Micro-Food Web Structure Shapes Rhizosphere Microbial Communities and Growth in Oak


Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Substrate
2.2. Ectomycorrhizal Inoculum
2.3. Oak Seedlings
2.4. Nematode Inoculum
2.5. Experimental Setup and Harvest
2.6. Soil Phospholipid Fatty Acid Analysis
2.7. Statistical Analysis
3. Results
3.1. Nematode Community Structure
3.2. Impact of the Nematode Trophic Structure on the Microbial Community
3.2.1. Biomass of Microorganisms
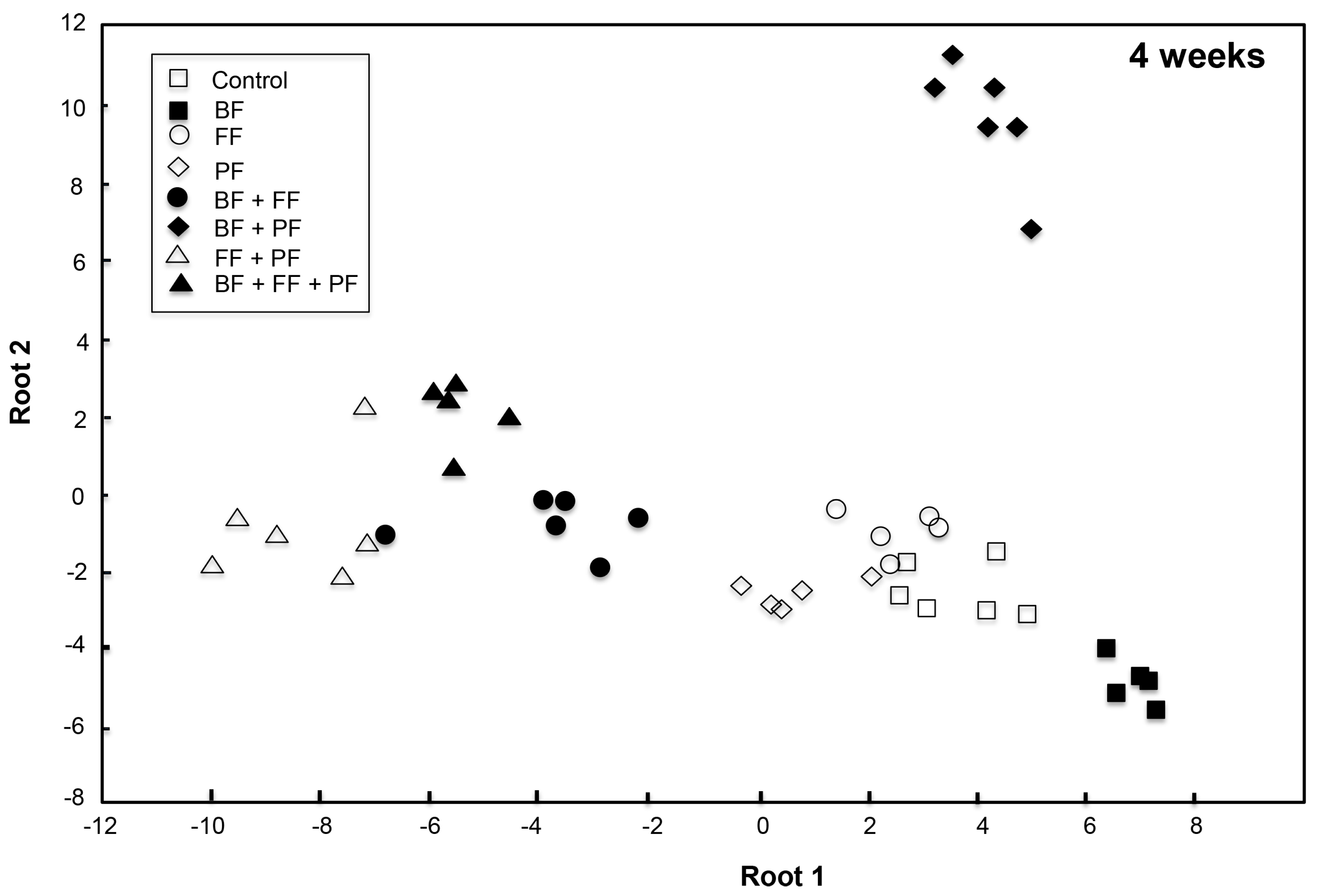
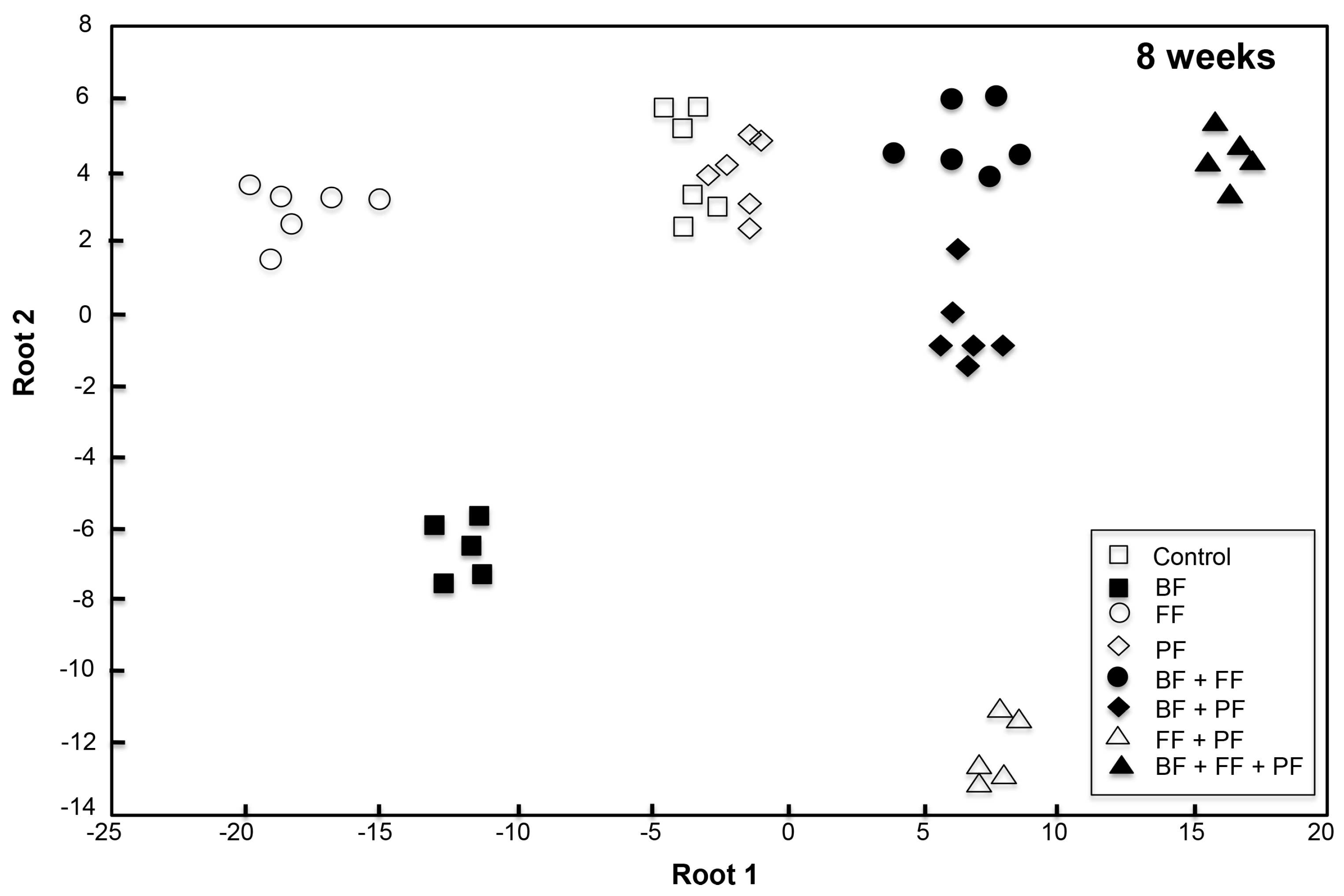
3.2.2. Community Structure of Microorganisms
3.3. Impact of the Nematode Trophic Structure on Oak Growth
4. Discussion
4.1. Experimental Manipulation of Trophic Structure in the Oak Rhizosphere
4.2. Effects of Multitrophic Interactions on Rhizosphere Microorganisms and Oak Performance
4.2.1. Plant Feeders
4.2.2. Bacterial Feeders
4.2.3. Fungal Feeders
4.2.4. Multitrophic Interactions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Högberg, P.; Nordgren, A.; Buchmann, N.; Taylor, A.F.S.; Ekblad, A.; Högberg, M.N.; Nyberg, G.; Ottosson-Löfvenius, M.; Read, D.J. Large-scale forest girdling shows that current photosynthesis drives soil respiration. Nature 2001, 411, 789–792. [Google Scholar] [CrossRef] [PubMed]
- Prescott, C.E.; Grayston, S.J. Tree species influence on microbial communities in litter and soil: Current knowledge and research needs. For. Ecol. Manag. 2013, 309, 19–27. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Courty, P.E.; Buée, M.; Diedhiou, A.G.; Frey-Klett, P.; Le Tacon, F.; Rineau, F.; Uroz, S.; Garbaye, J. The role of ectomycorrhizal communities in forest ecosystem processes: New perspectives and emerging concepts. Soil Biol. Biochem. 2010, 42, 679–698. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Smith, S.E.; Smith, F.A. Structure and function of the interfaces in biotrophic symbioses as they relate to nutrient transport. New Phytol. 1990, 114, 1–38. [Google Scholar] [CrossRef]
- Bardgett, R. The Biology of Soil: A Community and Ecosystem Approach; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Cole, L.; Staddon, P.L.; Sleep, D.; Bardgett, R.D. Soil animals influence microbial abundance, but not plant-microbial competition for soil organic nitrogen. Funct. Ecol. 2004, 18, 631–640. [Google Scholar] [CrossRef]
- Yeates, G.W.; Bongers, T.; de Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding habits in soil nematode families and genera—An outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar] [PubMed]
