
Anti-Nociceptive Effects of Sphingomyelinase and Methyl-Beta-Cyclodextrin in the Icilin-Induced Mouse Pain Model

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Cholesterol Depletion Decreases Cell Viability in a Concentration-Dependent Manner
2.2. SMase and MCD Reduced the Duration of Icilin-Induced Nocifensive Behavior
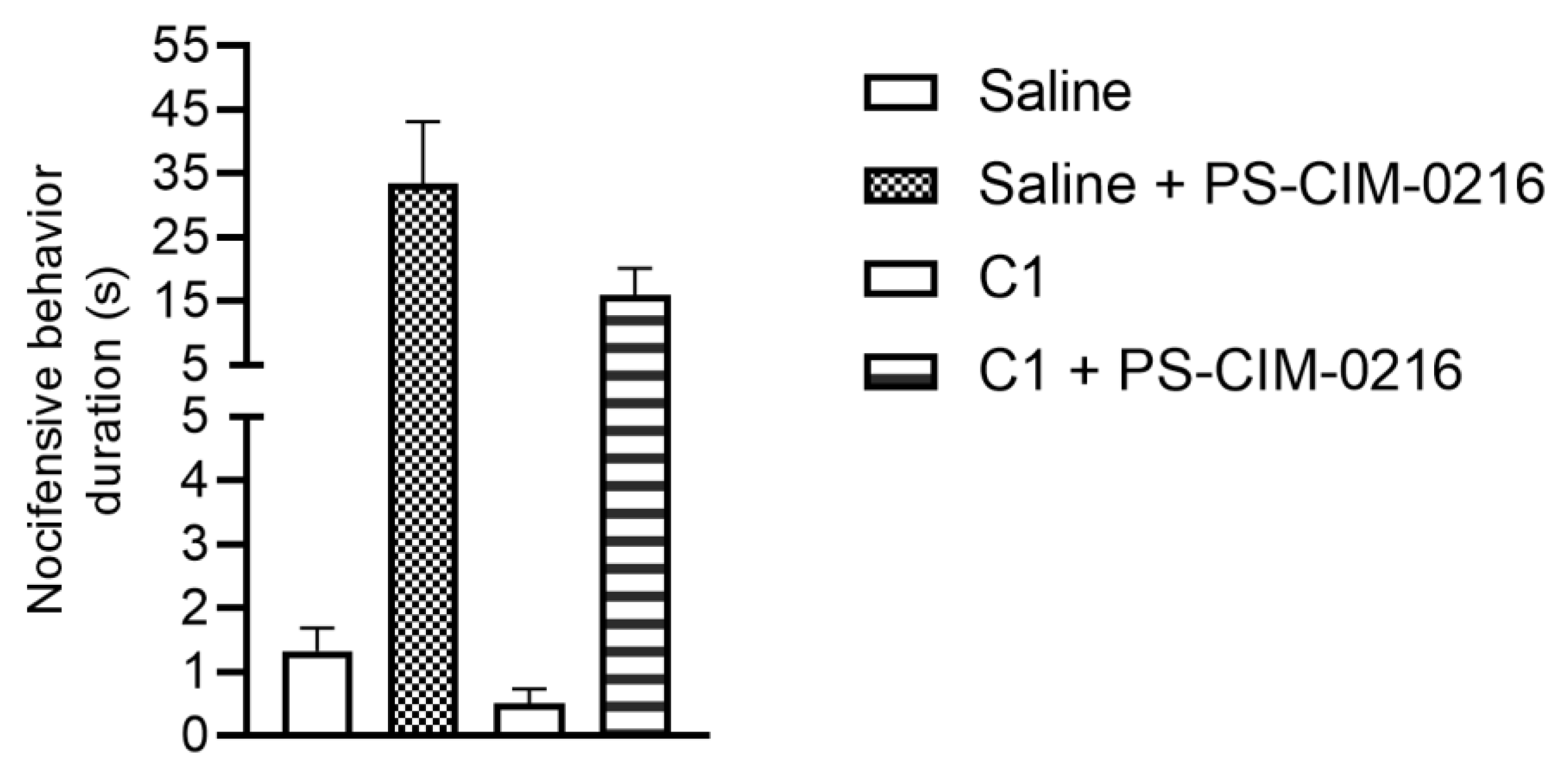
2.3. Effect of Carboxamido-Steroid Compound C1 on Acute Nocifensive Pain Reaction Induced by PS and CIM-0216
2.4. Lipid Raft Disruptors Do Not Inhibit the Activity of Recombinant TRPM3 and TRPM8
3. Discussion
4. Materials and Methods
4.1. Animals and Ethics
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Icilin-Induced Acute Nocifensive Pain Reaction
4.5. PS-CIM-0216 Evoked Acute Nocifensive Pain Reaction
4.6. Fluorescent Ca2+ Measurements
4.7. Drugs and Chemicals
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMPA | 2-amino-3-(5-methyl-3-oxo-1,2-oxazol-4-yl)-propanoic-acid |
| ANOVA | Analysis of Variance |
| ATP | adenosine triphosphate |
| CHO | Chinese Hamster Ovary |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| DMSO | Dimethyl sulfoxide |
| EC50 | Efficacy |
| GABA | Gamma-aminobutyric acid |
| HEK293T | Human Embryonic Kidney 293T |
| hTRPM8 | human Transient Receptor Potential Melastatin 8 |
| [Ca2+]IC | Intracellular Ca2+ |
| MCD | Methyl-beta-cyclodextrin |
| mTRPM3α2 | TRPM3α2 variant of mouse Transient Receptor Potential Melastatin 3 |
| Myr | Myriocin |
| PS | Pregnenolone sulphate |
| SEM | standard error of mean |
| SM | Sphingomyelins |
| SMase | Sphingomyelinase |
| TG | Trigeminal ganglion |
| TRP | Transient Receptor Potential |
| TRPA1 | Transient Receptor Potential Ankryrin 1 |
| TRPM | Transient Receptor Potential Melastatin |
| TRPV1 | Transient Receptor Potential Vanilloid 1 |
References
- Karnovsky, M.J.; Kleinfeld, A.M.; Hoover, R.L.; Klausner, R.D. The Concept of Lipid Domains in Membranes. J. Cell Biol. 1982, 94, 1–6. [Google Scholar] [CrossRef]
- Simons, K.; Ikonen, E. Functional Rafts in Cell Membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Parton, R.G.; Richards, A.A. Lipid Rafts and Caveolae as Portals for Endocytosis: New Insights and Common Mechanisms: Caveolae/Lipid Raft-Dependent Endocytosis. Traffic 2003, 4, 724–738. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Takahashi, M.; Nagatsuka, Y.; Hirabayashi, Y. Lipid Rafts: New Tools and a New Component. Biol. Pharm. Bull. 2006, 29, 1526–1531. [Google Scholar] [CrossRef]
- Kannan, K.B.; Barlos, D.; Hauser, C.J. Free Cholesterol Alters Lipid Raft Structure and Function Regulating Neutrophil Ca2+ Entry and Respiratory Burst: Correlations with Calcium Channel Raft Trafficking. J. Immunol. 2007, 178, 5253–5261. [Google Scholar] [CrossRef]
