
Effect of Poria cocos Terpenes: Verifying Modes of Action Using Molecular Docking, Drug-Induced Transcriptomes, and Diffusion Network Analyses


Abstract
:
1. Introduction
2. Results
2.1. Data on Terpenes Collected from PC
2.2. Molecular Docking Using Terpenes
2.2.1. Over-Representation Analysis (ORA) Using Unique Terpenes of PC
2.2.2. ORA Using Main Dockable Proteomes
2.2.3. GSEA Using Drug-Induced Transcriptomes
2.3. DN Analysis
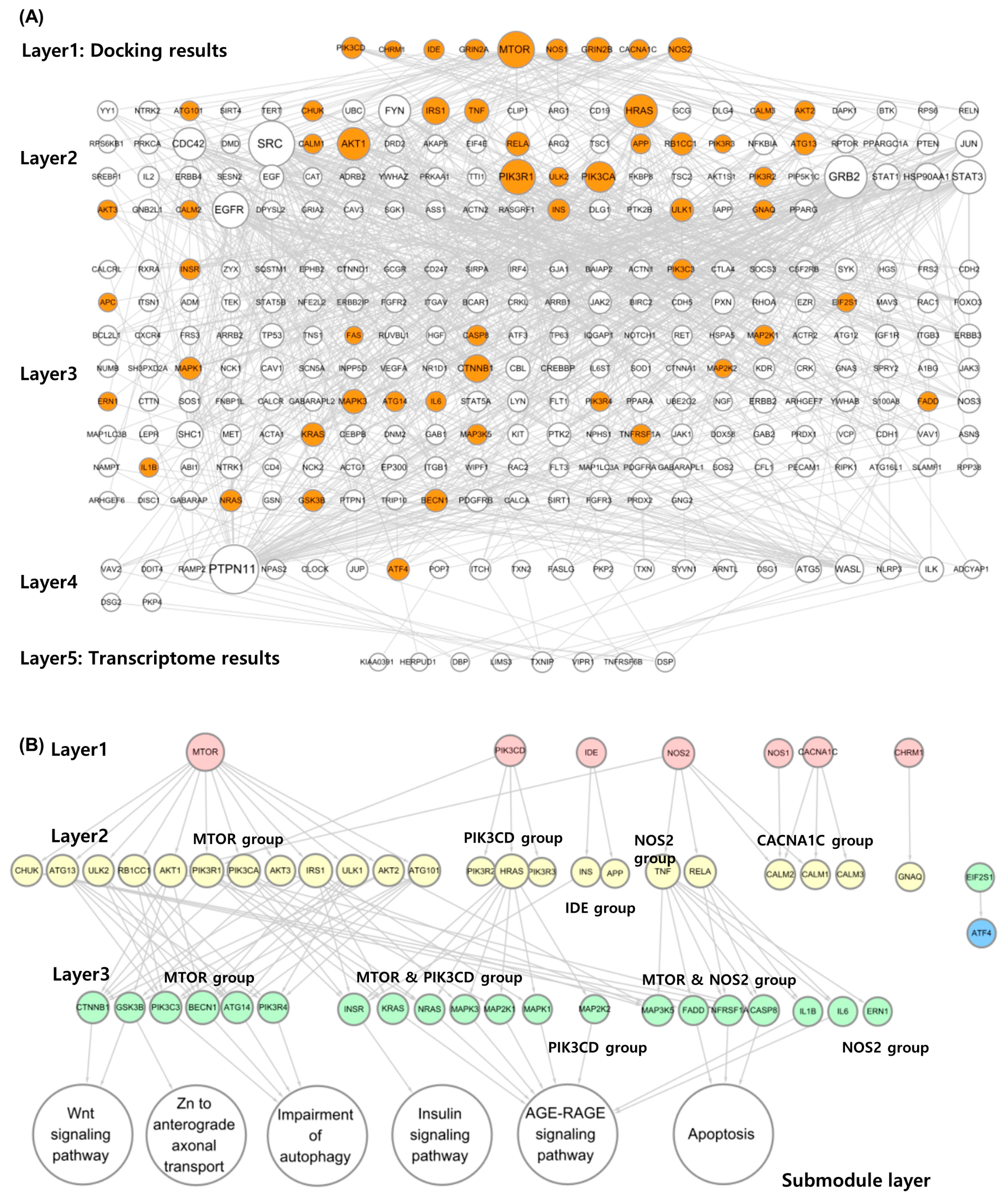
2.3.1. Alzheimer’s Diffusion Network (ADN) Construction
2.3.2. Submodule Construction for Identifying MOA
2.4. Antioxidant Activity of PC in Lipopolysaccharide (LPS)-Stimulated BV2 Microglial Cells
3. Discussion
4. Materials and Methods
4.1. Overview of the Study
4.2. In Silico MDA
4.2.1. Method for Collecting Data on Terpenes Present in PC
4.2.2. Method of Collecting Data Regarding Human-Derived Druggable Proteomes
4.2.3. Large-Scale Molecular Docking Method Using Terpenes and Druggable Proteomes
4.2.4. ORA Using Unique Compounds of PC
4.2.5. ORA Using Main Dockable Proteomes
4.3. Producing PC-Induced Transcriptomes in SW1783 and HT29 Cell Lines
4.3.1. Chemicals and Reagents
4.3.2. Preparation of Hot Water and Ethanol Extracts of PC
4.3.3. Cell Culture
4.3.4. Drug Treatment
4.3.5. RNA Preparation for RNA-Seq
4.3.6. RNA-Seq Preprocessing and Analysis of Differentially Expressed Genes (DEGs)
4.4. GSEA and Cluster Analysis of Pathways
4.5. DN Construction
4.5.1. Method for Construction of AD DN Based on the Disease Pathway
4.5.2. Submodule Construction Method for Identification of MOA
4.6. Determination of Antioxidant Activity of PC
4.6.1. Quantification of Intracellular ROS
4.6.2. Real-Time Polymerase Chain Reaction
4.6.3. Western Blotting
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AF | AlphaFold 2.0 database |
| AND | Alzheimer’s diffusion network |
| ATCC | American Type Culture Collection |
| BH | Benjamini–Hochberg procedure |
| DCF-DA | 2′,7′-Dichlorofluorescein diacetate |
| DEGs | Differentially expressed genes |
| DMEM | Dulbecco’s modified Eagle medium |
| DN | Diffusion network |
| DTA | Drug-induced transcriptome analysis |
| EEPC | 70% ethanol extract of Poria cocos F.A. Wolf |
| FBS | Fetal bovine serum |
| GABA | Gamma-aminobutyric acid |
| GO | Gene ontology |
| GSEA | Gene set enrichment analysis |
| HBSS | Hanks’ balanced salt solution |
| HPA | Human Protein Atlas |
| KAGS | KEGG Alzheimer’s pathway gene set |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LPS | Lipopolysaccharide |
| MDA | Molecular docking analysis |
| MOA | Modes of action |
| NAC | N-acetyl-L-cystein |
| OMIM | Online Mendelian Inheritance in Man disease |
| ORA | Over-representation analysis |
| PBS | Phosphate-buffered saline |
| PC | Poria cocos F.A. Wolf |