- Scharroba, A.; Dibbern, D.; Hünninghaus, M.; Kramer, S.; Moll, S.; Butenschoen, O.; Bonkowski, M.; Buscot, F.; Kandeler, E.; Koller, R.; et al. Effects of resource availability and quality on the structure of the micro-food web of an arable soil across depth. Soil Biol. Biochem. 2012, 50, 111–119. [Google Scholar] [CrossRef]
- Scharroba, A.; Kramer, S.; Kandeler, E.; Ruess, L. Spatial and temporal variation of resource allocation in an arable soil drives community structure and biomass of nematodes and their role in the micro-food web. Pedobiologia 2016, 59, 111–120. [Google Scholar] [CrossRef]
- Yeates, G.W. Nematodes in ecological webs. In Encyclopedia of Life Science; Wiley & Sons: Chichester, UK, 2010; pp. 1–10. [Google Scholar]
- Djigal, D.; Brauman, A.; Diop, T.A.; Chotte, J.L.; Villenave, C. Influence of bacterial-feeding nematodes (Cephalobidae) on soil microbial communities during maize growth. Soil Biol. Biochem. 2004, 36, 323–331. [Google Scholar] [CrossRef]
- Neher, D.A. Ecology of plant and free-living nematodes in natural and agricultural soil. Ann. Rev. Phytopathol. 2010, 48, 371–394. [Google Scholar] [CrossRef] [PubMed]
- Yeates, G.W.; Saggar, S.; Denton, C.S.; Mercer, C.F. Impact of clover cyst nematode (Heterodera trifolii) infection on soil microbial activity in the rhizosphere of white clover (Trifolium repens)—A pulse-labelling experiment. Nematologica 1998, 44, 81–90. [Google Scholar] [CrossRef]
- Yeates, G.W.; Saggar, S.; Hedley, C.B.; Mercer, C.F. Increase in 14C-carbon translocation to soil microbial biomass when five species of plant-parasitic nematodes infect roots of white clover. Nematology 1999, 1, 295–300. [Google Scholar] [CrossRef]
- Poll, J.; Marhan, S.; Haase, S.; Hallmann, J.; Kandeler, E.; Ruess, L. Low amounts of herbivory by root-knot nematodes affect microbial community dynamics and carbon allocation in the rhizosphere. FEMS Microbiol. Ecol. 2007, 62, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ferris, H. The effects of nematode grazing on nitrogen mineralization during fungal decomposition of organic matter. Soil Biol. Biochem. 1999, 31, 1265–1279. [Google Scholar] [CrossRef]
- Ferris, H.; Venette, R.C.; Scow, K.M. Soil management to enhance bacterivore and fungivore nematode populations and their nitrogen mineralisation function. Appl. Soil Ecol. 2004, 25, 19–35. [Google Scholar] [CrossRef]
- Sutherland, J.R.; Webster, J.M. Nematode pests of forest trees. In Plant Parasitic Nematodes in Temperate Agriculture; Evans, K., Trudgill, D.L., Webster, J.M., Eds.; CAB International: Tuscon, AZ, USA, 1993; pp. 351–380. [Google Scholar]
- Francl, L.J. Interactions of nematodes with mycorrhizae and mycorrhizal fungi. In Nematode Interactions; Kahn, M.W., Ed.; Chapman & Hall: New York, NY, USA, 1993; pp. 203–216. [Google Scholar]