- Staubach, S.; Razawi, H.; Hanisch, F.-G. Proteomics of MUC1-Containing Lipid Rafts from Plasma Membranes and Exosomes of Human Breast Carcinoma Cells MCF-7. Proteomics 2009, 9, 2820–2835. [Google Scholar] [CrossRef] [PubMed]
- Sezgin, E.; Levental, I.; Mayor, S.; Eggeling, C. The Mystery of Membrane Organization: Composition, Regulation and Roles of Lipid Rafts. Nat. Rev. Mol. Cell Biol. 2017, 18, 361–374. [Google Scholar] [CrossRef]
- Hering, H.; Lin, C.-C.; Sheng, M. Lipid Rafts in the Maintenance of Synapses, Dendritic Spines, and Surface AMPA Receptor Stability. J. Neurosci. 2003, 23, 3262–3271. [Google Scholar] [CrossRef]
- Zhu, D. Lipid Rafts Serve as a Signaling Platform for Nicotinic Acetylcholine Receptor Clustering. J. Neurosci. 2006, 26, 4841–4851. [Google Scholar] [CrossRef]
- Liu, M.; Huang, W.; Wu, D.; Priestley, J.V. TRPV1, but Not P2X3, Requires Cholesterol for Its Function and Membrane Expression in Rat Nociceptors. Eur. J. Neurosci. 2006, 24, 1–6. [Google Scholar] [CrossRef]
- Bobkov, D.; Semenova, S. Impact of Lipid Rafts on Transient Receptor Potential Channel Activities. J. Cell. Physiol. 2022, 237, 2034–2044. [Google Scholar] [CrossRef] [PubMed]
- Ilangumaran, S.; Hoessli, D.C. Effects of Cholesterol Depletion by Cyclodextrin on the Sphingolipid Microdomains of the Plasma Membrane. Biochem. J. 1998, 335, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Szánti-Pintér, E.; Balogh, J.; Csók, Z.; Kollár, L.; Gömöry, Á.; Skoda-Földes, R. Synthesis of Steroid–Ferrocene Conjugates of Steroidal 17-Carboxamides via a Palladium-Catalyzed Aminocarbonylation—Copper-Catalyzed Azide–Alkyne Cycloaddition Reaction Sequence. Steroids 2011, 76, 1377–1382. [Google Scholar] [CrossRef] [PubMed]
- Szánti-Pintér, E.; Wouters, J.; Gömöry, Á.; Sághy, É.; Szőke, É.; Helyes, Z.; Kollár, L.; Skoda-Földes, R. Synthesis of Novel 13α-18-Norandrostane–Ferrocene Conjugates via Homogeneous Catalytic Methods and Their Investigation on TRPV1 Receptor Activation. Steroids 2015, 104, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Kozutsumi, Y.; Nakamura, S.; Fujita, T.; Kawasaki, T. Serine Palmitoyltransferase Is the Primary Target of a Sphingosine-like Immunosuppressant, ISP-1/Myriocin. Biochem. Biophys. Res. Commun. 1995, 211, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Chao, L.; Chen, F.; Jensen, K.F.; Hatton, T.A. Two-Dimensional Solvent-Mediated Phase Transformation in Lipid Membranes Induced by Sphingomyelinase. Langmuir 2011, 27, 10050–10060. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Owsianik, G. The Transient Receptor Potential Family of Ion Channels. Genome Biol. 2011, 12, 218. [Google Scholar] [CrossRef] [PubMed]
