Individual Morphology and Habitat Structure Alter Social Interactions in a Range-Shifting Species


Abstract
:1. Introduction
2. Methods
2.1. Crab Collection and Preparation
2.2. Aggression Assays
2.3. Claw Morphology
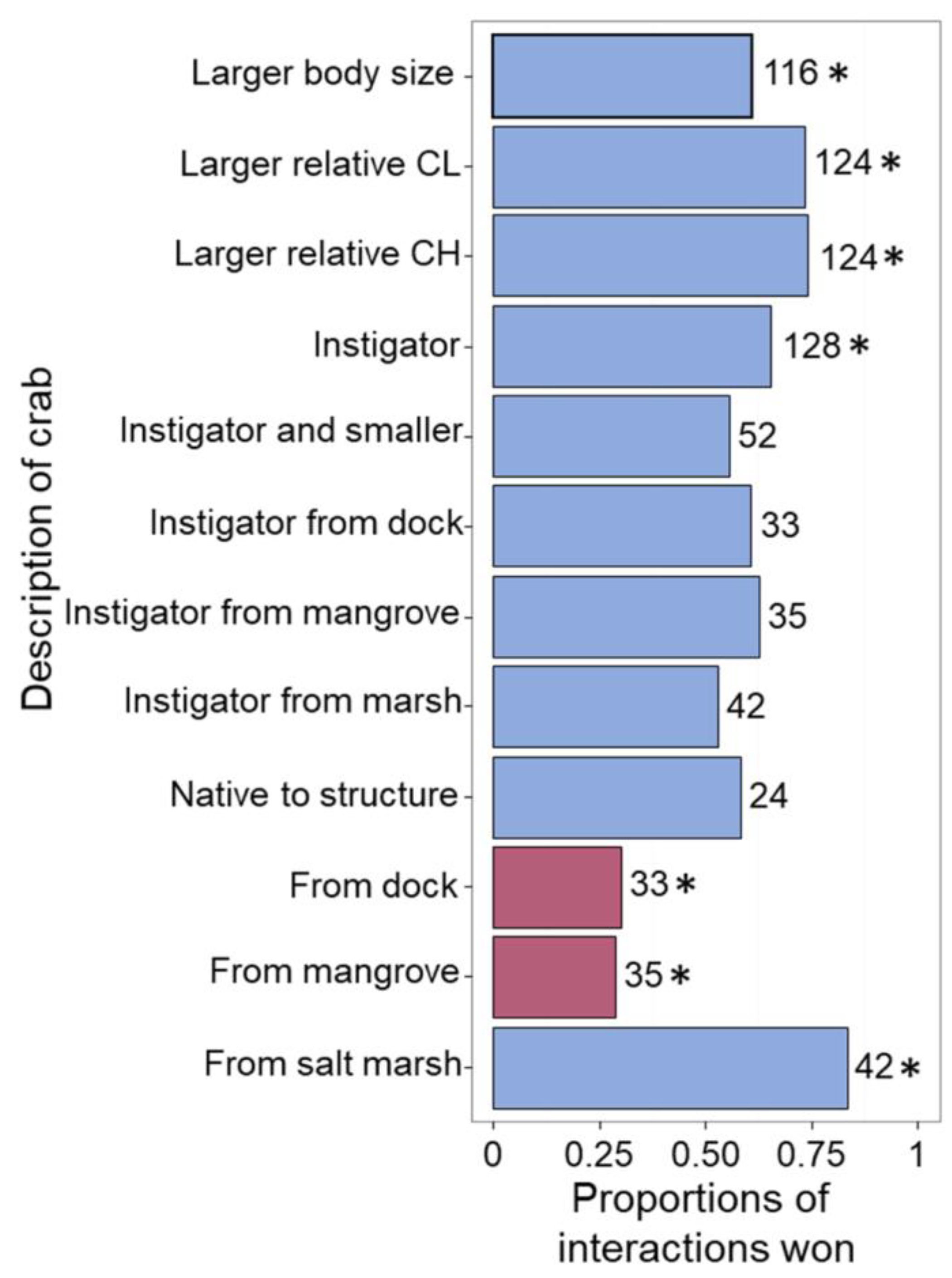
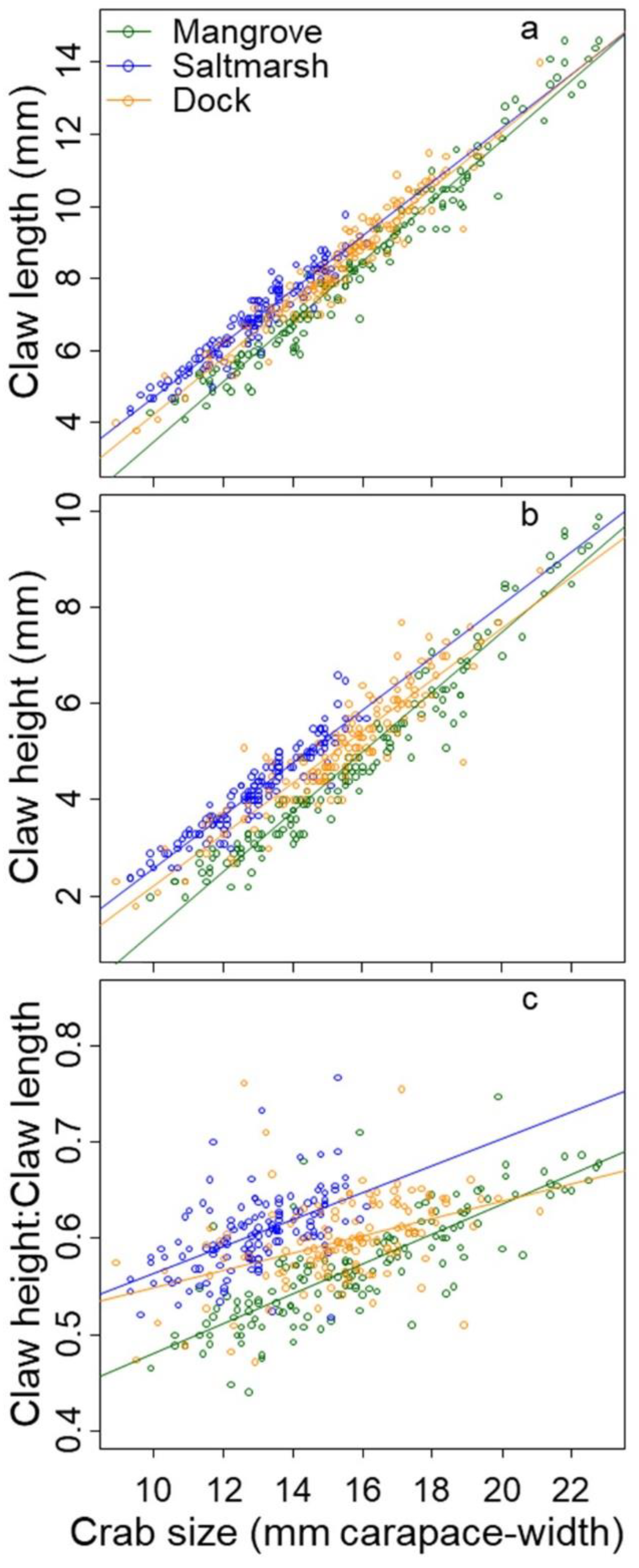
3. Results
3.1. Aggression Assays
3.2. Claw Morphology
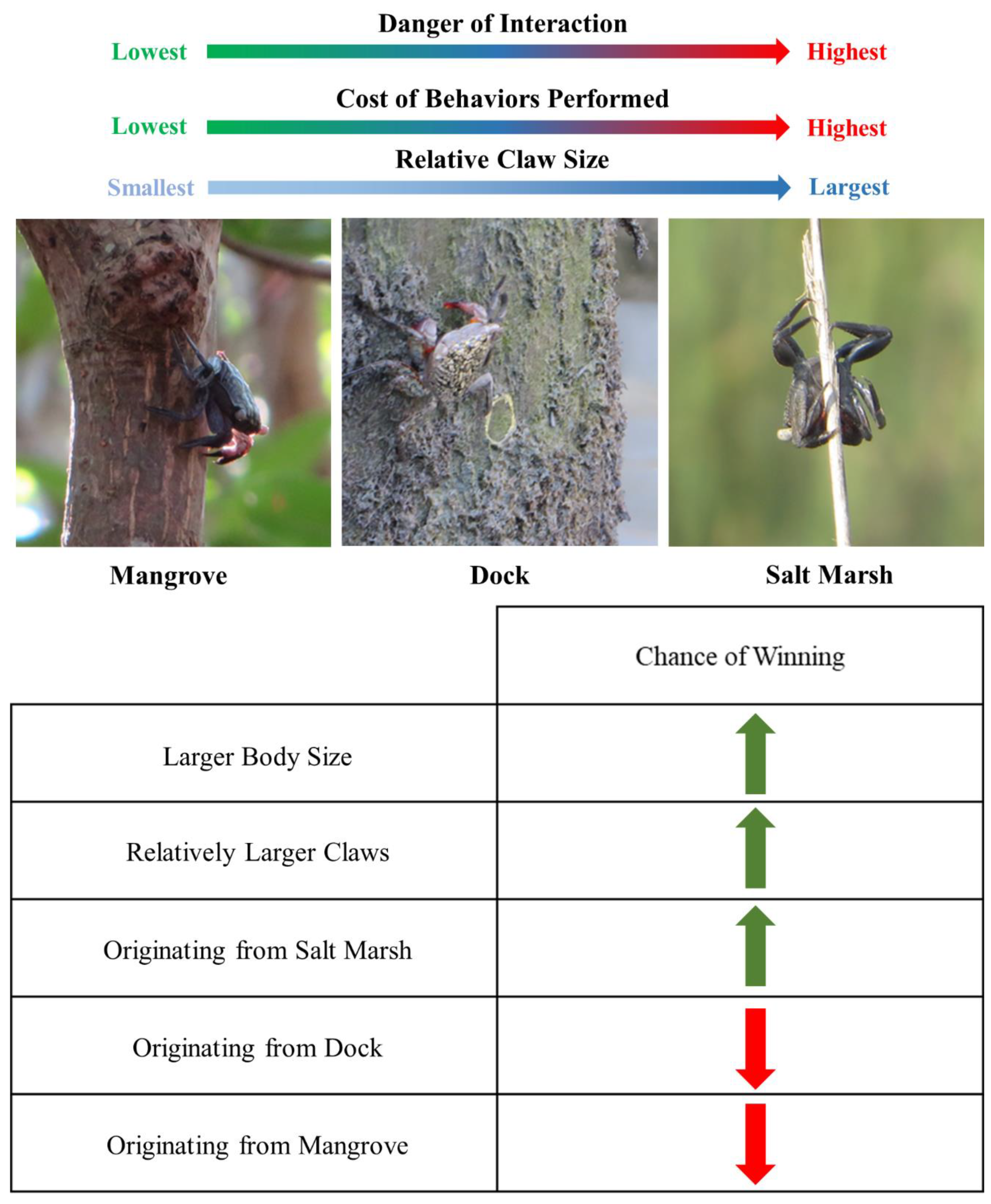
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Dayton, P.K. Toward an understanding of community resilience and the potential effects of enrichments to the benthos at McMurdo Sound, Antarctica. In Proceeding of the Colloquium on the Conservation Problems in Antarctica; Parker, B.C., Ed.; Allen Press: Lawrence, KS, USA, 1972; pp. 81–96. [Google Scholar]
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliot, K.; Webster, J.R. Loss of foundation species: Consequences for the structure and dynamics of forested ecosystems. Front. Ecol. Environ. 2005, 3, 479–486. [Google Scholar] [CrossRef]
- Cannizzo, Z.J.; Griffen, B.D. Changes in behaviour patterns by mangrove tree crabs following climate-induced range shift into novel habitat. Anim. Behav. 2016, 121, 79–86. [Google Scholar] [CrossRef]
- Bruschetti, M.; Bazterrica, C.; Luppi, T.; Iribarne, O. An invasive intertidal reef-forming polychaete affect habitat use and feeding behavior of migratory and local birds in a SW Atlantic coastal lagoon. J. Exp. Mar. Biol. Ecol. 2009, 375, 76–83. [Google Scholar] [CrossRef]
- Jayadevan, A.; Mukherjee, S.; Vanak, A.T. Bush encroachment influences nocturnal rodent community behaviour in a semi-arid grassland in Grujarat, India. J. Arid Environ. 2018, 153, 32–38. [Google Scholar] [CrossRef]
- Gribben, P.E.; Wright, J.T. Habitat-former effects on prey behaviour increase predation and non-predation mortality. J. Anim. Ecol. 2014, 83, 388–396. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, B.S.; Mello, K.; Litterer, A.; Dijkstra, J.A. Seaweed structure shapes trophic interactions: A case study using a mid-trophic level fish species. J. Exp. Mar. Biol. Ecol. 2018, 506, 1–8. [Google Scholar] [CrossRef]
