Could Hair-Lichens of High-Elevation Forests Help Detect the Impact of Global Change in the Alps?

 ,
,  , ,
, , {kind=link}
Abstract
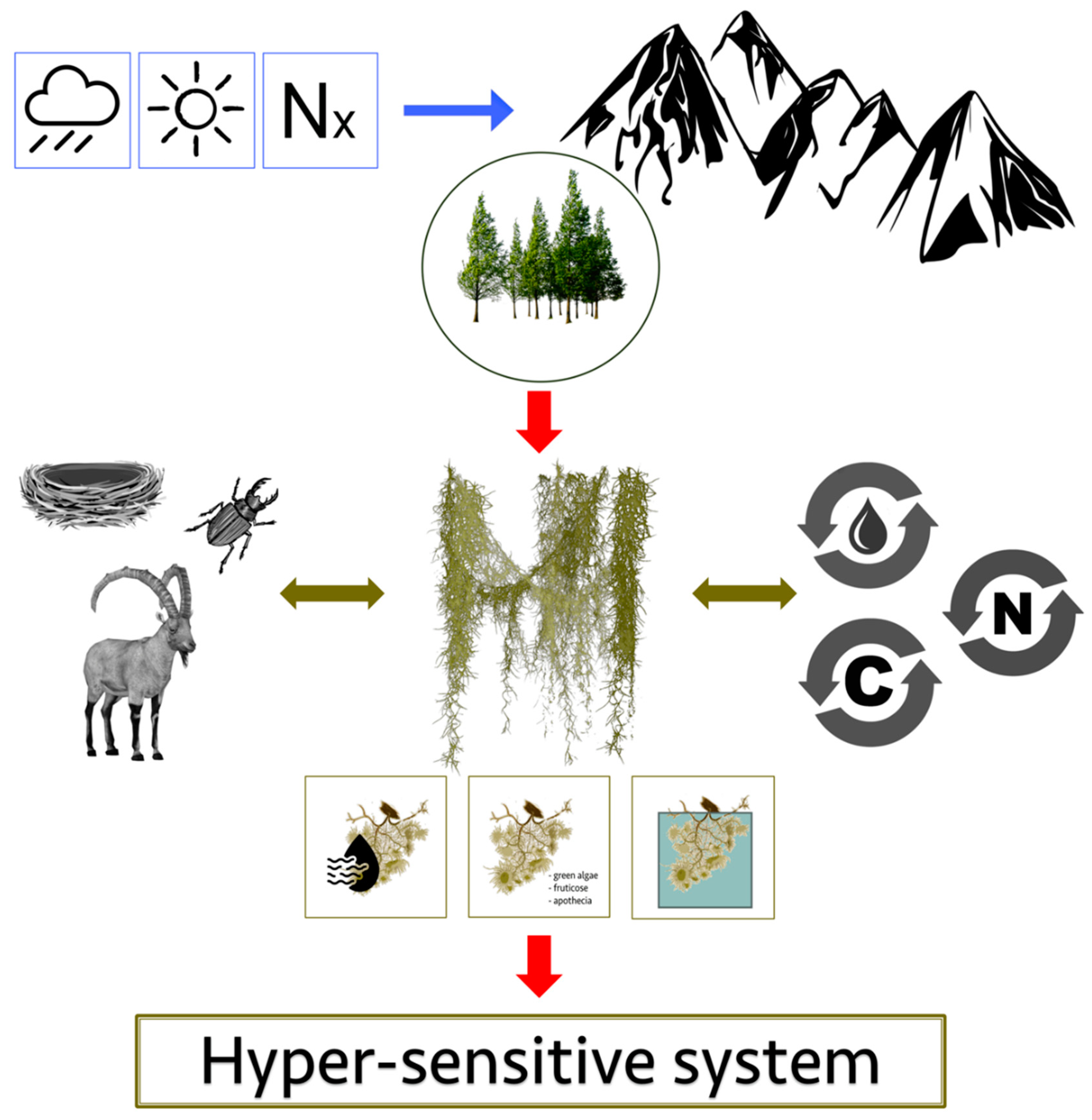
:1. Introduction
2. Main Patterns of Climate Change and Nitrogen Pollution in the Alps
3. Hair-Lichens
4. Lichen–Climate Relationships with Emphasis on Hair-Lichens
5. Lichen–Nitrogen Pollution Relationships with Emphasis on Hair-Lichens
6. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moore, F.C.; Diaz, D.B. Temperature impacts on economic growth warrant stringent mitigation policy. Nat. Clim. Chang. 2015, 5, 127–131. [Google Scholar] [CrossRef]
- Rands, M.R.W.; Adams, W.M.; Bennun, L.; Butchart, S.H.M.; Clements, A.; Coomes, D.; Entwistle, A.; Hodge, I.; Kapos, V.; Scharlemann, J.P.W.; et al. Biodiversity conservation: Challenges beyond 2010. Science 2010, 329, 1298–1303. [Google Scholar] [CrossRef] [PubMed]
- Pepin, N.; Bradley, R.S.; Diaz, H.F.; Baraer, M.; Caceres, E.B.; Forsythe, N.; Fowler, H.; Greenwood, G.; Hashmi, M.Z.; Liu, X.D.; et al. Elevation-dependent warming in mountain regions of the world. Nat. Clim. Chang. 2015, 5, 424–430. [Google Scholar] [Green Version]
- EEA. Regional Climate Change and Adaptation: The Alps Facing the Challenge of Changing Water Resources; EEA: Copenhagen, Denmark, 2009; Volume 8, 143p. [Google Scholar]
- Cristofolini, F.; Brunialti, G.; Giordani, P.; Nascimbene, J.; Cristofori, A.; Gottardini, E.; Frati, L.; Matos, P.; Batič, F.; Caporale, S.; et al. Towards the adoption of an international standard for biomonitoring with lichens—Consistency of assessment performed by experts from six European countries. Ecol. Indic. 2014, 45, 63–67. [Google Scholar] [CrossRef]
- Giordani, P.; Brunialti, G.; Bacaro, G.; Nascimbene, J. Functional traits of epiphytic lichens as potential indicators of environmental conditions in forest ecosystems. Ecol. Indic. 2012, 18, 413–420. [Google Scholar] [CrossRef]
- Bobbink, R.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Bustamante, M.; Cinderby, S.; Davidson, E.; Dentener, F.; et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis. Ecol. Appl. 2010, 20, 30–59. [Google Scholar] [CrossRef]
- Dise, N.B.; Ashmore, M.; Belyazid, S.; Bleeker, A.; Bobbink, R.; De Vries, W.; Erisman, J.W.; Spranger, T.; Stevens, C.J.; Van der Berg, L. Nitrogen as a threat to European terrestrial biodiversity. In The European Nitrogen Assessment; Sutton, M.A., Howard, C.M., Erisman, J.W., Billen, G., Bleeker, A., Grennfelt, P., Van Grinsven, H., Grizzetti, B., Eds.; Cambridge University Press, United States of America: New York, NY, USA, 2011; pp. 463–494. [Google Scholar]
- Rogora, M.; Mosello, R.; Arisci, S.; Brizzio, M.C.; Barbieri, A.; Balestrini, R.; Waldner, P.; Schmitt, M.; Stähli, M.; Thimonier, A.; et al. An overview of atmospheric deposition chemistry over the Alps: Present status and long-term trends. Hydrobiologia 2006, 562, 17–40. [Google Scholar] [CrossRef]
- Gobiet, A.; Kotlarski, S.; Beniston, M.; Heinrich, G.; Rajczak, J.; Stoffel, M. 21st century climate change in the European Alps—A review. Sci. Total Environ. 2014, 493, 1138–1151. [Google Scholar] [CrossRef] [Green Version]