- Gilbert, G.S. Evolutionary ecology of plant diseases in natural ecosystems. Ann. Rev. Phytopathol. 2002, 40, 13–43. [Google Scholar] [CrossRef] [PubMed]
- Wisz, M.S.; Pottier, J.; Kissling, W.D.; Pellissier, L.; Lenoir, J.; Damgaard, C.F.; Dormann, C.F.; Forchhammer, M.C.; Grytnes, J.A.; Guisan, A.; et al. The role of biotic interactions in shaping distributions and realised assemblages of species: Implications for species distribution modelling. Biol. Rev. 2013, 88, 15–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Praciak, A.; Pasiecznik, N.M.; Sheil, D.; van Heist, M.; Sassen, M.; Correia, C.S.; Dixon, C.; Fyson, G.E.; Rushforth, K.; Teeling, C. The CABI Encyclopedia of Forest Trees; CABI: Wallingford, UK, 2013. [Google Scholar]
- Caravaca, F.; Maboreke, H.; Kurth, F.; Hermann, S.; Tarkka, T.M.; Ruess, L. Synergists and antagonists in the rhizosphere modulate microbial communities and growth of Quercus robur L. Soil Biol. Biochem. 2015, 82, 65–73. [Google Scholar] [CrossRef]
- Maboreke, H.R.; Graf, M.; Grams, T.E.E.; Herrmann, S.; Scheu, S.; Ruess, L. Multitrophic interactions in the rhizosphere of a temperate forest tree affect plant carbon flow into the belowground food web. Soil Biol. Biochem. 2017, 115, 526–536. [Google Scholar] [CrossRef]
- Coûteaux, M.M.; Mousseau, M.; Celerier, M.L.; Bottner, P. Increased atmospheric CO2 and litter quality: Decomposition of sweet chestnut leaf litter with animal food webs of different complexities. Oikos 1991, 61, 54–64. [Google Scholar] [CrossRef]
- Setälä, H.; Laakso, J.; Mikola, J.; Huhta, V. Functional diversity of decomposer organisms in relation to primary production. Appl. Soil Ecol. 1998, 9, 25–31. [Google Scholar] [CrossRef]
- Laakso, J.; Setälä, H. Sensitivity of primary production to changes in the architecture of belowground food webs. Oikos 1999, 87, 57–64. [Google Scholar] [CrossRef]
- Setälä, H. Growth of birch and pine seedlings in relation to grazing by soil fauna on ectomycorrhizal fungi. Ecology 1995, 76, 1844–1851. [Google Scholar] [CrossRef]
- Gebremikael, M.T.; Steel, H.; Buchan, D.; Bert, W.; de Neve, S. Nematodes enhance plant growth and nutrient uptake under C and N-rich conditions. Nat. Sci. Rep. 2016, 6, 32862. [Google Scholar] [CrossRef] [PubMed]
- Van Gestel, C.A.M.; Kruidenier, M.; Berg, M.P. Suitability of wheat straw decomposition, cotton strip degradation and bait-lamina feeding tests to determine soil invertebrate activity. Biol. Fertil. Soils 2003, 37, 115–123. [Google Scholar]