- Schweiger, O.; Settle, J.; Kudrna, O. Climate change can cause spatial mismatch of trophically interacting species. Ecology 2008, 89, 3472–3479. [Google Scholar] [CrossRef] [PubMed]
- Walther, G.R. Community and ecosystem responses to recent climate change. Philos. Trans. R. Soc. B. Biol. Sci. 2010, 365, 2019–2024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, B.B.M.; Candolin, U. Behavioral responses to changing environments. Behav. Ecol. 2015, 26, 665–673. [Google Scholar] [CrossRef]
- Riley, M.E.; Johnston, C.A.; Feller, I.C.; Griffen, B.D. Range expansion of Aratus pisonii (mangrove tree crab) into novel vegetative habitats. Southeast. Nat. 2014, 13, N43–N48. [Google Scholar] [CrossRef]
- Riley, M.E.; Griffen, B.D. Habitat-specific differences alter traditional biogeographic patterns of life history in a climate-change induced range expansion. PLoS ONE 2017, 12, e0176263. [Google Scholar] [CrossRef] [PubMed]
- Cannizzo, Z.J.; Dixon, S.R.; Griffen, B.D. An anthropogenic habitat within a suboptimal colonized ecosystem provides improved conditions for a range-shifting species. Ecol. Evol. 2018, 8, 1524–1533. [Google Scholar] [CrossRef] [PubMed]
- Enquist, M.; Leimar, O. Evolution of fighting behaviour: Decision rules and assessment of relative strength. J. Theor. Biol. 1983, 102, 387–410. [Google Scholar] [CrossRef]
- Warner, G.F. Behaviour of two species of grapsid crab during intraspecific encounters. Behaviour 1970, 36, 9–19. [Google Scholar] [CrossRef]
- Greene, M.J.; Mason, R.T. Courtship, mating, and male combat of the brown tree snake, Boiga irregularis. Herpetologica 2000, 56, 166–175. [Google Scholar]
- Slater, K.Y.; Schaffner, C.M.; Aureli, F. Female-directed male aggression in wild Ateles geoffroyi ssp. yucatanensis. Int. J. Primatol. 2008, 29, 1657–1669. [Google Scholar] [CrossRef]