- Auer, I.; Böhm, R.; Jurkovic, A.; Lipa, W.; Orlik, A.; Potzmann, R.; Schöner, W.; Ungersböck, M.; Matulla, C.; Briffa, K.; et al. HISTALP-historical instrumental climatological surface time series of the Greater Alpine Region. Int. J. Climatol. 2007, 27, 17–46. [Google Scholar] [CrossRef]
- Isotta, F.A.; Frei, C.; Weilguni, V.; Perčec Tadić, M.; Lassègues, P.; Rudolf, B.; Pavan, V.; Cacciamani, C.; Antolini, G.; Ratto, S.M.; et al. The climate of daily precipitation in the Alps: Development and analysis of a high-resolution grid dataset from pan-Alpine rain-gauge data. Int. J. Climatol. 2014, 34, 1657–1675. [Google Scholar] [CrossRef]
- Peel, J.L.; Haeuber, R.; Garcia, V.; Russell, A.G.; Neas, L. Impact of nitrogen and climate change interactions on ambient air pollution and human health. Biogeochemistry 2013, 114, 121–134. [Google Scholar] [CrossRef]
- Kaiser, A. Origin of polluted air masses in the Alps. An overview and first results for MONARPOP. Environ. Pollut. 2009, 157, 3232–3237. [Google Scholar] [CrossRef]
- Humbert, J.-Y.; Dwyer, J.M.; Andrey, A.; Arlettaz, R. Impacts of nitrogen addition on plant biodiversity in mountain grasslands depend on dose, application duration and climate: A systematic review. Glob. Chang. Biol. 2015, 22, 110–120. [Google Scholar] [CrossRef]
- Hämmerlee, A.I.; Wessely, J.; Baatar, U.; Essl, F.; Moser, D.; Jiménez-Alfaro, B.; Jandt, U.; Agrillo, E.; Stancić, Z.; Dirnböck, T.; et al. A new method for jointly assessing effects of climate change and nitrogen deposition on habitats. Biol. Conserv. 2018, 228, 52–61. [Google Scholar] [CrossRef]
- Zhao, Z.; Dong, S.; Jiang, X.; Liu, S.; Ji, H.; Li, Y.; Han, Y.; Sha, W. Effects of warming and nitrogen deposition on CH4, CO2 and N2O emissions in alpine grassland ecosystems of the Qinghai-Tibetan Plateau. Sci. Total Environ. 2017, 592, 565–572. [Google Scholar] [CrossRef]
- Root, H.T.; Geiser, L.H.; Jovan, S.; Neitlich, P. Epiphytic macrolichen indication of air quality and climate in interiorforested mountains of the Pacific Northwest, USA. Ecol. Indic. 2015, 53, 95–105. [Google Scholar] [CrossRef]
- Boutin, M.; Corcket, E.; Alard, D.; Villar, L.; Jiménez, J.-J.; Blaix, C.; Lemaire, C.; Corriol, G.; Lamaze, T.; Pornon, A. Nitrogen deposition and climate change have increased vascular plant species richness and altered the composition of grazed subalpine grasslands. J. Ecol. 2017, 105, 1199–1209. [Google Scholar] [CrossRef]
- Kirchner, M.; Fegg, W.; Römmelt, H.; Leuchner, M.; Ries, L.; Zimmermann, R.; Michalke, B.; Wallasch, M.; Maguhn, J.; Faus-Kessler, T.; et al. Nitrogen deposition along differently exposed slopes in the Bavarian Alps. Sci. Total Environ. 2014, 470–471, 895–906. [Google Scholar] [CrossRef]
- Kinley, T.A.; Goward, T.; McLellan, B.N.; Serrouya, R. The influence of variable snowpacks on habitat use by mountain caribou. Rangifer 2006, 17, 93–102. [Google Scholar] [CrossRef]
