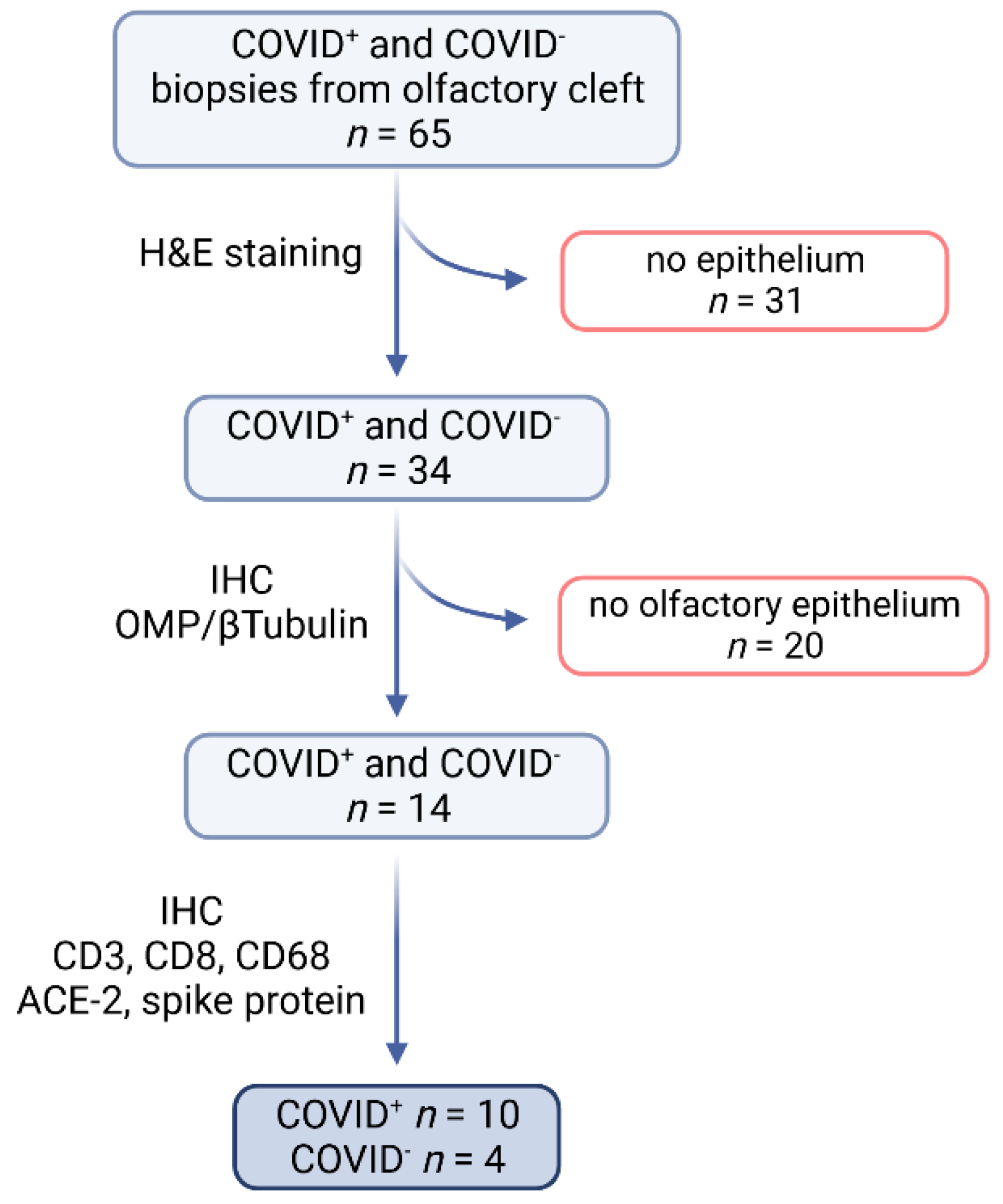
4.1. Study Design and Group
4.2. Sample Processing and Selection
4.3. Immunohistochemistry (IHC)
4.4. Quantification and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The Species Severe Acute Respiratory Syndrome-Related Coronavirus: Classifying 2019-nCoV and Naming It SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qiu, Y.; Wang, J.; Liu, Y.; Wei, Y.; et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: A descriptive study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.-J.; Ni, Z.-Y.; Hu, Y.; Liang, W.-H.; Ou, C.-Q.; He, J.-X.; Liu, L.; Shan, H.; Lei, C.-L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xu, Y.; Gao, R.; Lu, R.; Han, K.; Wu, G.; Tan, W. Detection of SARS-CoV-2 in Different Types of Clinical Specimens. JAMA 2020, 323, 1843–1844. [Google Scholar] [CrossRef]
- Zou, L.; Ruan, F.; Huang, M.; Liang, L.; Huang, H.; Hong, Z.; Yu, J.; Kang, M.; Song, Y.; Xia, J.; et al. SARS-CoV-2 Viral Load in Upper Respiratory Specimens of Infected Patients. N. Engl. J. Med. 2020, 382, 1177–1179. [Google Scholar] [CrossRef] [PubMed]
- Gallo, O.; Locatello, L.G.; Mazzoni, A.; Novelli, L.; Annunziato, F. The central role of the nasal microenvironment in the transmission, modulation, and clinical progression of SARS-CoV-2 infection. Mucosal Immunol. 2021, 14, 305–316. [Google Scholar] [CrossRef]
- Laue, M.; Kauter, A.; Hoffmann, T.; Möller, L.; Michel, J.; Nitsche, A. Morphometry of SARS-CoV and SARS-CoV-2 particles in ultrathin plastic sections of infected Vero cell cultures. Sci. Rep. 2021, 11, 3515. [Google Scholar] [CrossRef]
- Fehr, A.R.; Perlman, S. Coronaviruses: An overview of their replication and pathogenesis. In Coronaviruses; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2015; Volume 1282, pp. 1–23. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; van der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef]
- Sungnak, W.; Huang, N.; Bécavin, C.; Berg, M.; Queen, R.; Litvinukova, M.; Talavera-López, C.; Maatz, H.; Reichart, D.; Sampaziotis, F.; et al. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat. Med. 2020, 26, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, S. The immune mechanism of the nasal epithelium in COVID-19-related olfactory dysfunction. Front. Immunol. 2023, 14, 1045009. [Google Scholar] [CrossRef]
- Khan, M.; Yoo, S.-J.; Clijsters, M.; Backaert, W.; Vanstapel, A.; Speleman, K.; Lietaer, C.; Choi, S.; Hether, T.D.; Marcelis, L.; et al. Visualizing in deceased COVID-19 patients how SARS-CoV-2 attacks the respiratory and olfactory mucosae but spares the olfactory bulb. Cell 2021, 184, 5932–5949.e15. [Google Scholar] [CrossRef] [PubMed]