- Vriens, J.; Nilius, B.; Voets, T. Peripheral Thermosensation in Mammals. Nat. Rev. Neurosci. 2014, 15, 573–589. [Google Scholar] [CrossRef]
- Voets, T.; Vriens, J.; Vennekens, R. Targeting TRP Channels—Valuable Alternatives to Combat Pain, Lower Urinary Tract Disorders, and Type 2 Diabetes? Trends Pharmacol. Sci. 2019, 40, 669–683. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Fliegert, R.; Guse, A.H.; Lü, W.; Du, J. A Structural Overview of the Ion Channels of the TRPM Family. Cell Calcium 2020, 85, 102111. [Google Scholar] [CrossRef]
- Wagner, T.F.J.; Loch, S.; Lambert, S.; Straub, I.; Mannebach, S.; Mathar, I.; Düfer, M.; Lis, A.; Flockerzi, V.; Philipp, S.E.; et al. Transient Receptor Potential M3 Channels Are Ionotropic Steroid Receptors in Pancreatic β Cells. Nat. Cell Biol. 2008, 10, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Vriens, J.; Owsianik, G.; Hofmann, T.; Philipp, S.E.; Stab, J.; Chen, X.; Benoit, M.; Xue, F.; Janssens, A.; Kerselaers, S.; et al. TRPM3 Is a Nociceptor Channel Involved in the Detection of Noxious Heat. Neuron 2011, 70, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Held, K.; Voets, T.; Vriens, J. TRPM3 in Temperature Sensing and Beyond. Temperature 2015, 2, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Held, K.; Kichko, T.; De Clercq, K.; Klaassen, H.; Van Bree, R.; Vanherck, J.-C.; Marchand, A.; Reeh, P.W.; Chaltin, P.; Voets, T.; et al. Activation of TRPM3 by a Potent Synthetic Ligand Reveals a Role in Peptide Release. Proc. Natl. Acad. Sci. USA 2015, 112, E1363–E1372. [Google Scholar] [CrossRef]
- McKemy, D.D.; Neuhausser, W.M.; Julius, D. Identification of a Cold Receptor Reveals a General Role for TRP Channels in Thermosensation. Nature 2002, 416, 52–58. [Google Scholar] [CrossRef]
- Peier, A.M.; Moqrich, A.; Hergarden, A.C.; Reeve, A.J.; Andersson, D.A.; Story, G.M.; Earley, T.J.; Dragoni, I.; McIntyre, P.; Bevan, S.; et al. A TRP Channel That Senses Cold Stimuli and Menthol. Cell 2002, 108, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Bautista, D.M.; Siemens, J.; Glazer, J.M.; Tsuruda, P.R.; Basbaum, A.I.; Stucky, C.L.; Jordt, S.-E.; Julius, D. The Menthol Receptor TRPM8 Is the Principal Detector of Environmental Cold. Nature 2007, 448, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Morenilla-Palao, C.; Pertusa, M.; Meseguer, V.; Cabedo, H.; Viana, F. Lipid Raft Segregation Modulates TRPM8 Channel Activity. J. Biol. Chem. 2009, 284, 9215–9224. [Google Scholar] [CrossRef] [PubMed]
