Divergence, Convergence and Phenotypic Diversity of Neotropical Frugivorous Bats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cranial Morphology
2.2. Convergent Evolution in Neotropical Frugivorous Bats
2.3. Test and Quantification of Convergence
3. Results
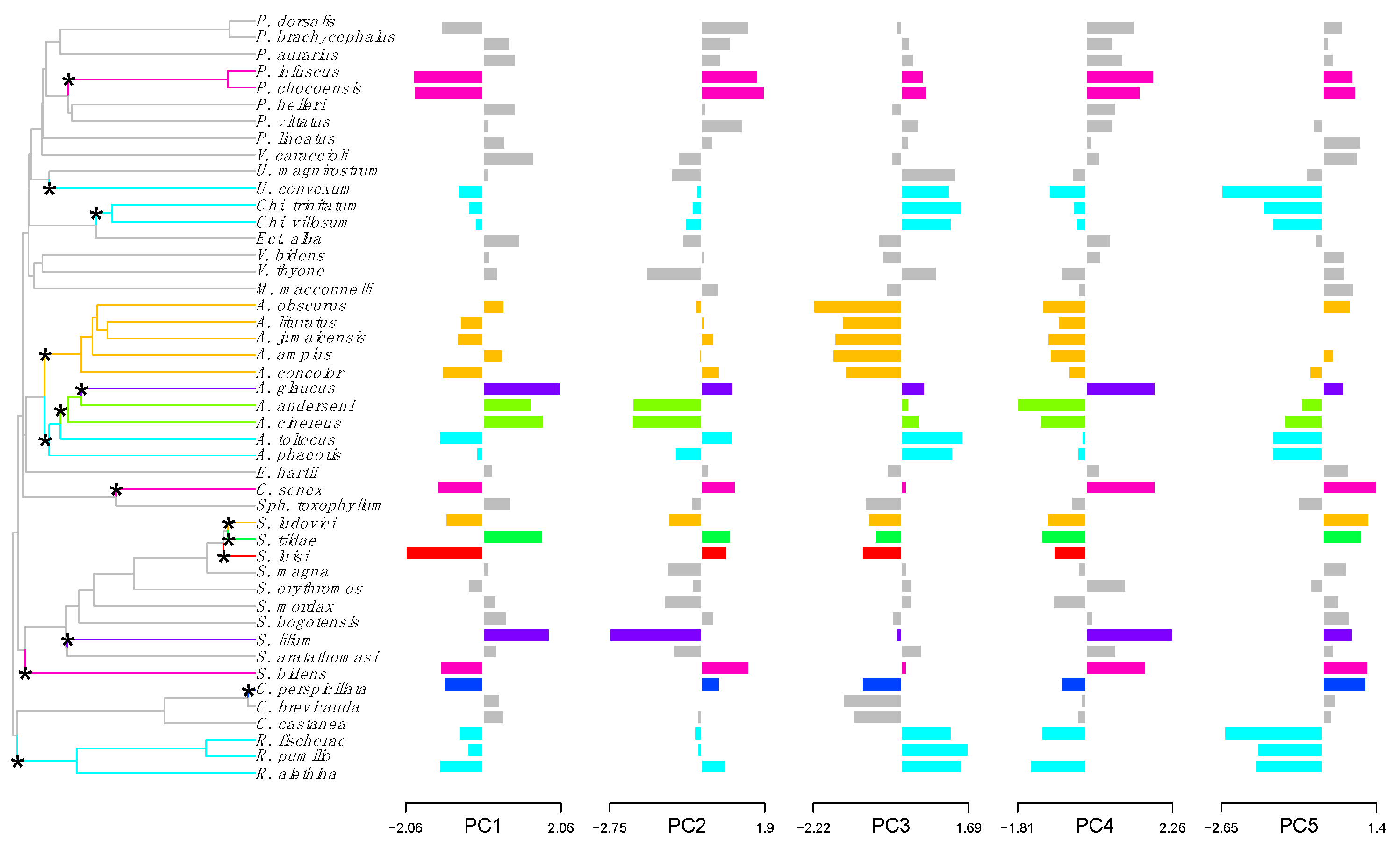
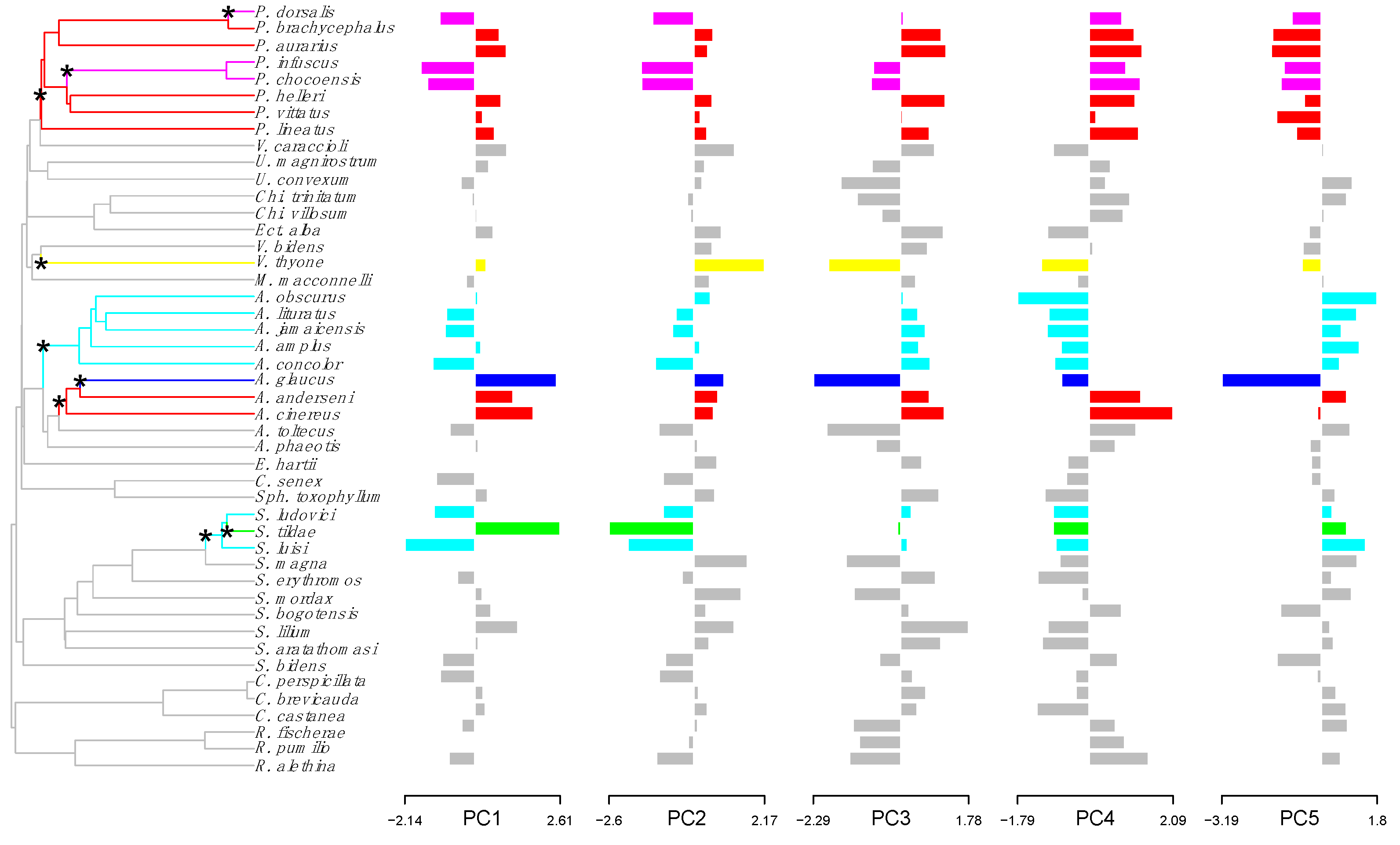
3.1. Cranial Morphology
3.2. Convergent Evolution in Neotropical Frugivorous Bats
3.3. Test and Quantification of Convergence
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Losos, J.B. The evolution of form and function: Morphology and locomotor performance in West Indian Anolis lizards. Evolution 1990, 44, 1189–1203. [Google Scholar] [CrossRef] [PubMed]
- Schluter, D. The Ecology of Adaptive Radiation; Oxford University Press: New York, NY, USA, 2000; ISBN 9780191588327. [Google Scholar]
- Gavrilets, S.; Losos, J.B. Adaptive radiation: Contrasting theory with data. Science 2009, 323, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Dumont, E.R.; Davalos, L.M.; Goldberg, A.; Santana, S.E.; Rex, K.; Voigt, C.C. Morphological innovation, diversification and invasion of a new adaptive zone. Proc. R. Soc. B Biol. Sci. 2012, 279, 1797–1805. [Google Scholar] [CrossRef] [PubMed]
- Parent, C.E.; Crespi, B.J. Ecological opportunity in adaptive radiation of Galápagos endemic land Snails. Am. Nat. 2009, 174, 898–905. [Google Scholar] [CrossRef] [PubMed]
- Losos, J.B. Adaptive radiation, ecological opportunity, and evolutionary determinism. Am. Nat. 2010, 175, 623–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellborn, G.A.; Langerhans, R.B. Ecological opportunity and the adaptive diversification of lineages. Ecol. Evol. 2015, 5, 176–195. [Google Scholar] [CrossRef] [PubMed]
- Burress, E.D.; Holcomb, J.M.; Tan, M.; Armbruster, J.W. Ecological diversification associated with the benthic-to-pelagic transition by North American minnows. J. Evol. Biol. 2017, 30, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Simpson, G.G. The Major Features of Evolution; Columbia University Press: New York, NY, USA, 1953. [Google Scholar]
- Mahler, D.L.; Revell, L.J.; Glor, R.E.; Losos, J.B. Ecological opportunity and the rate of morphological evolution in the diversification of greater Antillean anoles. Evolution 2010, 64, 2731–2745. [Google Scholar] [CrossRef] [PubMed]