| PPI | Protein–protein interaction |
| qPCR | Real-time polymerase chain reaction |
| ROS | Reactive oxygen species |
| TCMID | Traditional Chinese Medicine Integrated Database |
| TCMSP | Traditional Chinese Medicine Systems Pharmacology Database and Analysis Platform |
| WEPC | water extract of Poria cocos F.A. Wolf |
References
- Chang, R. Functional properties of edible mushrooms. Nutr. Rev. 1996, 54, S91–S93. [Google Scholar] [CrossRef]
- Breitmaier, E. Terpenes: Flavors, Fragrances, Pharmaca, Pheromones; John Wiley & Sons: Hoboken, NJ, USA, 2006. [Google Scholar]
- Wang, W.H.; Dong, H.J.; Yan, R.Y.; Li, H.; Li, P.Y.; Chen, P.; Yang, B.; Wang, Z.M. Comparative study of lanostane-type triterpene acids in different parts of Poria cocos (Schw.) Wolf by UHPLC-Fourier transform MS and UHPLC-triple quadruple MS. J. Pharm. Biomed. 2015, 102, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Nukaya, H.; Yamashiro, H.; Fukazawa, H.; Ishida, H.; Tsuji, K. Isolation of inhibitors of TPA-induced mouse ear edema from hoelen, Poria cocos. Chem. Pharm. Bull. 1996, 44, 847–849. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.-J.; Kang, K.-W.; Kang, J.-Y.; Yoon, J.-H.; Choi, Y.-M.; Kim, H.-J.; Sun, S.-H.; Jang, I.-S. An Overview of the Applicability of Oryung-san as an Antihypertensive Agent. J. Intern. Korean Med. 2017, 38, 443–454. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, J.I.; Baeg, M.K.; Sunwoo, Y.Y.; Do, K.; Lee, J.H.; Kim, H.J.; Choi, J.S.; Kim, J.; Seo, C.S.; et al. Effect of Samryungbaekchul-san Combined with Otilonium Bromide on Diarrhea-Predominant Irritable Bowel Syndrome: A Pilot Randomized Controlled Trial. J. Clin. Med. 2019, 8, 1558. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.J.; Park, J.W.; Lee, J.H.; Cho, S.H.; Lee, J.; Nam, S.; Kim, J. Herbal medicine Yukgunja-tang for functional dyspepsia protocol for a systematic review of randomized controlled trials. Medicine 2018, 97, e12555. [Google Scholar] [CrossRef]
- Li, H.; Li, Q.; Yang, X.; Wang, Y.; Tian, X.; Chen, X.; Li, Y. Clinical study on Gui Pi Tang treating the depression and improving the life quality in elderly patients. China J. Tradit. Chin. Med. Pharm. 2014, 29, 1855–1859. [Google Scholar]
- Rios, J.L. Chemical Constituents and Pharmacological Properties of Poria cocos. Planta Med. 2011, 77, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Bemis, G.W.; Murcko, M.A. The properties of known drugs. 1. Molecular frameworks. J. Med. Chem. 1996, 39, 2887–2893. [Google Scholar] [CrossRef]
- Hu, Y.; Stumpfe, D.; Bajorath, J. Computational Exploration of Molecular Scaffolds in Medicinal Chemistry. J. Med. Chem. 2016, 59, 4062–4076. [Google Scholar] [CrossRef]