- Hatton, C.F.; Botting, R.A.; Dueñas, M.E.; Haq, I.J.; Verdon, B.; Thompson, B.J.; Spegarova, J.S.; Gothe, F.; Stephenson, E.; Gardner, A.I.; et al. Delayed induction of type I and III interferons mediates nasal epithelial cell permissiveness to SARS-CoV-2. Nat. Commun. 2021, 12, 7092. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, C.G.K.; Miao, V.N.; Owings, A.H.; Navia, A.W.; Tang, Y.; Bromley, J.D.; Lotfy, P.; Sloan, M.; Laird, H.; Williams, H.B.; et al. Impaired local intrinsic immunity to SARS-CoV-2 infection in severe COVID-19. Cell 2021, 184, 4713–4733.e22. [Google Scholar] [CrossRef]
- Minkoff, J.M.; tenOever, B. Innate immune evasion strategies of SARS-CoV-2. Nat. Rev. Microbiol. 2023, 21, 178–194. [Google Scholar] [CrossRef]
- Zazhytska, M.; Kodra, A.; Hoagland, D.A.; Fullard, J.F.; Shayya, H.; Omer, A.; Firestein, S.; Gong, Q.; Canoll, P.D.; Goldman, J.E.; et al. Disruption of nuclear architecture as a cause of COVID-19 induced anosmia. bioRxiv 2021. [Google Scholar] [CrossRef]
- Verma, A.K.; Zheng, J.; Meyerholz, D.K.; Perlman, S. SARS-CoV-2 infection of sustentacular cells disrupts olfactory signaling pathways. JCI Insight 2022, 7, e160277. [Google Scholar] [CrossRef]
- Bourgon, C.; Albin, A.S.; Ando-Grard, O.; Da Costa, B.; Domain, R.; Korkmaz, B.; Klonjkowski, B.; Le Poder, S.; Meunier, N. Neutrophils play a major role in the destruction of the olfactory epithelium during SARS-CoV-2 infection in hamsters. Cell. Mol. Life Sci. 2022, 79, 616. [Google Scholar] [CrossRef]
- Wellford, S.A.; Moseman, E.A. Olfactory immune response to SARS-CoV-2. Cell. Mol. Immunol. 2024, 21, 134–143. [Google Scholar] [CrossRef]
- Ho, C.-Y.; Salimian, M.; Hegert, J.; O’Brien, J.; Choi, S.G.; Ames, H.; Morris, M.; Papadimitriou, J.C.; Mininni, J.; Niehaus, P.; et al. Postmortem Assessment of Olfactory Tissue Degeneration and Microvasculopathy in Patients with COVID-19. JAMA Neurol. 2022, 79, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Doty, R.L. (Ed.) Handbook of Olfaction and Gustation, 3rd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2015; ISBN 9781118971758. [Google Scholar]
- Witt, M. Anatomy and Development of the Human Gustatory and Olfactory Systems. In The Senses: A Comprehensive Reference, 2nd ed.; Fritzsch, B., Ed.; Elsevier: Oxford, UK, 2020; pp. 85–118. ISBN 978-0-12-805409-3. [Google Scholar]