- Nosil, P.; Reimchen, T.E. Ecological opportunity and levels of morphological variance within freshwater stickleback populations. Biol. J. Linn. Soc. 2005, 86, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Harmon, L.J.; Melville, J.; Larson, A.; Losos, J.B. The role of geography and ecological opportunity in the diversification of day geckos (Phelsuma). Syst. Biol. 2008, 57, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Wiens, J.J.; Graham, C.H. Niche Conservatism: Integrating evolution, ecology, and conservation biology. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 519–539. [Google Scholar] [CrossRef]
- Moen, D.S.; Irschick, D.J.; Wiens, J.J. Evolutionary conservatism and convergence both lead to striking similarity in ecology, morphology and performance across continents in frogs. Proc. R. Soc. B Biol. Sci. 2013, 280, 20132156. [Google Scholar] [CrossRef] [PubMed]
- Losos, J.B.; Warheitt, K.I.; Schoener, T.W. Adaptive differentiation following experimental island colonization in Anolis lizards. Nature 1997, 387, 70–73. [Google Scholar] [CrossRef]
- Rüber, L.; Adams, D.C. Evolutionary convergence of body shape and trophic morphology in cichlids from Lake Tanganyika. J. Evol. Biol. 2001, 14, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, A.Y.; Rubinoff, D. Convergent evolution of morphology and habitat use in the explosive Hawaiian fancy case caterpillar radiation. J. Evol. Biol. 2013, 26, 1763–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingram, T.; Kai, Y. The geography of morphological convergence in the radiations of Pacific Sebastes rockfishes. Am. Nat. 2014, 184, E115–E131. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.T.; Price, S.A.; Hoey, A.S.; Wainwright, P.C. Ecomorphological convergence in planktivorous surgeonfishes. J. Evol. Biol. 2016, 29, 965–978. [Google Scholar] [CrossRef] [PubMed]
- McKown, A.D.; Akamine, M.E.; Sack, L. Trait convergence and diversification arising from a complex evolutionary history in Hawaiian species of Scaevola. Oecologia 2016, 181, 1083–1100. [Google Scholar] [CrossRef] [PubMed]