- Kivinen, S.; Moen, J.; Berg, A.; Eriksson, A. Effects of modern forest management on winter grazing resources for reindeer in Sweden. Ambio 2010, 39, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, R.B.; Ball, J.P.; Renhorn, K.E.; Esseen, P.-A.; Sjöberg, K. Invertebrate communities in boreal forest canopies as influenced by forestry and lichens with implications for passerine birds. Biol. Conserv. 1995, 74, 57–63. [Google Scholar] [CrossRef]
- Campbell, J.; Coxson, D.S. Canopy microclimate and arboreal lichen loading in subalpine spruce-fir forest. Can. J. Bot. 2001, 79, 537–555. [Google Scholar]
- Gauslaa, Y. Rain, dew, and humid air as drivers of morphology, function and spatial distribution in epiphytic lichens. Lichenologist 2014, 46, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ellis, C.J.; Eaton, S.; Theodoropoulos, M.; Coppins, B.J.; Seaward, M.R.D.; Simkin, J. Response of epiphytic lichens to 21st Century climate change and tree disease scenarios. Biol. Conserv. 2014, 180, 153–164. [Google Scholar] [CrossRef]
- Giordani, P.; Incerti, G. The influence of climate on the distribution of lichens: A case study in a borderline area (Liguria, NW Italy). Plant Ecol. 2008, 195, 257–272. [Google Scholar] [CrossRef]
- Marini, L.; Nascimbene, J.; Nimis, P.L. Large-scale patterns of epiphytic lichen species richness: Photobiont-dependent response to climate and forest structure. Sci. Total Environ. 2011, 409, 4381–4386. [Google Scholar] [CrossRef] [PubMed]
- Aragón, G.; Martínez, I.; García, A. Loss of epiphytic diversity along a latitudinal gradient in southern Europe. Sci. Total Environ. 2012, 426, 188–195. [Google Scholar] [CrossRef]
- Elbert, W.; Weber, B.; Burrows, S.; Steinkamp, J.; Büdel, B.; Andreae, M.O.; Pöschl, U. Contribution of cryptogamic covers to the global cycles of carbon and nitrogen. Nat. Geosci. 2012, 5, 459–462. [Google Scholar] [CrossRef]
- Porada, P.; Weber, B.; Elbert, W.; Pöschl, U.; Kleidon, A. Estimating global carbon uptake by lichens and bryophytes with a process-based model. Biogeosciences 2013, 10, 6989–7033. [Google Scholar] [CrossRef] [Green Version]
- Porada, P.; Van Stan, J.T., II; Kleidon, A. Significant contribution of non-vascular vegetation to global rainfall interception. Nat. Geosci. 2018, 11, 563–567. [Google Scholar] [CrossRef]
- Insarov, G.; Schroeter, B. Lichen monitoring and climate change. In Monitoring with Lichens, Monitoring Lichens; Nimis, P.L., Scheidegger, C., Wolseley, P., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 183–201. [Google Scholar]
- Green, T.G.A.; Nash, T.H., III; Lange, O.L. Physiological ecology of carbon dioxide exchange. In Lichen Biology; Nash, T.H., III, Ed.; Cambridge University Press: Cambridge, UK, 2008; pp. 152–181. [Google Scholar]
- Schroeter, B.; Kappen, L.; Schulz, F.; Sancho, L.G. Seasonal variation in the carbon balance of lichens in the maritime Antarctic: Long-term measurements of photosynthetic activity in Usnea aurantiaco-atra. In Antarctic Ecosystems: Models for Wider Ecological Understanding; Davison, W., Howard-Williams, C., Broady, P., Eds.; Caxton Press: Christchurch, New Zealand, 2000; pp. 258–262. [Google Scholar]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.-F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, water, and broad-scale geographic patterns of species richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef]