- Theil, M.; Lovrich, G.A. Agonistic behaviour and reproductive biology of squat lobsters. In Biology of Squat Lobsters; Poore, G.C.B., Ahyong, S.T., Taylor, J., Eds.; Csiro Publishing: Clayton, Vic, Australia, 2011; pp. 233–247. [Google Scholar]
- Schofield, G.; Katselidis, K.A.; Pantis, J.D.; Dimopoulos, P.; Hays, G.C. Female-female aggression: Structure of interaction and outcome in loggerhead sea turtles. Mar. Ecol. Prog. Ser. 2007, 336, 267–274. [Google Scholar] [CrossRef]
- Fernandez, A.A.; Fasel, N.; Knörnschild, M.; Richner, H. When bats are boxing: Aggressive behaviour and communication in male Seba’short-tailes fruit bat. Anim. Behav. 2014, 98, 149–156. [Google Scholar] [CrossRef]
- Mercier, J.L.; Lenoir, A.; Dejean, A. Ritualised versus aggressive beahviours displayed by Polrhachis laboriosa (F. Smith) during intraspecific competition. Behav. Process. 1997, 41, 36–50. [Google Scholar] [CrossRef]
- Skaggs, R.; Jackson, J.C.; Toth, A.L.; Schneider, S.S. The possible role of ritualized aggression in the vibration signal of the honeybee, Apis mellifera. Anim. Behav. 2014, 98, 103–111. [Google Scholar] [CrossRef]
- Trisko, R.K.; Smuts, B.B. Dominance relationships in a group of domestic dogs (Canis lupus famililaris). Behaviour 2015, 152, 677–704. [Google Scholar] [CrossRef]
- Wilczynski, W.; Black, M.P.; Salem, S.J.; Ezeoke, C.B. Behavioural persistence during an agonistic encounter differentiates winners from losers in green anole lizards. Behaviour 2015, 152, 563–591. [Google Scholar] [CrossRef]
- Warner, G.F. The life history of the Mangrove Tree Crab, Aratus pisonii. J. Zool. 1967, 153, 321–335. [Google Scholar] [CrossRef]
- Sneddon, L.U.; Huntingford, F.A.; Taylor, A.C. Weapon size versus body size as a predictor of winning in fights between shore crabs, Carcinus maenas (L.). Behav. Ecol. Sociobiol. 1997, 41, 237–242. [Google Scholar] [CrossRef]
- Brian, J.V.; Fernandes, T.; Ladle, R.J.; Todd, P.A. Patterns of morphological and genetic variability in UK populations of the shore crab, Carcius maenas Linnaeus, 1758 (Crustacea: Decapoda: Brachyura). J. Exp. Mar. Biol. Ecol. 2006, 329, 47–54. [Google Scholar] [CrossRef]