- Losos, J.B. Convergence, adaptation, and constraint. Evolution 2011, 65, 1827–1840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumont, E.R.; Herrel, A.; Medellín, R.A.; Vargas-Contreras, J.A.; Santana, S.E. Built to bite: Cranial design and function in the wrinkle-faced bat. J. Zool. 2009, 279, 329–337. [Google Scholar] [CrossRef]
- Aguirre, L.F.; Herrel, A.; Van Damme, R.; Matthysen, E. The implications of food hardness for diet in bats. Funct. Ecol. 2003, 17, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Santana, S.E.; Dumont, E.R.; Davis, J.L. Mechanics of bite force production and its relationship to diet in bats. Funct. Ecol. 2010, 24, 776–784. [Google Scholar] [CrossRef] [Green Version]
- Dumont, E.R. Cranial shape in fruit, nectar, and exudate feeders: Implications for interpreting the fossil record. Am. J. Phys. Anthropol. 1997, 102, 187–202. [Google Scholar] [CrossRef]
- Santana, S.E.; Grosse, I.R.; Dumont, E.R. Dietary hardness, loading behavior, and the evolution of skull form in bats. Evolution 2012, 66, 2587–2598. [Google Scholar] [CrossRef] [PubMed]
- Rojas, D.; Vale, Á.; Ferrero, V.; Navarro, L. The role of frugivory in the diversification of bats in the Neotropics. J. Biogeogr. 2012, 39, 1948–1960. [Google Scholar] [CrossRef]
- Rossoni, D.M.; Assis, A.P.A.; Giannini, N.P.; Marroig, G. Intense natural selection preceded the invasion of new adaptive zones during the radiation of New World leaf-nosed bats. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gardner, A. Mammals of South. America, Volume 1: Marsupials, Xenarthrans, Shrews, and Bats; The University of Chicago Press: Chicago, IL, USA, 2008; ISBN 9780226282404. [Google Scholar]
- Rolland, J.; Condamine, F.L.; Jiguet, F.; Morlon, H. Faster speciation and reduced extinction in the tropics contribute to the mammalian latitudinal diversity gradient. PLoS Biol. 2014, 12, e1001775. [Google Scholar] [CrossRef] [PubMed]
- Morlon, H.; Lewitus, E.; Condamine, F.L.; Manceau, M.; Clavel, J.; Drury, J. RPANDA: An R package for macroevolutionary analyses on phylogenetic trees. Methods Ecol. Evol. 2016, 7, 589–597. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Hansen, T.F. Stabilizing selection and the comparative analysis of adaptation. Evolution 1997, 51, 1341–1351. [Google Scholar] [CrossRef] [PubMed]