- Park, M.; Baek, S.-J.; Park, S.-M.; Yi, J.-M.; Cha, S. Comparative study of the mechanism of natural compounds with similar structures using docking and transcriptome data for improving in silico herbal medicine experimentations. bioRxiv 2023, bioRxiv:2023.04.23.538005. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Zhang, J.; Zhao, Y.L.; Li, T.; Shen, T.; Li, J.Q.; Li, W.Y.; Liu, H.G. Mycology, cultivation, traditional uses, phytochemistry and pharmacology of Wolfiporia cocos (Schwein.) Ryvarden et Gilb.: A review. J. Ethnopharmacol. 2013, 147, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Tian, J.; Zhou, L.; Meng, L.J.; Chen, S.T.; Ma, C.Y.; Wang, J.M.; Liu, Z.H.; Li, C.Q.; Kang, W.Y. Phytochemistry and Biological Activities of Poria. J. Chem. 2021, 2021, 6659775. [Google Scholar] [CrossRef]
- Nie, A.; Chao, Y.; Zhang, X.; Jia, W.; Zhou, Z.; Zhu, C. Phytochemistry and Pharmacological Activities of Wolfiporia cocos (F.A. Wolf) Ryvarden & Gilb. Front. Pharmacol. 2020, 11, 505249. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Fu, A.; Zhang, L. Progress in molecular docking. Quant. Biol. 2019, 7, 83–89. [Google Scholar] [CrossRef]
- Dobin, A.; Gingeras, T.R. Mapping RNA-seq Reads with STAR. Curr. Protoc. Bioinform. 2015, 51, 11.14.1–11.14.19. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, Y.H.; Zhang, Z.; Huang, T.; Cai, Y.D. Inferring Novel Tumor Suppressor Genes with a Protein-Protein Interaction Network and Network Diffusion Algorithms. Mol. Ther. Methods Clin. Dev. 2018, 10, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Cobourne-Duval, M.K.; Taka, E.; Mendonca, P.; Bauer, D.; Soliman, K.F.A. The Antioxidant Effects of Thymoquinone in Activated BV-2 Murine Microglial Cells. Neurochem. Res. 2016, 41, 3227–3238. [Google Scholar] [CrossRef] [PubMed]
- Del Prado-Audelo, M.L.; Cortes, H.; Caballero-Floran, I.H.; Gonzalez-Torres, M.; Escutia-Guadarrama, L.; Bernal-Chavez, S.A.; Giraldo-Gomez, D.M.; Magana, J.J.; Leyva-Gomez, G. Therapeutic Applications of Terpenes on Inflammatory Diseases. Front. Pharmacol. 2021, 12, 704197. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.J.; Antonetti, D.A. The role of small GTPases and EPAC-Rap signaling in the regulation of the blood-brain and blood-retinal barriers. Tissue Barriers 2017, 5, e1339768. [Google Scholar] [CrossRef]
- Baulch, J.E.; Acharya, M.M.; Agrawal, S.; Apodaca, L.A.; Monteiro, C.; Agrawal, A. Immune and Inflammatory Determinants Underlying Alzheimer’s Disease Pathology. J. Neuroimmune Pharmacol. 2020, 15, 852–862. [Google Scholar] [CrossRef]