- Sághy, É.; Payrits, M.; Bíró-Sütő, T.; Skoda-Földes, R.; Szánti-Pintér, E.; Erostyák, J.; Makkai, G.; Sétáló, G.; Kollár, L.; Kőszegi, T.; et al. Carboxamido Steroids Inhibit the Opening Properties of Transient Receptor Potential Ion Channels by Lipid Raft Modulation. J. Lipid Res. 2018, 59, 1851–1863. [Google Scholar] [CrossRef]
- Szőke, É.; Börzsei, R.; Tóth, D.M.; Lengl, O.; Helyes, Z.; Sándor, Z.; Szolcsányi, J. Effect of Lipid Raft Disruption on TRPV1 Receptor Activation of Trigeminal Sensory Neurons and Transfected Cell Line. Eur. J. Pharmacol. 2010, 628, 67–74. [Google Scholar] [CrossRef]
- Sághy, É.; Szőke, É.; Payrits, M.; Helyes, Z.; Börzsei, R.; Erostyák, J.; Jánosi, T.Z.; Sétáló, G., Jr.; Szolcsányi, J. Evidence for the Role of Lipid Rafts and Sphingomyelin in Ca2+-Gating of Transient Receptor Potential Channels in Trigeminal Sensory Neurons and Peripheral Nerve Terminals. Pharmacol. Res. 2015, 100, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Horváth, Á.; Biró-Sütő, T.; Kántás, B.; Payrits, M.; Skoda-Földes, R.; Szánti-Pintér, E.; Helyes, Z.; Szőke, É. Antinociceptive Effects of Lipid Raft Disruptors, a Novel Carboxamido-Steroid and Methyl β-Cyclodextrin, in Mice by Inhibiting Transient Receptor Potential Vanilloid 1 and Ankyrin 1 Channel Activation. Front. Physiol. 2020, 11, 559109. [Google Scholar] [CrossRef] [PubMed]
- Horváth, Á.; Payrits, M.; Steib, A.; Kántás, B.; Biró-Süt, T.; Erostyák, J.; Makkai, G.; Sághy, É.; Helyes, Z.; Szőke, É. Analgesic Effects of Lipid Raft Disruption by Sphingomyelinase and Myriocin via Transient Receptor Potential Vanilloid 1 and Transient Receptor Potential Ankyrin 1 Ion Channel Modulation. Front. Pharmacol. 2021, 11, 593319. [Google Scholar] [CrossRef] [PubMed]
- Startek, J.B.; Boonen, B.; López-Requena, A.; Talavera, A.; Alpizar, Y.A.; Ghosh, D.; Van Ranst, N.; Nilius, B.; Voets, T.; Talavera, K. Mouse TRPA1 Function and Membrane Localization Are Modulated by Direct Interactions with Cholesterol. eLife 2019, 8, e46084. [Google Scholar] [CrossRef]
- Startek, J.B.; Talavera, K. Lipid Raft Destabilization Impairs Mouse TRPA1 Responses to Cold and Bacterial Lipopolysaccharides. Int. J. Mol. Sci. 2020, 21, 3826. [Google Scholar] [CrossRef]
- Ferrari, L.F.; Levine, J.D. Plasma Membrane Mechanisms in a Preclinical Rat Model of Chronic Pain. J. Pain 2015, 16, 60–66. [Google Scholar] [CrossRef]
- Lin, C.-L.; Chang, C.-H.; Chang, Y.-S.; Lu, S.-C.; Hsieh, Y.-L. Treatment with Methyl-β-Cyclodextrin Prevents Mechanical Allodynia in Resiniferatoxin Neuropathy in a Mouse Model. Biol. Open 2019, 8, bio039511. [Google Scholar] [CrossRef]