- World Health Organization. Technical Advisory Group on SARS-CoV-2 Virus Evolution. Available online: https://www.who.int/groups/technical-advisory-group-on-sars-cov-2-virus-evolution (accessed on 5 March 2024).
- Whitaker, M.; Elliott, J.; Bodinier, B.; Barclay, W.; Ward, H.; Cooke, G.; Donnelly, C.A.; Chadeau-Hyam, M.; Elliott, P. Variant-specific symptoms of COVID-19 in a study of 1,542,510 adults in England. Nat. Commun. 2022, 13, 6856. [Google Scholar] [CrossRef] [PubMed]
- De Candia, P.; Prattichizzo, F.; Garavelli, S.; Matarese, G. T Cells: Warriors of SARS-CoV-2 Infection. Trends Immunol. 2021, 42, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Shang, K.; Wang, W.; et al. Dysregulation of Immune Response in Patients With Coronavirus 2019 (COVID-19) in Wuhan, China. Clin. Infect. Dis. 2020, 71, 762–768. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Long, W.; Tu, M.; Chen, S.; Huang, Y.; Wang, S.; Zhou, W.; Chen, D.; Zhou, L.; Wang, M.; et al. Lymphocyte subset (CD4+, CD8+) counts reflect the severity of infection and predict the clinical outcomes in patients with COVID-19. J. Infect. 2020, 81, 318–356. [Google Scholar] [CrossRef]
- Mallajosyula, V.; Ganjavi, C.; Chakraborty, S.; McSween, A.M.; Pavlovitch-Bedzyk, A.J.; Wilhelmy, J.; Nau, A.; Manohar, M.; Nadeau, K.C.; Davis, M.M. CD8+ T cells specific for conserved coronavirus epitopes correlate with milder disease in COVID-19 patients. Sci. Immunol. 2021, 6, eabg5669. [Google Scholar] [CrossRef]
- Gao, F.; Mallajoysula, V.; Arunachalam, P.S.; van der Ploeg, K.; Manohar, M.; Röltgen, K.; Yang, F.; Wirz, O.; Hoh, R.; Haraguchi, E.; et al. Spheromers reveal robust T cell responses to the Pfizer/BioNTech vaccine and attenuated peripheral CD8+ T cell responses post SARS-CoV-2 infection. Immunity 2023, 56, 864–878.e4. [Google Scholar] [CrossRef] [PubMed]
- Bergwerk, M.; Gonen, T.; Lustig, Y.; Amit, S.; Lipsitch, M.; Cohen, C.; Mandelboim, M.; Levin, E.G.; Rubin, C.; Indenbaum, V.; et al. Covid-19 Breakthrough Infections in Vaccinated Health Care Workers. N. Engl. J. Med. 2021, 385, 1474–1484. [Google Scholar] [CrossRef]
- Eyre, D.W.; Taylor, D.; Purver, M.; Chapman, D.; Fowler, T.; Pouwels, K.B.; Walker, A.S.; Peto, T.E.A. Effect of Covid-19 Vaccination on Transmission of Alpha and Delta Variants. N. Engl. J. Med. 2022, 386, 744–756. [Google Scholar] [CrossRef]
- Hall, V.; Foulkes, S.; Insalata, F.; Kirwan, P.; Saei, A.; Atti, A.; Wellington, E.; Khawam, J.; Munro, K.; Cole, M.; et al. Protection against SARS-CoV-2 after Covid-19 Vaccination and Previous Infection. N. Engl. J. Med. 2022, 386, 1207–1220. [Google Scholar] [CrossRef]