- Nascimbene, J.; Nimis, P.L.; Dainese, M. Epiphytic lichen conservation in the Italian Alps: The role of forest type. Fungal Ecol. 2014, 11, 164–172. [Google Scholar] [CrossRef]
- Diaz, S.; Cabido, M. Vive la difference: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Nascimbene, J.; Marini, L. Epiphytic lichen diversity along elevational gradients: Biological traits reveal a complex response to water and Energy. J. Biogeogr. 2015, 42, 1222–1232. [Google Scholar] [CrossRef]
- Ellis, C.J.; Coppins, B.J. Contrasting functional traits maintain lichen epiphyte diversity in response to climate and autogenic succession. J. Biogeogr. 2006, 33, 1643–1656. [Google Scholar] [CrossRef]
- Esseen, P.-A.; Ekström, M.; Westerlund, B.; Palmqvist, K.; Jonsson, B.G.; Grafström, A.; Ståhl, G. Broad-scale distribution of epiphytic hair lichens correlates more with climate and nitrogen deposition than with forest structure. Can. J. For. Res. 2016, 46, 1348–1358. [Google Scholar] [CrossRef] [Green Version]
- Coxson, D.S.; Coyle, M. Niche partitioning and photosynthetic response of alectorioid lichens from subalpine spruce–fir forest in north-central British Columbia, Canada: The role of canopy microclimate gradients. Lichenologist 2003, 35, 157–175. [Google Scholar] [CrossRef]
- Gauslaa, Y.; Lie, M.; Ohlson, M. Epiphytic lichen biomass in a boreal Norway spruce forest. Lichenologist 2008, 40, 257–266. [Google Scholar] [CrossRef]
- Farber, L.; Solhaug, K.; Esseen, P.; Bilger, W.; Gauslaa, Y. Sunscreening fungal pigments influence the vertical gradient of pendulous lichens in boreal forest canopies. Ecology 2014, 95, 1464–1471. [Google Scholar] [CrossRef] [Green Version]
- Phinney, N.H.; Gauslaa, Y.; Solhaug, K.A. Why chartreuse? The pigment of vulpinic acid screens blue light in the lichen Letharia vulpine. Planta 2018. [Google Scholar] [CrossRef]
- Merinero, S.; Hilmo, O.; Gauslaa, Y. Size is a main driver for hydration traits in cyano- and cephalolichens of boreal rainforest canopies. Fungal Ecol. 2014, 7, 59–66. [Google Scholar] [CrossRef]
- Esseen, P.-A.; Rönnqvist, M.; Gauslaa, Y.; Coxson, D.S. Externally held water—A key factor for hair lichens in boreal forest canopies. Fungal Ecol. 2017, 30, 29–38. [Google Scholar] [CrossRef]
- Phinney, N.H.; Solhaug, K.A.; Gauslaa, Y. Rapid resurrection of chlorolichens in humid air: Specific thallus mass drives rehydration and reactivation kinetics. Environ. Exp. Bot. 2018, 148, 184–191. [Google Scholar] [CrossRef]
- Link, T.E.; Unsworth, M.H.; Marks, D. The dynamics of rainfall interception by a seasonal temperate rainforest. Agric. For. Meteorol. 2004, 124, 171–191. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, A.; Gauslaa, Y.; Palmqvist, K.; Ekström, M.; Esseen, P.-A. Morphology drives water storage traits in the globally widespread lichen genus Usnea. Fungal Ecol. 2018, 35, 51–61. [Google Scholar] [CrossRef]