- Sauer, R.-S.; Rittner, H.L.; Roewer, N.; Sohajda, T.; Shityakov, S.; Brack, A.; Broscheit, J.-A. A Novel Approach for the Control of Inflammatory Pain: Prostaglandin E2 Complexation by Randomly Methylated β-Cyclodextrins. Anesth. Analg. 2017, 124, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Julius, D.; Cheng, Y. Structural Snapshots of TRPV1 Reveal Mechanism of Polymodal Functionality. Cell 2021, 184, 5138–5150.e12. [Google Scholar] [CrossRef]
- Horváth, Á.; Erostyák, J.; Szőke, É. Effect of Lipid Raft Disruptors on Cell Membrane Fluidity Studied by Fluorescence Spectroscopy. Int. J. Mol. Sci. 2022, 23, 13729. [Google Scholar] [CrossRef]
- Sántha, P.; Dobos, I.; Kis, G.; Jancsó, G. Role of Gangliosides in Peripheral Pain Mechanisms. Int. J. Mol. Sci. 2020, 21, 1005. [Google Scholar] [CrossRef]
- Drews, A.; Mohr, F.; Rizun, O.; Wagner, T.F.J.; Dembla, S.; Rudolph, S.; Lambert, S.; Konrad, M.; Philipp, S.E.; Behrendt, M.; et al. Structural Requirements of Steroidal Agonists of Transient Receptor Potential Melastatin 3 (TRPM 3) Cation Channels. Br. J. Pharmacol. 2014, 171, 1019–1032. [Google Scholar] [CrossRef]
- Grimm, C.; Kraft, R.; Schultz, G.; Harteneck, C. Activation of the Melastatin-Related Cation Channel TRPM3 by d-Erythro-Sphingosine. Mol. Pharmacol. 2005, 67, 798–805. [Google Scholar] [CrossRef]
- Majeed, Y.; Agarwal, A.; Naylor, J.; Seymour, V.; Jiang, S.; Muraki, K.; Fishwick, C.; Beech, D. Cis-Isomerism and Other Chemical Requirements of Steroidal Agonists and Partial Agonists Acting at TRPM3 Channels: TRPM3 Steroid Stereo-Selectivity. Br. J. Pharmacol. 2010, 161, 430–441. [Google Scholar] [CrossRef]
- Naylor, J.; Li, J.; Milligan, C.J.; Zeng, F.; Sukumar, P.; Hou, B.; Sedo, A.; Yuldasheva, N.; Majeed, Y.; Beri, D.; et al. Pregnenolone Sulphate- and Cholesterol-Regulated TRPM3 Channels Coupled to Vascular Smooth Muscle Secretion and Contraction. Circ. Res. 2010, 106, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Allsopp, R.C.; Lalo, U.; Evans, R.J. Lipid Raft Association and Cholesterol Sensitivity of P2X1-4 Receptors for ATP. J. Biol. Chem. 2010, 285, 32770–32777. [Google Scholar] [CrossRef] [PubMed]
- Payrits, M.; Sághy, É.; Cseko, K.; Pohóczky, K.; Bölcskei, K.; Ernszt, D.; Barabás, K.; Szolcsányi, J.; Ábrahám, I.M.; Helyes, Z.; et al. Estradiol Sensitizes the Transient Receptor Potential Vanilloid 1 Receptor in Pain Responses. Endocrinology 2017, 158, 3249–3258. [Google Scholar] [CrossRef] [PubMed]
- Janssens, A.; Gees, M.; Toth, B.I.; Ghosh, D.; Mulier, M.; Vennekens, R.; Vriens, J.; Talavera, K.; Voets, T. Definition of Two Agonist Types at the Mammalian Cold-Activated Channel TRPM8. eLife 2016, 5, e17240. [Google Scholar] [CrossRef]