- Zhou, D.; Chan, J.F.-W.; Zhou, B.; Zhou, R.; Li, S.; Shan, S.; Liu, L.; Zhang, A.J.; Chen, S.J.; Chan, C.C.-S.; et al. Robust SARS-CoV-2 infection in nasal turbinates after treatment with systemic neutralizing antibodies. Cell Host Microbe 2021, 29, 551–563.e5. [Google Scholar] [CrossRef] [PubMed]
- Wellford, S.A.; Moseman, A.P.; Dao, K.; Wright, K.E.; Chen, A.; Plevin, J.E.; Liao, T.-C.; Mehta, N.; Moseman, E.A. Mucosal plasma cells are required to protect the upper airway and brain from infection. Immunity 2022, 55, 2118–2134.e6. [Google Scholar] [CrossRef] [PubMed]
- Kirschenbaum, D.; Imbach, L.L.; Rushing, E.J.; Frauenknecht, K.B.M.; Gascho, D.; Ineichen, B.V.; Keller, E.; Kohler, S.; Lichtblau, M.; Reimann, R.R.; et al. Intracerebral endotheliitis and microbleeds are neuropathological features of COVID-19. Neuropathol. Appl. Neurobiol. 2021, 47, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Finlay, J.B.; Brann, D.H.; Abi Hachem, R.; Jang, D.W.; Oliva, A.D.; Ko, T.; Gupta, R.; Wellford, S.A.; Moseman, E.A.; Jang, S.S.; et al. Persistent post-COVID-19 smell loss is associated with immune cell infiltration and altered gene expression in olfactory epithelium. Sci. Transl. Med. 2022, 14, eadd0484. [Google Scholar] [CrossRef] [PubMed]
- Roukens, A.H.E.; Pothast, C.R.; König, M.; Huisman, W.; Dalebout, T.; Tak, T.; Azimi, S.; Kruize, Y.; Hagedoorn, R.S.; Zlei, M.; et al. Prolonged activation of nasal immune cell populations and development of tissue-resident SARS-CoV-2-specific CD8+ T cell responses following COVID-19. Nat. Immunol. 2022, 23, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, J.; Radke, J.; Dittmayer, C.; Franz, J.; Thomas, C.; Mothes, R.; Laue, M.; Schneider, J.; Brünink, S.; Greuel, S.; et al. Olfactory transmucosal SARS-CoV-2 invasion as a port of central nervous system entry in individuals with COVID-19. Nat. Neurosci. 2021, 24, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Schwabenland, M.; Salié, H.; Tanevski, J.; Killmer, S.; Lago, M.S.; Schlaak, A.E.; Mayer, L.; Matschke, J.; Püschel, K.; Fitzek, A.; et al. Deep spatial profiling of human COVID-19 brains reveals neuroinflammation with distinct microanatomical microglia-T-cell interactions. Immunity 2021, 54, 1594–1610.e11. [Google Scholar] [CrossRef]
- Matschke, J.; Lütgehetmann, M.; Hagel, C.; Sperhake, J.P.; Schröder, A.S.; Edler, C.; Mushumba, H.; Fitzek, A.; Allweiss, L.; Dandri, M.; et al. Neuropathology of patients with COVID-19 in Germany: A post-mortem case series. Lancet Neurol. 2020, 19, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Bertoletti, A.; Le Bert, N.; Tan, A.T. SARS-CoV-2-specific T cells in the changing landscape of the COVID-19 pandemic. Immunity 2022, 55, 1764–1778. [Google Scholar] [CrossRef]