- Stokman, G.; Qin, Y.; Booij, T.H.; Ramaiahgari, S.; Lacombe, M.; Dolman, M.E.M.; van Dorenmalen, K.M.A.; Teske, G.J.D.; Florquin, S.; Schwede, F.; et al. Epac-Rap Signaling Reduces Oxidative Stress in the Tubular Epithelium. J. Am. Soc. Nephrol. 2014, 25, 1474–1485. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.K.; Kirton, H.M.; Al-Owais, M.; Thireau, J.; Richard, S.; Peers, C.; Steele, D.S. Epac2-Rap1 Signaling Regulates Reactive Oxygen Species Production and Susceptibility to Cardiac Arrhythmias. Antioxid. Redox Signal. 2017, 27, 117–132. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zheng, Y.L.; Bian, X.W.; Wang, M.H.; Chou, J.S.; Liu, H.F.; Wang, Z.M. Identification of Key Genes and Pathways Associated with Oxidative Stress in Periodontitis. Oxidative Med. Cell. Longev. 2022, 2022, 9728172. [Google Scholar] [CrossRef] [PubMed]
- Raghavendra, S.; Rao, S.J.A.; Kumar, V.; Ramesh, C.K. Multiple ligand simultaneous docking (MLSD): A novel approach to study the effect of inhibitors on substrate binding to PPO. Comput. Biol. Chem. 2015, 59, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Li, T.H.; Hou, C.C.; Chang, C.L.; Yang, W.C. Anti-hyperglycemic properties of crude extract and triterpenes from Poria cocos. Evid. Based Complement. Altern. Med. 2011, 2011, 128402. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.L.; Huang, H.W.; Su, M.H.; Lin, H.C.; Wu, W.M. The Lanostane Triterpenoids in Poria cocos Play Beneficial Roles in Immunoregulatory Activity. Life 2021, 11, 111. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Yang, Y.; Zhang, F.; Li, Y.; Li, X.; Pu, X.; He, X.; Zhang, M.; Yang, X.; Yu, Q.; et al. Effects of Poria cocos extract on metabolic dysfunction-associated fatty liver disease via the FXR/PPARalpha-SREBPs pathway. Front. Pharmacol. 2022, 13, 1007274. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.F.; Liu, Z.Q.; Pi, Z.F.; Song, F.R.; Wu, J.L.; Liu, S. Poria cocos could ameliorate cognitive dysfunction in APP/PS1 mice by restoring imbalance of A beta production and clearance and gut microbiota dysbiosis. Phytother. Res. 2021, 35, 2678–2690. [Google Scholar] [CrossRef] [PubMed]
- Na, S.-S.; Chong, M.-S.; Woo, J.-H.; Kwon, Y.-O.; Lee, M.K.; Oh, K.-W. Poria cocos ethanol extract and its active constituent, pachymic acid, modulate sleep architectures via activation of GABAA-ergic transmission in rats. J. Biomed. Transl. Res. 2015, 16, 84–92. [Google Scholar] [CrossRef]
- Jia, X.J.; Ma, L.S.; Li, P.; Chen, M.W.; He, C.W. Prospects of Poria cocos polysaccharides: Isolation process, structural features and bioactivities. Trends Food Sci. Technol. 2016, 54, 52–62. [Google Scholar] [CrossRef]
- Ru, J.L.; Li, P.; Wang, J.N.; Zhou, W.; Li, B.H.; Huang, C.; Li, P.D.; Guo, Z.H.; Tao, W.Y.; Yang, Y.F.; et al. TCMSP: A database of systems pharmacology for drug discovery from herbal medicines. J. Cheminform. 2014, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Xie, D.; Yu, Y.; Liu, H.; Shi, Y.; Shi, T.; Wen, C. TCMID 2.0: A comprehensive resource for TCM. Nucleic Acids Res. 2018, 46, D1117–D1120. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Nam, S.; Jang, H.; Kim, A.; Lee, J.J. TM-MC: A database of medicinal materials and chemical compounds in Northeast Asian traditional medicine. BMC Complement. Altern. Med. 2015, 15, 218. [Google Scholar] [CrossRef] [PubMed]