- Smith, L.D.; Palmer, A.R. Effects of manipulated diet on size and performance of bracyuran crab claws. Science 1994, 264, 710–712. [Google Scholar] [CrossRef]
- De Lemos Santana, J.; Dos Santos Calado, T.C.; de Alemeids Alves-Júnior, F.; de Oliveira, M.A.; de Sá Leitão Câmara de Araújo, M. Populational structure and sexual maturity of Aratus pisonii (H. Milne Edwarrds, 1837) (Crustacea, Decapoda, Sesarmidae) in the estuarine channels of Mundaú Lagoon, Northeastern Brazil. Panam. J. Aquat. Sci. 2018, 13, 1–12. [Google Scholar]
- Neat, F.C.; Huntingford, F.A.; Beveridge, M.M.C. Fighting and assessmet in male sichlid fish: The effects of asymmetries in gonadal state and body size. Anim. Behav. 1998, 55, 883–891. [Google Scholar] [CrossRef]
- Hofmann, H.A.; Schildberger, K. Assessment of strength and wiliness to fight during aggressive encounters in crickets. Anim. Behav. 2001, 62, 337–348. [Google Scholar] [CrossRef]
- Johnston, C.A.; Smith, R.S. Vegetation structure mediates a shift in predator avoidance behavior in a range-edge population. Behav. Ecol. 2018, 29, 1124–1131. [Google Scholar] [CrossRef]
- Huxley, J.A. A discussion on ritualization of beahviour in animals and man. R. Soc. 1966, 4, 1–60. [Google Scholar]
- Caldwell, R.L.; Dingle, J. The influence of size differential on agonistic encounters in the mantis shrimp Gonodactylus viridis. Behaviour 1979, 69, 255–264. [Google Scholar] [CrossRef]
- Gabbanini, F.; Gherardi, F.; Vannini, M. Force and dominance in the agonistic behavior of the freshwater crab Potamon fluviatile. Aggress. Behav. 1995, 21, 451–462. [Google Scholar] [CrossRef]
- Parker, G.A. Assessment strategy and the evolution of fighting behaviour. J. Theor. Biol. 1974, 74, 223–243. [Google Scholar] [CrossRef]
- Smith, I.P.; Huntingford, F.A.; Atkinson, R.J.A.; Taylor, A.C. Strategic decisions during anostic behaviour in the velvet swimming crab, Necora puber (L.). Anim. Behav. 1994, 47, 885–894. [Google Scholar] [CrossRef]