- Kirschenbaum, D.; Imbach, L.L.; Ulrich, S.; Rushing, E.J.; Keller, E.; Reimann, R.R.; Frauenknecht, K.B.M.; Lichtblau, M.; Witt, M.; Hummel, T.; et al. Inflammatory olfactory neuropathy in two patients with COVID-19. Lancet 2020, 396, 166. [Google Scholar] [CrossRef]
- Meinhardt, J.; Streit, S.; Dittmayer, C.; Manitius, R.V.; Radbruch, H.; Heppner, F.L. The neurobiology of SARS-CoV-2 infection. Nat. Rev. Neurosci. 2024, 25, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Butowt, R.; Meunier, N.; Bryche, B.; von Bartheld, C.S. The olfactory nerve is not a likely route to brain infection in COVID-19: A critical review of data from humans and animal models. Acta Neuropathol. 2021, 141, 809–822. [Google Scholar] [CrossRef]
- Lee, M.-H.; Perl, D.P.; Nair, G.; Li, W.; Maric, D.; Murray, H.; Dodd, S.J.; Koretsky, A.P.; Watts, J.A.; Cheung, V.; et al. Microvascular Injury in the Brains of Patients with COVID-19. N. Engl. J. Med. 2021, 384, 481–483. [Google Scholar] [CrossRef]
- Krasemann, S.; Dittmayer, C.; von Stillfried, S.; Meinhardt, J.; Heinrich, F.; Hartmann, K.; Pfefferle, S.; Thies, E.; von Manitius, R.; Aschman, T.A.D.; et al. Assessing and improving the validity of COVID-19 autopsy studies—A multicentre approach to establish essential standards for immunohistochemical and ultrastructural analyses. EBioMedicine 2022, 83, 104193. [Google Scholar] [CrossRef] [PubMed]
- Varga, Z.; Flammer, A.J.; Steiger, P.; Haberecker, M.; Andermatt, R.; Zinkernagel, A.S.; Mehra, M.R.; Schuepbach, R.A.; Ruschitzka, F.; Moch, H. Endothelial cell infection and endotheliitis in COVID-19. Lancet 2020, 395, 1417–1418. [Google Scholar] [CrossRef] [PubMed]
- Robles, J.P.; Zamora, M.; Adan-Castro, E.; Siqueiros-Marquez, L.; La Martinez de Escalera, G.; Clapp, C. The spike protein of SARS-CoV-2 induces endothelial inflammation through integrin α5β1 and NF-κB signaling. J. Biol. Chem. 2022, 298, 101695. [Google Scholar] [CrossRef]
- Graham, M.S.; Sudre, C.H.; May, A.; Antonelli, M.; Murray, B.; Varsavsky, T.; Kläser, K.; Canas, L.S.; Molteni, E.; Modat, M.; et al. Changes in symptomatology, reinfection, and transmissibility associated with the SARS-CoV-2 variant B.1.1.7: An ecological study. Lancet Public Health 2021, 6, e335–e345. [Google Scholar] [CrossRef]