- Herrmann, S.; Buscot, F. Cross talks at the morphogenetic, physiological and gene regulation levels between the mycobiont Piloderma croceum and oak microcuttings (Quercus robur) during formation of ectomycorrhizas. Phytochemistry 2007, 68, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Tarkka, M.T.; Herrmann, S.; Wubet, T.; Feldhahn, L.; Recht, S.; Kurth, F.; Mailander, S.; Bonn, M.; Neef, M.; Angay, O.; et al. OakContigDF159.1, a reference library for studying differential gene expression in Quercus robur during controlled biotic interactions: Use for quantitative transcriptomic profiling of oak roots in ectomycorrhizal symbiosis. New Phytol. 2013, 199, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Maboreke, H. Effect of Plant-Parasitic Nematodes on Rhizosphere Interactions in Oaks. Ph.D. Thesis, Humboldt-Universität zu Berlin, Berlin, Germany, 2016. [Google Scholar]
- Marx, D.H. Influence of ectotrophic mycorrhizal fungi on resistance of pine roots to pathogenic infections. I. Antagonsim of mycorrhizal fungi to pathogenic fungi and soil bacteria. Phytopathology 1969, 59, 153–163. [Google Scholar]
- Ruess, L.; Dighton, J. Cultural studies on soil nematodes and their fungal hosts. Nematologica 1996, 42, 330–346. [Google Scholar] [CrossRef]
- O’Bannon, J.H.; Taylor, A.L. Migratory endoparasitic nematodes reared on carrot discs. Phytopathology 1968, 58, 385. [Google Scholar]
- Ruess, L. Studies on the nematode fauna of an acid forest soil: Spatial distribution and extraction. Nematologica 1995, 41, 229–239. [Google Scholar] [CrossRef]
- Byrd, D.W.; Kirkpatrick, T.; Barker, K.R. An improved technique for clearing and staining plant tissues for detection of nematodes. J. Nematol. 1983, 15, 42–143. [Google Scholar]
- Frostegård, A.; Tunlid, A.; Bååth, E. Phospholipids fatty acid composition, biomass and activity of microbial communities from two soil types experimentally exposed to different heavy metals. Appl. Environ. Microbiol. 1993, 59, 3605–3617. [Google Scholar] [PubMed]
- Ruess, L.; Chamberlain, P.M. The fat that matters: Soil food web analysis using fatty acids and their carbon stable isotope signature. Soil Biol. Biochem. 2010, 42, 1898–1910. [Google Scholar] [CrossRef]
- Frostegård, A.; Bååth, E. The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biol. Fertil. Soils 1996, 22, 59–65. [Google Scholar] [CrossRef]
- Gebremikael, M.T.; Buchan, D.; de Neve, S. Quantifying the influence of free-living nematodes on soil nitrogen and microbial biomass dynamics in bare and planted microcosms. Soil Biol. Biochem. 2014, 70, 131–141. [Google Scholar] [CrossRef]
- Huhta, V.; Wright, D.H.; Coleman, D.C. Characteristics of defaunated soil. I. A comparison of three techniques applied to two different forest soils. Pedobiologia 1989, 33, 415–424. [Google Scholar]