- Maynard Smith, J.; Parker, G.A. The logic of asymmetric contests. Anim. Behav. 1976, 24, 159–175. [Google Scholar] [CrossRef]
- Poole, J.H. Announcing intent: The aggressive state of musth in African elephants. Anim. Behav. 1989, 37, 140–152. [Google Scholar] [CrossRef]
- Arnott, G.; Elwood, R.W. Assessment of fighting ability in animal contests. Anim. Behav. 2009, 77, 991–1004. [Google Scholar] [CrossRef]
- Payne, R.J.H. Gradually escalating fights and displays: The cumulative assessment model. Anim. Behav. 1998, 56, 651–662. [Google Scholar] [CrossRef]
- Enquist, M.; Leimar, O.; Ljungberg, T.; Mallner, Y.; Segerdahl, N. A test of the sequential assessment game—Fighting in the cichlid fish Nannacara anomala. Anim. Behav. 1990, 40, 1–14. [Google Scholar] [CrossRef]
- Taylor, P.W.; Elwood, R.W. The mismeasure of animal contests. Anim. Behav. 2003, 65, 1195–1202. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat | Site | Lat-Long |
|---|---|---|
| Mangrove | Pepper Park | 27°29′42″ N 80°18′12″ W |
| Mangrove | Round Island Park | 27°33′33″ N 80°19′53″ W |
| Mangrove | Oslo Road | 27°35′14″ N 80°21′55″ W |
| Mangrove | North Causeway | 27°28′28″ N 80°19′12″ W |
| Salt marsh | GTM NERR | 30°0′49″ N 81°20′42″ W |
| Salt marsh | Anastasia State Park | 29°52′40″ N 81°16′32″ W |
| Salt marsh | Vilano Marsh | 29°55′16″ N 81°17′57″ W |
| Dock | Palm Valley | 30°07′57″ N 81°23′08″ W |
| Dock | Yacht Club | 29°53′09″ N 81°17′08″ W |
| Dock | Vilano Dock | 29°56′33″ N 81°18′32″ W |
| Behavior | Description | Progression Order | Cost-Ranking (low to high) |
|---|---|---|---|
| Shield Posture | Body held high off the substrate with claws pointed down in a shield-like manner perpendicular to structure | 1 | 1 |
| Pre-display | Vigorous vibration of claws followed by raising of claws from shield position in an arc above the crab and back to shield posture. May be repeated many times | 2 | 2 |
| Fighting | Opponents grasp chelae and push each other. A fight was considered to begin when claws touched and end when claws separated | 3 | 3 |