- Degtyarenko, K.; de Matos, P.; Ennis, M.; Hastings, J.; Zbinden, M.; McNaught, A.; Alcantara, R.; Darsow, M.; Guedj, M.; Ashburner, M. ChEBI: A database and ontology for chemical entities of biological interest. Nucleic Acids Res. 2008, 36, D344–D350. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem Substance and Compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Bjorling, E.; Agaton, C.; Szigyarto, C.A.; Amini, B.; Andersen, E.; Andersson, A.C.; Angelidou, P.; Asplund, A.; Asplund, C.; et al. A human protein atlas for normal and cancer tissues based on antibody proteomics. Mol. Cell. Proteom. 2005, 4, 1920–1932. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Hamosh, A.; Scott, A.F.; Amberger, J.S.; Bocchini, C.A.; McKusick, V.A. Online Mendelian Inheritance in Man (OMIM), a knowledgebase of human genes and genetic disorders. Nucleic Acids Res. 2005, 33, D514–D517. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Liu, X.; Peltz, G. GSEApy: A comprehensive package for performing gene set enrichment analysis in Python. Bioinformatics 2023, 39, btac757. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Welch, W.J. Construction of permutation tests. J. Am. Stat. Assoc. 1990, 85, 693–698. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Liberzon, A.; Subramanian, A.; Pinchback, R.; Thorvaldsdottir, H.; Tamayo, P.; Mesirov, J.P. Molecular signatures database (MSigDB) 3.0. Bioinformatics 2011, 27, 1739–1740. [Google Scholar] [CrossRef]
- Korotkevich, G.; Sukhov, V.; Budin, N.; Shpak, B.; Artyomov, M.N.; Sergushichev, A. Fast gene set enrichment analysis. bioRxiv 2016, bioRxiv:060012. [Google Scholar] [CrossRef]
- Murtagh, F.; Contreras, P. Algorithms for hierarchical clustering: An overview. Wires Data Min. Knowl. 2012, 2, 86–97. [Google Scholar] [CrossRef]
- von Mering, C.; Huynen, M.; Jaeggi, D.; Schmidt, S.; Bork, P.; Snel, B. STRING: A database of predicted functional associations between proteins. Nucleic Acids Res. 2003, 31, 258–261. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Lanostane-Type Triterpenes | Seco-Lanostane-Type Triterpenes | Non-Lanostane-Type |
|---|---|---|---|
| ITGAL | 0.910714 | 0.774194 | 0.263158 |
| ITGAX | 0.821429 | 0.806452 | 0.131579 |
| GRIN2B | 0.821429 | 0.903226 | 0.368421 |
| CHRM4 | 0.803571 | 0.967742 | 0.394737 |
| HCRTR1 | 0.785714 | 1 | 0.210526 |
| SLC6A8 | 0.785714 | 0.903226 | 0.394737 |
| SLC6A4 | 0.767857 | 0.709677 | 0.184211 |
| GRIN2A | 0.696429 | 0.967742 | 0.210526 |
| HCRTR2 | 0.678571 | 0.967742 | 0.263158 |
| PRKCZ | 0.678571 | 0.741935 | 0.236842 |