- Buchan, D.; Moeskops, B.; Ameloot, N.; de Neve, S.; Sleutel, S. Selective sterilisation of undisturbed soil cores by gamma irradiation: Effects on free-living nematodes, microbial community and nitrogen dynamics. Soil Biol. Biochem. 2012, 47, 10–13. [Google Scholar] [CrossRef]
- Stenberg, B.; Johansson, M.; Pell, M.; Sjodahl-Svensson, K.; Stenstrom, J.; Torstensson, L. Microbial biomass and activities in soil as affected by frozen and cold storage. Soil Biol. Biochem. 1998, 30, 393–402. [Google Scholar] [CrossRef]
- Kampichler, C.; Bruckner, A.; Baumgarten, A.; Berthold, A.; Zechmeister-Boltenstern, S. Field mesocosms for assessing biotic processes in soils: How to avoid side effects. Eur. J. Soil Biol. 1999, 35, 135–143. [Google Scholar] [CrossRef]
- Griffiths, B.S.; Bengough, A.G.; Neilson, R.; Trudgill, D.L. The extent to which nematode communities are affected by soil factors—A pot experiment. Nematology 2002, 4, 943–952. [Google Scholar] [CrossRef]
- Griffiths, B.S.; Neilson, R.; Bengough, A.G. Soil factors determined nematode community composition in a two years pot experiment. Nematology 2003, 5, 889–897. [Google Scholar] [CrossRef]
- Griffiths, B.S.; Young, I.M.; Boag, B. Nematodes associated with the rhizosphere of barley (Hordeum vulgare). Pedobiologia 1991, 35, 265–272. [Google Scholar]
- Waldrop, M.P.; Balser, T.C.; Firestone, M.K. Linking microbial community composition to function in a tropical soil. Soil Biol. Biochem. 2000, 32, 1837–1846. [Google Scholar] [CrossRef]
- Pausch, J.; Hofmann, S.; Scharroba, A.; Kuzyakov, Y.; Ruess, L. Fluxes of root-derived carbon into nematode micro-food web of an arable soil. Food Webs 2016, 9, 39–45. [Google Scholar] [CrossRef]
- Maboreke, H.R.; Feldhahn, L.; Bonn, M.; Tarkka, M.T.; Buscot, F.; Herrmann, S.; Menzel, R.; Ruess, L. Transcriptome analysis in oak uncovers a strong impact of endogenous rhythmic growth on interaction with plant-parasitic nematodes. BMC Genom. 2016, 17, 627. [Google Scholar] [CrossRef] [PubMed]
- Postma-Blaauw, M.B.; de Vries, F.T.; de Goede, R.G.M.; Bloem, J.; Faber, J.H.; Brussaard, L. Within-trophic group interactions of bacterivorous nematode species and their effects on the bacterial community and nitrogen mineralization. Oecologia 2005, 142, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Elliott, E.T.; Anderson, R.V.; Coleman, D.C.; Cole, C.V. Habitable pore space and microbial trophic interactions. Oikos 1980, 35, 327–335. [Google Scholar] [CrossRef]
- Salinas, K.A.; Edenborn, S.L.; Sexstone, A.J.; Kotcon, J.B. Bacterial preferences of the bacterivorous soil nematode Cephalobus brevicauda (Cephalobidae): Effect of bacterial type and size. Pedobiologia 2007, 51, 55–64. [Google Scholar] [CrossRef]