- Butowt, R.; Bilinska, K.; von Bartheld, C.S. Olfactory dysfunction in COVID-19: New insights into the underlying mechanisms. Trends Neurosci. 2023, 46, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, A.K.; Hummel, T. Intranasal trigeminal function in chronic rhinosinusitis: A review. Expert Rev. Clin. Immunol. 2023, 19, 921–938. [Google Scholar] [CrossRef]
- Otte, M.S.; Bork, M.-L.; Zimmermann, P.H.; Klußmann, J.P.; Lüers, J.-C. Patients with COVID-19-associated olfactory impairment also show impaired trigeminal function. Auris Nasus Larynx 2022, 49, 147–151. [Google Scholar] [CrossRef]
- Frasnelli, J.; Albrecht, J.; Bryant, B.; Lundström, J.N. Perception of specific trigeminal chemosensory agonists. Neuroscience 2011, 189, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Fitzek, M.; Patel, P.K.; Solomon, P.D.; Lin, B.; Hummel, T.; Schwob, J.E.; Holbrook, E.H. Integrated age-related immunohistological changes occur in human olfactory epithelium and olfactory bulb. J. Comp. Neurol. 2022, 530, 2154–2175. [Google Scholar] [CrossRef]
- World Medical Association Declaration of Helsinki. Ethical principles for medical research involving human subjects. Bull. World Health Organ. 2001, 79, 373–374. [Google Scholar]
- Robert Koch Institut. SARS-CoV-2 Varianten in Deutschland: Daten aus der Integrierten Genomischen Surveillance von SARS-CoV-2. Available online: https://public.data.rki.de/t/public/views/IGS_Dashboard/DashboardVOC?%3Aembed=y&%3AisGuestRedirectFromVizportal=y (accessed on 5 January 2024).
- Robert-Koch-Institut. Coronavirus SARS-CoV-2—SARS-CoV-2: Virologische Basisdaten Sowie Virusvarianten im Zeitraum von 2020–2022. Available online: https://www.rki.de/DE/Content/InfAZ/N/Neuartiges_Coronavirus/Virologische_Basisdaten.html?nn=13490888#doc14716546bodyText9 (accessed on 19 January 2024).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COVID+ | Reference Values | COVID− | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | Mean | SD | Max | Min | n | Mean | SD | Max | Min | |||
| Gender | female | 3 | 3 | |||||||||
| male | 7 | 1 | ||||||||||
| Age [years] | 10 | 78.5 | 12.55 | 94 | 58 | 4 | 73.5 | 12.66 | 85 | 56 | ||
| Weight [kg] | 10 | 63.1 | 12.94 | 83 | 47 | 4 | 77.5 | 16.86 | 92 | 54 | ||
| Days between death and autopsy | 10 | 4.8 | 5.33 | 19 | 1 | 4 | 4.25 | 2.5 | 7 | 1 | ||
| Blood parameters | ||||||||||||
| Leukocytes [GPt/L] | 8 | 8.19 | 5.22 | 17.69 | 1.05 | 3.8–9.8 | 2 | 8.61 | 3.87 | 11.35 | 5.87 | |
| Neutrophilic granulocytes [%] | 5 | 77.76 | 11.99 | 91.4 | 66.8 | 36.0–77.0 | 1 | 48.7 | - | - | - | |
| Neutrophilic granulocytes [GPt/L] | 5 | 24.12↑ | 37.96 | 91.4 | 2.82 | 1.80–7.55 | 1 | 5.53 | - | - | - | |