- Khabbazian, M.; Kriebel, R.; Rohe, K.; Ané, C. Fast and accurate detection of evolutionary shifts in Ornstein-Uhlenbeck models. Methods Ecol. Evol. 2016, 7, 811–824. [Google Scholar] [CrossRef]
- Ingram, T.; Mahler, D.L. SURFACE: Detecting convergent evolution from comparative data by fitting Ornstein-Uhlenbeck models with stepwise Akaike Information Criterion. Methods Ecol. Evol. 2013, 4, 416–425. [Google Scholar] [CrossRef]
- Arbuckle, K.; Bennett, C.M.; Speed, M.P. A simple measure of the strength of convergent evolution. Methods Ecol. Evol. 2014, 5, 685–693. [Google Scholar] [CrossRef] [Green Version]
- Arbuckle, K.; Minter, A. Windex: Analyzing convergent evolution using the wheatsheaf index in R. Evol. Bioinforma. 2015, 11, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Herring, S.W.; Rafferty, K.L.; Liu, Z.J.; Marshall, C.D. Jaw muscles and the skull in mammals: The biomechanics of mastication. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 131, 207–219. [Google Scholar] [CrossRef]
- Cox, P.G. A quantitative analysis of the Eutherian orbit: Correlations with masticatory apparatus. Biol. Rev. 2008, 83, 35–69. [Google Scholar] [CrossRef] [PubMed]
- Pitnick, S.; Jones, K.E.; Wilkinson, G.S. Mating system and brain size in bats. Proc. R. Soc. B Biol. Sci. 2006, 273, 719–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellens, H.L.L.; Kuijpers-Jagtman, A.M.; Halazonetis, D.J. Geometric morphometric analysis of craniofacial variation, ontogeny and modularity in a cross-sectional sample of modern humans. J. Anat. 2013, 222, 397–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handley, C.O.; Wilson, D.E.; Gardner, A.L. Demography and Natural History of the Common Fruit Bat, Artibeus Jamaicensis, on Barro Colorado Island, Panamá; Smithsonian Institution Press: Washington, DC, USA, 1991; ISBN 0081-0282. [Google Scholar]
- Dumont, E.R. The correlated evolution of cranial morphology and feeding behavior in New World fruit bats. In Functional and Ecological Morphology of Bats; Kunz, G., McCraken, G., Akbar, Z., Eds.; Oxford University Press: New York, NY, USA, 2006; pp. 160–177. ISBN 139780195154726. [Google Scholar]
- Freeman, P.W. Form, function, and evolution in skulls and teeth of bats. In Bat Biology and Conservation; Kunz, T.H., Racey, P.A., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1998; pp. 140–156. ISBN 9781560988250. [Google Scholar]