- Abada, E.A.; Sung, H.; Dwivedi, M.; Park, B.J.; Lee, S.K.; Ahnn, J. C. elegans behaviour of preference choice on bacterial food. Mol. Cells 2009, 28, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.F.; Griffiths, B.; Chen, X.; Liu, M.; Jiao, J.; Hu, F.; Li, H. Influence of bacterial-feeding nematodes on nitrification and the ammonia-oxidizing bacteria (AOB) community composition. Appl. Soil Ecol. 2010, 45, 131–137. [Google Scholar] [CrossRef]
- Griffiths, B.S.; Bonkowski, M.; Dobson, G.; Caul, S. Changes in soil microbial community structure in the presence of microbial-feeding nematodes and protozoa. Pedobiologia 1999, 43, 297–304. [Google Scholar]
- Mazzola, M. Manipulation of rhizosphere bacterial communities to induce suppressive soils. J. Nematol. 2007, 39, 213–220. [Google Scholar] [PubMed]
- Ruess, L.; Häggblom, M.M.; Garzía Zapata, E.J.; Dighton, J. Fatty acids of fungi and nematodes—Possible biomarkers in the soil food chain? Soil Biol. Biochem. 2002, 34, 745–756. [Google Scholar] [CrossRef]
- Setälä, H.; Rissanen, J.; Markkola, A.M. Conditional outcomes in the relationship between pine and ectomycorrhizal fungi in relation to biotic and abiotic environment. Oikos 1997, 80, 112–122. [Google Scholar] [CrossRef]
- Mamilov, A.Sh.; Byzov, B.A.; Zvyagintsev, D. G.; Dilly, O.M. Predation on fungal and bacterial biomass in a soddy-podzolic soil amended with starch, wheat straw and alfalfa meal. Appl. Soil Ecol. 2001, 16, 131–139. [Google Scholar] [CrossRef]
- Denton, C.S.; Bardgett, R.D.; Cook, R.; Hobbs, P.J. Low amounts of root herbivory positively influence the rhizosphere microbial community in a temperate grassland soil. Soil Biol. Biochem. 1999, 31, 155–165. [Google Scholar] [CrossRef]
- Tu, C.; Koenning, S.R.; Hu, S. Root-parasitic nematodes enhance soil microbial activities and nitrogen mineralization. Microb. Ecol. 2003, 46, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Wardle, D.A.; Yeates, G.W. The dual importance of competition and predation as regulatory forces in terrestrial ecosystems-evidence from decomposer food webs. Oecologia 1993, 93, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Setälä, H.; Huhta, V. Evaluation of the soil fauna impact on decomposition in a simulated coniferous forest soil. Biol. Fertil. Soils 1990, 10, 163–169. [Google Scholar]
- Bakhtiar, Y.; Miller, D.; Cavagnaro, T.; Smith, S. Interactions between two arbuscular mycorrhizal fungi and fungivorous nematodes and control of nematodes with fenamifos. Appl. Soil Ecol. 2001, 17, 107–117. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Control | BF | FF | PF | BF+FF | BF+PF | FF+PF | BF+FF+PF | ANOVA | ||