| Lymphocytes [%] | 6 | 10.57↓ | 4.24 | 15.4 | 5.3 | 20.0–49.0 | 1 | 43.7 | - | - | - | |
| Lymphocytes [GPt/L] | 6 | 3.14 | 6.25 | 15.89 | 0.16 | 1.50–4.00 | 1 | 4.96↑ | - | - | - | |
| Monocytes [%] | 6 | 8.83 | 8.09 | 21.8 | 1 | 0.0–9.0 | 1 | 6.3 | - | - | - | |
| Monocytes [GPt/L] | 6 | 0.82 | 0.76 | 2.1 | 0.01 | 0.20–1.00 | 1 | 0.71 | - | - | - | |
| C-reactive protein [mg/L] | 8 | 57.81↑↑ | 74.76 | 211 | 1.6 | <5.0 | 3 | 3.33 | 1.72 | 5.3 | 2.1 | |
| Primary Antibody | Company (No./Clone No.) | Dilution | Secondary Antibody | Company | Buffer |
|---|---|---|---|---|---|
| OMP (rabbit) | Sigma-Aldrich (Darmstadt, Germany) (MFCD09265364) | 1:6000 | Biotinylated gt-anti-rabbit | Vectorlabs (VEC-BA-1000) | TRIS-EDTA |
| Beta-Tubulin (mouse) | BioLegend (Amsterdam, Netherlands) (TUBB3; TUJ1)) | 1:1000 | Biotinylated gt-anti-mouse | Vectorlabs (VEC-BA-9200) | TRIS-EDTA |
| CD3 (rat) | abcam (Cambridge, UK) (CD3-12) | 1:250 | Biotinylated gt- anti-rat | Vectorlabs (VEC-BA-9400) | TRIS-EDTA |
| CD8 (mouse) | abcam (C8/144B) | 1:100 | Biotinylated gt-anti-mouse | Vectorlabs (VEC-BA-9200) | TRIS-EDTA |
| CD68 (mouse) | abcam (KP1) | 1:500 | Biotinylated gt-anti-mouse | Vectorlabs (VEC-BA-9200) | TRIS-EDTA |
| ACE2 (rabbit) | abcam (Anti-ACE2 antibody, ab15348) | 1:4000 | Biotinylated gt-anti-Rb | Vectorlabs (VEC-BA-1000) | Citrate |
| SARS spike glycoprotein (mouse) | abcam (3A2) | 1:100 | Biotinylated gt-anti-mouse | Vectorlabs (VEC-BA-9200) | Citrate |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Power Guerra, N.; Bierkämper, M.; Pablik, J.; Hummel, T.; Witt, M. Histochemical Evidence for Reduced Immune Response in Nasal Mucosa of Patients with COVID-19. Int. J. Mol. Sci. 2024, 25, 4427. https://doi.org/10.3390/ijms25084427
Power Guerra N, Bierkämper M, Pablik J, Hummel T, Witt M. Histochemical Evidence for Reduced Immune Response in Nasal Mucosa of Patients with COVID-19. International Journal of Molecular Sciences. 2024; 25(8):4427. https://doi.org/10.3390/ijms25084427
Chicago/Turabian StylePower Guerra, Nicole, Martin Bierkämper, Jessica Pablik, Thomas Hummel, and Martin Witt. 2024. "Histochemical Evidence for Reduced Immune Response in Nasal Mucosa of Patients with COVID-19" International Journal of Molecular Sciences 25, no. 8: 4427. https://doi.org/10.3390/ijms25084427
APA StylePower Guerra, N., Bierkämper, M., Pablik, J., Hummel, T., & Witt, M. (2024). Histochemical Evidence for Reduced Immune Response in Nasal Mucosa of Patients with COVID-19. International Journal of Molecular Sciences, 25(8), 4427. https://doi.org/10.3390/ijms25084427