- Schluter, D.; Nagel, L.M. Parallel speciation by natural selection. Am. Nat. 1995, 146, 292–301. [Google Scholar] [CrossRef]
- Muschick, M.; Indermaur, A.; Salzburger, W. Convergent evolution within an adaptive radiation of cichlid fishes. Curr. Biol. 2012, 22, 2362–2368. [Google Scholar] [CrossRef] [PubMed]
- Bonaccorso, F.J.; Winkelmann, J.R.; Shin, D.; Agrawal, C.I.; Aslami, N.; Bonney, C.; Hsu, A.; Jekielek, P.E.; Knox, A.K.; Kopach, S.J.; et al. Evidence for exploitative competition: Comparative foraging behavior and roosting ecology of short-tailed fruit bats (Phyllostomidae). Biotropica 2007, 39, 249–256. [Google Scholar] [CrossRef]
- Burbrink, F.T.; Pyron, R.A. How does ecological opportunity influence rates of speciation, extinction, and morphological diversification in New World ratsnakes (Tribe Lampropeltini)? Evolution 2009, 64, 934–943. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Trait | pPC1 | pPC2 | pPC3 | pPC4 | pPC5 |
|---|---|---|---|---|---|
| Jaw | |||||
| MDD | −0.850 | 0.519 | −0.041 | 0.049 | −0.066 |
| CPH | −0.990 | 0.112 | 0.045 | −0.013 | 0.071 |
| CH | −0.988 | −0.093 | 0.102 | 0.050 | −0.031 |
| CC | −0.997 | −0.041 | −0.064 | −0.013 | −0.006 |
| Cm1 | −0.996 | −0.043 | −0.058 | −0.035 | −0.006 |
| Cm3 | −0.988 | 0.021 | 0.039 | −0.142 | −0.034 |
| TDL | −0.995 | −0.065 | −0.073 | 0.004 | −0.029 |
| CCL | −0.998 | −0.014 | 0.019 | −0.012 | −0.001 |
| CAL | −0.995 | 0.016 | 0.084 | 0.014 | 0.019 |
| MAN | −0.989 | −0.043 | −0.129 | 0.042 | 0.024 |
| Skull | |||||
| TSL | −0.980 | −0.171 | −0.094 | 0.002 | −0.002 |
| MSW | −0.904 | −0.353 | −0.189 | −0.009 | −0.041 |
| ZB | −0.667 | −0.718 | −0.131 | −0.077 | −0.106 |
| PSW | −0.784 | −0.564 | −0.249 | −0.016 | −0.015 |
| PW | 0.118 | −0.988 | 0.077 | 0.069 | −0.010 |
| PW1 | −0.006 | −0.981 | 0.097 | −0.153 | 0.068 |
| TPL | −0.986 | 0.148 | 0.074 | 0.024 | 0.010 |
| ASL | −0.994 | −0.084 | −0.048 | 0.023 | 0.046 |
| PPL | −0.192 | 0.772 | −0.577 | 0.106 | 0.134 |
| MTL | −0.995 | −0.070 | 0.017 | −0.039 | −0.020 |
| Regimes | Phenotype | w | Lower Bound | Upper Bound | p-Value |
|---|---|---|---|---|---|
| 1 | pPC1 | 3.40 | 3.27 | 4.19 | 0.02 |