|---|---|---|---|---|---|---|---|---|---|---|
| 4 weeks | F7,37 | P | ||||||||
| Total amount | 36.1 ± 2.0cd | 35.7 ± 0.5cd | 34.9 ± 0.7de | 36.5 ± 1.5bcd | 37.7 ± 1.3abc | 32.7 ± 1.5de | 39.0 ± 0.6a | 38.9 ± 0.6ab | 16.21 | <0.001 |
| Gram+ | 8.6 ± 0.2cd | 8.7 ± 0.1bcd | 8.5 ± 0.4d | 8.7 ± 0.2bcd | 9.1 ± 0.4ab | 7.1 ± 0.2e | 9.3 ± 0.4a | 9.0 ± 0.0abc | 39.53 | <0.001 |
| Gram− | 3.4 ± 0.1cd | 3.3 ± 0.0cd | 3.3 ± 0.0d | 3.5 ± 0.1c | 3.7 ± 0.1b | 3.3 ± 0.0d | 4.0 ± 0.1a | 3.8 ± 0.0ab | 52.17 | <0.001 |
| Bacteria | 6.8 ± 0.6ab | 6.7 ± 0.1ab | 6.3 ± 0.0bc | 6.5 ± 0.5abc | 6.7 ± 0.1ab | 6.06 ± 0.32bc | 7.1 ± 0.1a | 7.0 ± 0.0a | 6.40 | <0.001 |
| Actinobacteria | 4.4 ± 0.3abc | 4.3 ± 0.1bc | 4.3 ± 0.1bc | 4.4 ± 0.4abc | 4.7 ± 0.1ab | 4.2 ± 0.4c | 4.8 ± 0.2a | 4.8 ± 0.1a | 6.11 | <0.001 |
| Fungi | 3.9 ± 0.6 | 3.7 ± 0.5 | 3.7 ± 0.4 | 3.9 ± 0.6 | 4.1 ± 0.2 | 3.9± 0.5 | 4.2 ± 0.1 | 4.3 ± 0.1 | n.s | n.s |
| F/B ratio | 0.2 ± 0.00 | 0.16 ± 0.02 | 0.17 ± 0.01 | 0.17 ± 0.02 | 0.17 ± 0.01 | 0.19 ± 0.02 | 0.17 ± 0.01 | 0.18 ± 0.00 | n.s | n.s |
| 8 weeks | F7, 39 | P | ||||||||
| Total amount | 33.4 ± 0.8d | 33.7 ± 0.8d | 30.6 ± 0.5e | 34.0 ± 0.3d | 35.4 ± 0.8c | 36.3 ± 0.7bc | 36.8 ± 0.4b | 38.1 ± 0.4a | 83.90 | <0.001 |
| Gram+ | 7.9 ± 0.1c | 7.6 ± 0.22d | 7.2 ± 0.2e | 8.2 ± 0.2bc | 8.3 ± 0.2b | 8.8 ± 0.1a | 8.3 ± 0.1b | 8.7 ± 0.0a | 83.00 | <0.001 |
| Gram− | 3.3 ± 0.1d | 3.3 ± 0.0d | 3.1 ± 0.0e | 3.5 ± 0.0c | 3.70 ± 0.1b | 3.7 ± 0.1b | 3.6 ± 0.0c | 4.0 ± 0.1a | 178.90 | <0.001 |
| Bacteria | 5.9 ± 0.1c | 5.5 ± 0.1d | 5.1 ± 0.1e | 5.6 ± 0.1d | 6.3 ± 0.1b | 6.0 ± 0.1c | 6.3 ± 0.1 b | 6.9 ± 0.1a | 190.20 | <0.001 |
| Actinobacteria | 4.2 ± 0.1e | 4.3 ± 0.1de | 3.8 ± 0.1f | 4.3 ± 0.1e | 4.5 ± 0.0b | 4.5 ± 0.1bc | 4.4 ± 0.1 cd | 4.8 ± 0.1a | 132.20 | <0.001 |
| Fungi | 3.6 ± 0.1d | 3.8 ± 0.1c | 3.1 ± 0.1e | 3.8 ± 0.1c | 4.02 ± 0.1b | 4.1 ± 0.1b | 5.0 ± 0.1a | 4.2 ± 0.1b | 227.66 | <0.001 |
| F/B ratio | 0.17 ± 0.01e | 0.18 ± 0.00b | 0.16 ± 0.00e | 0.18 ± 0.00cd | 0.18 ± 0.00bcd | 0.18 ± 0.00bc | 0.22 ± 0.01a | 0.17 ± 0.00de | 120.83 | <0.001 |
| 4 weeks | 8 weeks | |||
|---|---|---|---|---|
| Root 1 | Root 2 | Root 1 | Root 2 | |
| i15:0 (G+) | −0.540 *** | −0.806 *** | 0.865 *** | 0.014 |
| a15:0 (G+) | −0.519 *** | −0.804 *** | 0.779 *** | 0.189 |
| i16:0 (G+) | −0.607 *** | −0.275 | 0.756 *** | −0.120 |
| i17:0 (G+) | −0.407 ** | −0.343 * | 0.643 *** | −0.165 |
| a17:0 (G+) | −0.462 ** | −0.282 | 0.902 *** | 0.187 |
| cy17:0 (G−) | −0.935 *** | −0.176 | 0.902 *** | 0.163 |
| cy19:0 (G−) | −0.736 *** | −0.092 | 0.806 *** | −0.066 |