- Kelemen, B.; Lisztes, E.; Vladár, A.; Hanyicska, M.; Almássy, J.; Oláh, A.; Szöllősi, A.G.; Pénzes, Z.; Posta, J.; Voets, T.; et al. Volatile Anaesthetics Inhibit the Thermosensitive Nociceptor Ion Channel Transient Receptor Potential Melastatin 3 (TRPM3). Biochem. Pharmacol. 2020, 174, 113826. [Google Scholar] [CrossRef] [PubMed]
- Knowlton, W.M.; Bifolck-Fisher, A.; Bautista, D.M.; McKemy, D.D. TRPM8, but Not TRPA1, Is Required for Neural and Behavioral Responses to Acute Noxious Cold Temperatures and Cold-Mimetics in Vivo. Pain 2010, 150, 340–350. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pretreatment | hTRPM8 | mTRPM3α2 | ||
|---|---|---|---|---|
| Δ(pEC50) of Icilin | Maximal Response to Icilin (Vehicle Pretreated Control = 1) | Δ(pEC50) of PS * | Maximal Response to PS/CIM-0216 (10:1) (Vehicle Pretreated Control = 1) | |
| 30 mU SMase | 0.38 | 0.96 | 0.22 | 0.96 |
| 300 nM Myriocin | 0.31 | 0.92 | 0.22 | 0.98 |
| 3 mM MCD | - | - | −0.04 | 1.10 |
| 10 mM MCD | 0.24 | 1.14 | - | - |
| 100 µM C1 | 0.33 | 1.21 | −0.19 | 1.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horváth, Á.; Steib, A.; Nehr-Majoros, A.; Kántás, B.; Király, Á.; Racskó, M.; Tóth, B.I.; Szánti-Pintér, E.; Kudová, E.; Skoda-Földes, R.; et al. Anti-Nociceptive Effects of Sphingomyelinase and Methyl-Beta-Cyclodextrin in the Icilin-Induced Mouse Pain Model. Int. J. Mol. Sci. 2024, 25, 4637. https://doi.org/10.3390/ijms25094637
Horváth Á, Steib A, Nehr-Majoros A, Kántás B, Király Á, Racskó M, Tóth BI, Szánti-Pintér E, Kudová E, Skoda-Földes R, et al. Anti-Nociceptive Effects of Sphingomyelinase and Methyl-Beta-Cyclodextrin in the Icilin-Induced Mouse Pain Model. International Journal of Molecular Sciences. 2024; 25(9):4637. https://doi.org/10.3390/ijms25094637
Chicago/Turabian StyleHorváth, Ádám, Anita Steib, Andrea Nehr-Majoros, Boglárka Kántás, Ágnes Király, Márk Racskó, Balázs István Tóth, Eszter Szánti-Pintér, Eva Kudová, Rita Skoda-Földes, and et al. 2024. "Anti-Nociceptive Effects of Sphingomyelinase and Methyl-Beta-Cyclodextrin in the Icilin-Induced Mouse Pain Model" International Journal of Molecular Sciences 25, no. 9: 4637. https://doi.org/10.3390/ijms25094637
APA StyleHorváth, Á., Steib, A., Nehr-Majoros, A., Kántás, B., Király, Á., Racskó, M., Tóth, B. I., Szánti-Pintér, E., Kudová, E., Skoda-Földes, R., Helyes, Z., & Szőke, É. (2024). Anti-Nociceptive Effects of Sphingomyelinase and Methyl-Beta-Cyclodextrin in the Icilin-Induced Mouse Pain Model. International Journal of Molecular Sciences, 25(9), 4637. https://doi.org/10.3390/ijms25094637