| pPC2 | 2.69 | 2.52 | 3.84 | 0.08 | |
| pPC3 | 4.08 | 3.96 | 5.44 | 0.04 | |
| pPC4 | 3.98 | 3.44 | 6.98 | 0.02 | |
| pPC5 | 3.65 | 3.51 | 5.04 | 0.06 | |
| 2 | pPC1 | 2.92 | 2.77 | 3.24 | <0.01 |
| pPC2 | 2.12 | 1.98 | 2.72 | 0.03 | |
| pPC3 | 4.84 | 4.59 | 6.10 | <0.01 | |
| pPC4 | 1.69 | 1.56 | 2.14 | 0.11 | |
| pPC5 | 1.73 | 1.62 | 1.92 | 0.14 | |
| 3 | pPC1 | 8.58 | 8.29 | 8.72 | 0.14 |
| pPC2 | 0.68 | 0.65 | 0.69 | 0.96 | |
| pPC3 | 3.37 | 3.27 | 3.41 | 0.36 | |
| pPC4 | 5.50 | 5.33 | 5.60 | 0.19 | |
| pPC5 | 10.28 | 9.87 | 10.45 | 0.20 | |
| 4 | pPC1 | 1.02 | 0.97 | 1.29 | 0.14 |
| pPC2 | 1.62 | 1.55 | 1.93 | 0.08 | |
| pPC3 | 1.81 | 1.72 | 2.01 | 0.01 | |
| pPC4 | 2.91 | 2.77 | 3.42 | <0.01 | |
| pPC5 | 1.31 | 1.26 | 1.57 | 0.17 |
| Regimes | Phenotype | w | Lower Bound | Upper Bound | p-Value |
|---|---|---|---|---|---|
| 1 | pPC1 | 2.57 | 2.42 | 3.98 | 0.06 |
| pPC2 | 4.95 | 4.71 | 5.86 | 0.03 | |
| pPC3 | 2.04 | 1.96 | 2.56 | 0.12 | |
| pPC4 | 2.37 | 2.27 | 4.10 | 0.12 | |
| pPC5 | 4.14 | 4.01 | 7.27 | 0.03 | |
| 2 | pPC1 | 1.90 | 1.79 | 2.57 | 0.03 |
| pPC2 | 5.08 | 4.76 | 5.69 | 0.01 | |
| pPC3 | 2.24 | 2.11 | 2.28 | 0.03 | |
| pPC4 | 1.57 | 1.45 | 2.01 | 0.05 | |
| pPC5 | 1.00 | 0.96 | 1.06 | 0.44 | |
| 3 | pPC1 | 0.99 | 0.93 | 1.15 | 0.18 |
| pPC2 | 0.91 | 0.87 | 1.04 | 0.35 | |
| pPC3 | 2.94 | 2.79 | 3.56 | <0.01 | |
| pPC4 | 2.29 | 2.03 | 2.56 | 0.01 | |
| pPC5 | 1.30 | 1.24 | 1.49 | 0.05 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murillo-García, O.E.; De la vega, M.E. Divergence, Convergence and Phenotypic Diversity of Neotropical Frugivorous Bats. Diversity 2018, 10, 100. https://doi.org/10.3390/d10030100
Murillo-García OE, De la vega ME. Divergence, Convergence and Phenotypic Diversity of Neotropical Frugivorous Bats. Diversity. 2018; 10(3):100. https://doi.org/10.3390/d10030100
Chicago/Turabian StyleMurillo-García, Oscar E., and Maria E. De la vega. 2018. "Divergence, Convergence and Phenotypic Diversity of Neotropical Frugivorous Bats" Diversity 10, no. 3: 100. https://doi.org/10.3390/d10030100
APA StyleMurillo-García, O. E., & De la vega, M. E. (2018). Divergence, Convergence and Phenotypic Diversity of Neotropical Frugivorous Bats. Diversity, 10(3), 100. https://doi.org/10.3390/d10030100