| 16:1 ω7c (B) | −0.430 ** | −0.442 | 0.914 *** | 0.237 |
| 16:1 ω5c (B) | −0.238 | −0.531 | 0.871 *** | −0.141 |
| 18:1 ω9t (B) | −0.454 ** | −0.155 | 0.793 *** | −0.271 |
| 16:0 10-meth (A) | −0.589 *** | −0.308 * | 0.874 *** | −0.042 |
| 18:0 10-meth (A) | −0.590 *** | 0.097 | 0.805 *** | 0.033 |
| 18:2 ω6,9c (F) | −0.224 | 0.183 | 0.486 ** | −0.822 *** |
| 18:1 ω9c (F) | −0.4186 ** | 0.072 | 0.864 *** | −0.371 * |
| 14:0 | −0.225 | −0.408 ** | 0.389 | 0.240 |
| 15:0 | 0.145 | −0.561 *** | 0.349 | 0.355* |
| 16:0 | −0.645 *** | −0.416 ** | 0.364 | −0.457 ** |
| 18:0 | −0.496 ** | 0.117 | 0.000 | −0.334 * |
| Control | BF | FF | PF | BF + FF | BF + PF | FF + PF | BF + FF + PF | |
|---|---|---|---|---|---|---|---|---|
| 4 weeks | ||||||||
| Root length (cm) | 38.8 ± 8.1 | 42.8 ± 21.5 | 37.5 ± 13.8 | 37.5 ± 10.1 | 37.2 ± 15.0 | 38.3 ± 13.7 | 35.7 ± 11.6 | 45.7 ± 10.6 |
| Root biomass (g) | 3.8 ± 0.7 | 3.1 ± 1.4 | 3.2 ± 0.7 | 3.2 ± 0.7 | 3.2 ± 0.7 | 3.2 ± 0.9 | 3.8 ± 1.1 | 3.9 ± 0.9 |
| Shoot biomass (g) | 2.6 ± 0.6 | 2.4 ± 0.5 | 2.7 ± 0.3 | 2.6 ± 0.3 | 2.9 ± 0.7 | 2.5 ± 0.8 | 2.6 ± 0.3 | 2.7 ± 0.6 |
| root/shoot ratio | 1.5 ± 0.4 | 1.3 ± 0.6 | 1.2 ± 0.3 | 1.3 ± 0.4 | 1.2 ± 0.4 | 1.3 ± 0.2 | 1.5 ± 0.4 | 1.5 ± 0.5 |
| 8 weeks | ||||||||
| Root length (cm) | 39.0 ± 10.2 | 30.3 ± 10.0 | 35.3 ± 13.7 | 41.7 ± 9.4 | 46.5 ± 5.8 | 35.5 ± 12.9 | 35.0 ± 11.5 | 43.8 ± 9.4 |
| Root biomass (g) | 6.7 ± 0.7 | 5.8 ± 1.3 | 5.5 ± 0.9 | 4.7 ± 1.1 | 6.3 ± 2.1 | 5.8 ± 0.7 | 4.8 ± 0.6 | 6.8 ± 2.7 |
| Shoot biomass (g) | 3.0 ± 0.8 | 3.2 ± 0.6 | 3.2 ± 0.4 | 3.0 ± 0.5 | 3.1 ± 0.7 | 3.0 ± 0.6 | 3.2 ± 0.5 | 2.9 ± 0.5 |
| root/shoot ratio | 2.4 ± 0.2ab | 1.9 ± 0.1c | 1.8 ± 0.3cd | 1.5 ± 0.1d | 2.1 ± 0.1bc | 2.0 ± 0.2c | 1.5 ± 0.1d | 2.4 ± 0.1a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maboreke, H.R.; Bartel, V.; Seiml-Buchinger, R.; Ruess, L. Micro-Food Web Structure Shapes Rhizosphere Microbial Communities and Growth in Oak. Diversity 2018, 10, 15. https://doi.org/10.3390/d10010015
Maboreke HR, Bartel V, Seiml-Buchinger R, Ruess L. Micro-Food Web Structure Shapes Rhizosphere Microbial Communities and Growth in Oak. Diversity. 2018; 10(1):15. https://doi.org/10.3390/d10010015
Chicago/Turabian StyleMaboreke, Hazel R., Veronika Bartel, René Seiml-Buchinger, and Liliane Ruess. 2018. "Micro-Food Web Structure Shapes Rhizosphere Microbial Communities and Growth in Oak" Diversity 10, no. 1: 15. https://doi.org/10.3390/d10010015
APA StyleMaboreke, H. R., Bartel, V., Seiml-Buchinger, R., & Ruess, L. (2018). Micro-Food Web Structure Shapes Rhizosphere Microbial Communities and Growth in Oak. Diversity, 10(1), 15. https://doi.org/10.3390/d10010015