| PTGIS | 0.678571 | 0.645161 | 0.184211 |
| PRKCI | 0.660714 | 0.83871 | 0.315789 |
| DHFR | 0.642857 | 1 | 0.473684 |
| JAK3 | 0.642857 | 0.645161 | 0.394737 |
| JAK1 | 0.642857 | 0.935484 | 0.394737 |
| ACE | 0.625 | 0.677419 | 0.315789 |
| NOS3 | 0.607143 | 0.741935 | 0.447368 |
| SLC22A6 | 0.607143 | 0.709677 | 0.289474 |
| ADORA1 | 0.607143 | 0.935484 | 0.315789 |
| TFRC | 0.517857 | 0.870968 | 0.236842 |
| KEGG Pathway | Docking Count | p-Value |
|---|---|---|
| Calcium signaling pathway | 125 | 0.006 |
| cAMP signaling pathway | 123 | <0.001 |
| Cholinergic synapse | 115 | <0.001 |
| MAPK signaling pathway | 115 | <0.001 |
| Serotonergic synapse | 115 | <0.001 |
| Type II diabetes mellitus | 114 | <0.001 |
| Hypertrophic cardiomyopathy | 110 | <0.001 |
| Oxytocin signaling pathway | 107 | <0.001 |
| GnRH secretion | 102 | <0.001 |
| cGMP-PKG signaling pathway | 101 | <0.001 |
| GABAergic synapse | 100 | <0.001 |
| Adrenergic signaling in cardiomyocytes | 99 | <0.001 |
| Circadian entrainment | 98 | <0.001 |
| Arrhythmogenic right ventricular cardiomyopathy | 92 | <0.001 |
| Dilated cardiomyopathy | 91 | <0.001 |
| Cortisol synthesis and secretion | 90 | <0.001 |
| Aldosterone synthesis and secretion | 88 | <0.001 |
| Prion disease | 87 | <0.001 |
| PI3K-Akt signaling pathway | 84 | <0.001 |
| Pathways of neurodegeneration | 80 | <0.001 |
| Dopaminergic synapse | 79 | <0.001 |
| Renin secretion | 75 | <0.001 |
| Chemical carcinogenesis | 74 | <0.001 |
| Alzheimer’s disease | 73 | <0.001 |
| Cushing’s syndrome | 70 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, M.; Yi, J.-M.; Kim, N.S.; Lee, S.-Y.; Lee, H. Effect of Poria cocos Terpenes: Verifying Modes of Action Using Molecular Docking, Drug-Induced Transcriptomes, and Diffusion Network Analyses. Int. J. Mol. Sci. 2024, 25, 4636. https://doi.org/10.3390/ijms25094636
Park M, Yi J-M, Kim NS, Lee S-Y, Lee H. Effect of Poria cocos Terpenes: Verifying Modes of Action Using Molecular Docking, Drug-Induced Transcriptomes, and Diffusion Network Analyses. International Journal of Molecular Sciences. 2024; 25(9):4636. https://doi.org/10.3390/ijms25094636
Chicago/Turabian StylePark, Musun, Jin-Mu Yi, No Soo Kim, Seo-Young Lee, and Haeseung Lee. 2024. "Effect of Poria cocos Terpenes: Verifying Modes of Action Using Molecular Docking, Drug-Induced Transcriptomes, and Diffusion Network Analyses" International Journal of Molecular Sciences 25, no. 9: 4636. https://doi.org/10.3390/ijms25094636
APA StylePark, M., Yi, J. -M., Kim, N. S., Lee, S. -Y., & Lee, H. (2024). Effect of Poria cocos Terpenes: Verifying Modes of Action Using Molecular Docking, Drug-Induced Transcriptomes, and Diffusion Network Analyses. International Journal of Molecular Sciences, 25(9), 4636. https://doi.org/10.3390/ijms25094636