| Post-display | Display behavior performed by winner of interaction. May occur with or independently from chasing behavior | 4 | -- |
| Chasing | Following a retreating opponent; performed by winning competitor | 4 | -- |
| Retreating | The purposeful backing away from an opponent; performed by losing competitor | 4 | -- |
| Measure of Claw Size | Habitat | Relationship to Body Size | Adjusted r2 |
|---|---|---|---|
| Claw Length (CL) | Mangrove | CL = 0.832 × body_size − 4.817 | 0.964 |
| Salt Marsh | CL = 0.745 × body_size − 2.717 | 0.916 | |
| Dock | CL = 0.781 × body_size − 3.536 | 0.928 | |
| Claw Height (CH) | Mangrove | CH = 0.620 × body_size − 4.932 | 0.947 |
| Salt Marsh | CH = 0.545 × body_size − 2.841 | 0.890 | |
| Dock | CH = 0.534 × body_size − 3.123 | 0.836 | |
| Claw Height-Claw Length Ratio (CH:CL) | Mangrove | CH:CL = 0.015 × body_size + 0.327 | 0.660 |
| Salt Marsh | CH:CL = 0.014 × body_size + 0.424 | 0.288 | |
| Dock | CH:CL = 0.009 × body_size + 0.458 | 0.173 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cannizzo, Z.J.; Nix, S.K.; Whaling, I.C.; Griffen, B.D. Individual Morphology and Habitat Structure Alter Social Interactions in a Range-Shifting Species. Diversity 2019, 11, 6. https://doi.org/10.3390/d11010006
Cannizzo ZJ, Nix SK, Whaling IC, Griffen BD. Individual Morphology and Habitat Structure Alter Social Interactions in a Range-Shifting Species. Diversity. 2019; 11(1):6. https://doi.org/10.3390/d11010006
Chicago/Turabian StyleCannizzo, Zachary J., Sara K. Nix, Isabel C. Whaling, and Blaine D. Griffen. 2019. "Individual Morphology and Habitat Structure Alter Social Interactions in a Range-Shifting Species" Diversity 11, no. 1: 6. https://doi.org/10.3390/d11010006
APA StyleCannizzo, Z. J., Nix, S. K., Whaling, I. C., & Griffen, B. D. (2019). Individual Morphology and Habitat Structure Alter Social Interactions in a Range-Shifting Species. Diversity, 11(1), 6. https://doi.org/10.3390/d11010006