- Esseen, P.-A.; Olsson, T.; Coxson, D.; Gauslaa, Y. Morphology influences water storage in hair lichens from boreal forest canopies. Fungal Ecol. 2015, 18, 26–35. [Google Scholar] [CrossRef]
- Lange, O.L.; Kilian, E.; Ziegler, H. Water vapor uptake and photosynthesis of lichens: Performance differences in species with green and blue-green algae as phycobionts. Oecologia 1986, 71, 104–110. [Google Scholar] [CrossRef]
- Stevenson, S.K.; Coxson, D.S. Arboreal forage lichens in partial cuts: A synthesis of research results from British Columbia, Canada. Rangifer 2007, 17, 155–165. [Google Scholar] [CrossRef]
- Ahlner, S. Utbredningstyper bland Nordiska barrträdslavar. Acta Phytogeogr. Suec. 1948, 22, 1–257. [Google Scholar]
- Bruteig, I.E. Large-scale survey of the distribution and ecology of common epiphytic lichens on Pinus sylvestris in Norway. Ann. Bot. Fenn. 1993, 30, 161–179. [Google Scholar]
- Goward, T. Observations on the ecology of the lichen genus Bryoria in high elevation conifer forests. Can. Field Nat. 1998, 112, 496–501. [Google Scholar]
- Bjerke, J.W.; Elvebakk, A.; Elverland, E. The lichen genus Usnea in Norway north of the Arctic Circle: Biogeography and ecology. Nova Hedwig. 2006, 83, 293–309. [Google Scholar] [CrossRef]
- Van Herk, C.M. Mapping of ammonia pollution with epiphytic lichens in The Netherlands. Lichenologist 1999, 31, 9–20. [Google Scholar]
- Frati, L.; Santoni, S.; Nicolardi, V.; Gaggi, C.; Brunialti, G.; Guttova, A.; Gaudino, S.; Pati, A.; Pirintsos, S.A.; Loppi, S. Lichen biomonitoring of ammonia emission and nitrogen deposition around a pig stockfarm. Environ. Pollut. 2007, 146, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Pinho, P.; Theobald, M.R.; Dias, T.; Tang, Y.S.; Cruz, C.; Martins- Loução, M.A.; Sutton, M.; Branquinho, C. Critical loads of nitrogen deposition and critical levels of atmospheric ammonia for semi-natural Mediterranean evergreen woodlands. Biogeosciences 2012, 9, 1205–1215. [Google Scholar] [CrossRef] [Green Version]
- Giordani, P.; Calatayud, V.; Stofer, S.; Seidling, W.; Granke, O.; Fischer, R. Detecting the nitrogen critical loads on European forests by means of epiphytic lichens. A signal-to-noise evaluation. For. Ecol. Manag. 2014, 311, 29–40. [Google Scholar] [CrossRef]
- Cristofolini, F.; Giordani, P.; Gottardini, E.; Modenesi, P. The response of epiphytic lichens to air pollution and subsets of ecological predictors: A case study from the Italian Prealps. Environ. Pollut. 2008, 151, 308–317. [Google Scholar] [CrossRef]
- Giordani, P.; Malaspina, P. Do tree-related factors mediate the response of lichen functional groups to eutrophication? Plant Byosyst. 2016. [Google Scholar] [CrossRef]
- Nascimbene, J.; Fontana, V.; Spitale, D. A multi-taxon approach reveals the effect of management intensity on biodiversity in Alpine larch grasslands. Sci. Total Environ. 2014, 487, 110–116. [Google Scholar] [CrossRef]
- Root, H.T.; Geiser, L.H.; Fenn, M.E.; Jovan, S.; Hutten, M.A.; Ahuja, S.; Dillman, K.; Schirokauer, D.; Berryman, S.; McMurray, J.A. A simple tool for estimating throughfall nitrogen deposition in forests of western North America using lichens. For. Ecol. Manag. 2013, 306, 1–8. [Google Scholar] [CrossRef]
- Giordani, P.; Matteucci, E.; Redana, M.; Ferrarese, A.; Isocrono, D. Unsustainable cattle load in alpine pastures alters the diversity and the composition of lichen functional groups for nitrogen requirement. Fungal Ecol. 2014, 9, 69–72. [Google Scholar] [CrossRef]
- Van Herk, C.M.; Mathijssen-Spiekman, E.A.M.; de Zwart, D. Long distance nitrogen air pollution effects on lichens in Europe. Lichenologist 2003, 35, 347–359. [Google Scholar] [CrossRef]
- Geiser, L.H.; Jovan, S.E.; Glavich, D.A.; Porter, M.K. Lichen-based critical loads for atmospheric nitrogen deposition in western Oregon and Washington forests, USA. Environ. Pollut. 2010, 158, 2412–2421. [Google Scholar] [CrossRef]
- Bermejo-Orduna, R.; McBride, J.R.; Shiraishi, K.; Elustondo, D.; Lasheras, E.; Santamaría, J.M. Biomonitoring of traffic-related nitrogen pollution using Letharia vulpina (L.) Hue in the Sierra Nevada, California. Sci. Total Environ. 2014, 490, 205–212. [Google Scholar] [CrossRef]
- Johansson, O.; Olofsson, J.; Giesler, R.; Palmqvist, K. Lichen responses to nitrogen and phosphorus additions can be explained by the different symbiont responses. New Phytol. 2011, 191, 795–805. [Google Scholar] [CrossRef] [Green Version]
- Johansson, O.; Palmqvist, K.; Olofsson, J. Nitrogen deposition drives lichen community changes through differential species responses. Glob. Chang. Biol. 2012, 18, 26–35. [Google Scholar] [CrossRef]
- Hauck, M.; de Bruyn, U.; Leuschner, C. Dramatic diversity losses in epiphytic lichens in temperate broad-leaved forests during the last 150 years. Biol. Conserv. 2013, 157, 136–145. [Google Scholar] [CrossRef]
- McCune, B.; Geiser, L. Macrolichens of the Pacific Northwest, 2nd ed.; Oregon State University Press: Corvallis, OR, USA, 2009. [Google Scholar]
- McMurray, J.A.; Roberts, D.W.; Geiser, L.H. Epiphytic lichen indication of nitrogen deposition and climate in the northern rocky mountains, USA. Ecol. Indic. 2015, 49, 154–161. [Google Scholar] [CrossRef]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nascimbene, J.; Benesperi, R.; Giordani, P.; Grube, M.; Marini, L.; Vallese, C.; Mayrhofer, H. Could Hair-Lichens of High-Elevation Forests Help Detect the Impact of Global Change in the Alps? Diversity 2019, 11, 45. https://doi.org/10.3390/d11030045
Nascimbene J, Benesperi R, Giordani P, Grube M, Marini L, Vallese C, Mayrhofer H. Could Hair-Lichens of High-Elevation Forests Help Detect the Impact of Global Change in the Alps? Diversity. 2019; 11(3):45. https://doi.org/10.3390/d11030045
Chicago/Turabian StyleNascimbene, Juri, Renato Benesperi, Paolo Giordani, Martin Grube, Lorenzo Marini, Chiara Vallese, and Helmut Mayrhofer. 2019. "Could Hair-Lichens of High-Elevation Forests Help Detect the Impact of Global Change in the Alps?" Diversity 11, no. 3: 45. https://doi.org/10.3390/d11030045
APA StyleNascimbene, J., Benesperi, R., Giordani, P., Grube, M., Marini, L., Vallese, C., & Mayrhofer, H. (2019). Could Hair-Lichens of High-Elevation Forests Help Detect the Impact of Global Change in the Alps? Diversity, 11(3), 45. https://doi.org/10.3390/d11030045