The Role of Maximum Shelf Depth versus Distance from Shore in Explaining a Diversity Gradient of Mushroom Corals (Fungiidae) off Jakarta



Abstract
:1. Introduction
2. Materials and Methods
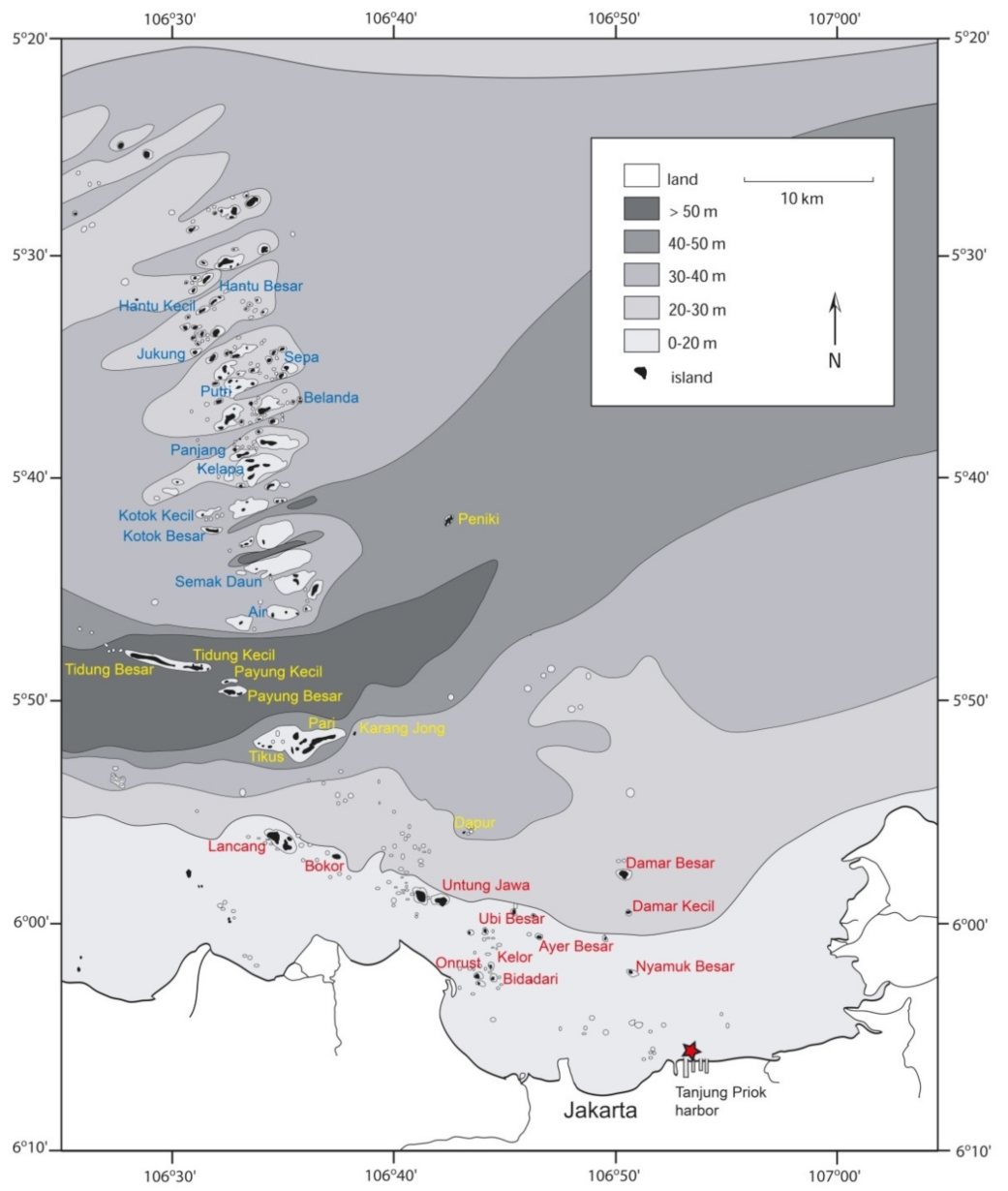
2.1. Research Area and Abiotic Parameters
2.2. Coral Data
2.3. Data Analysis
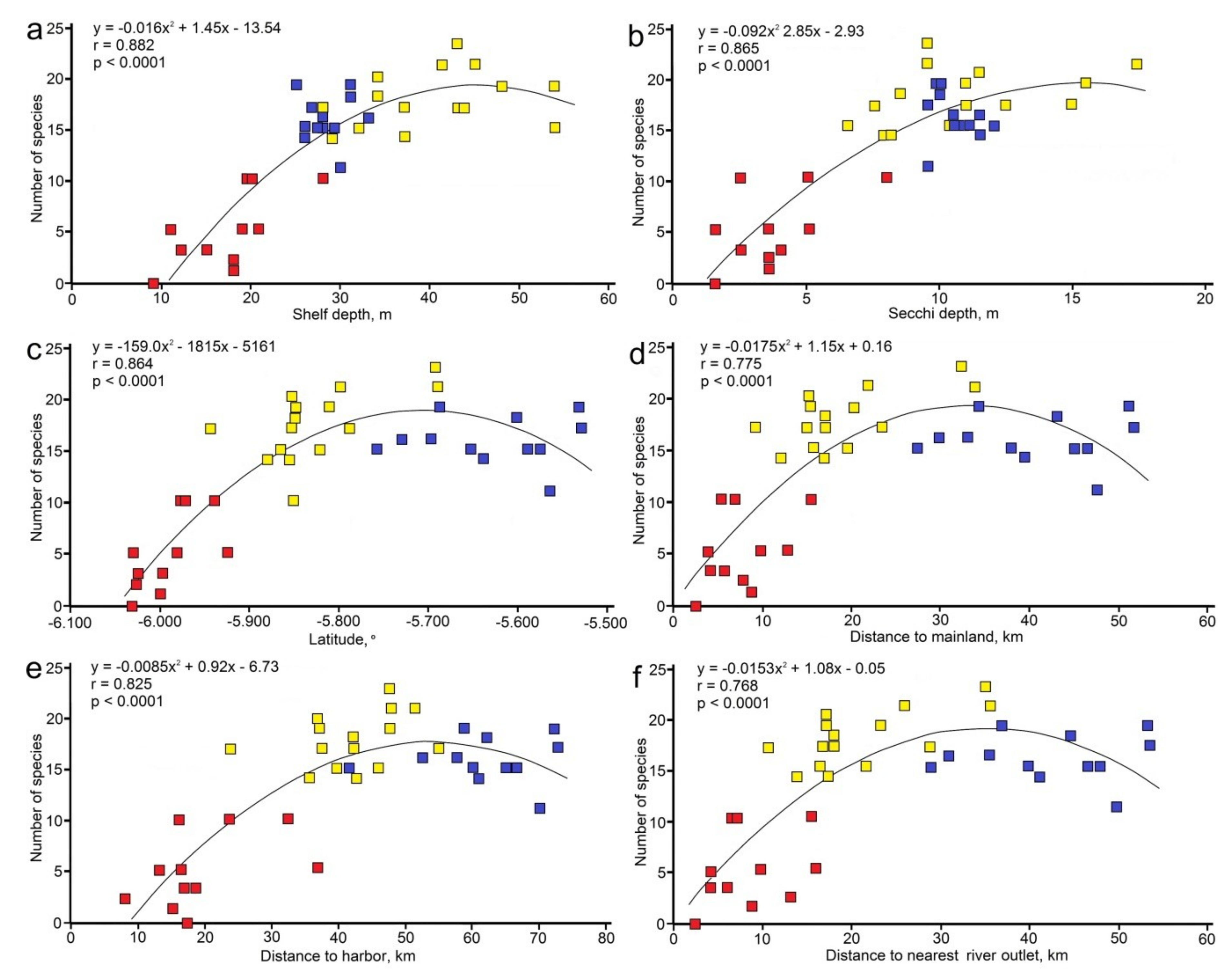
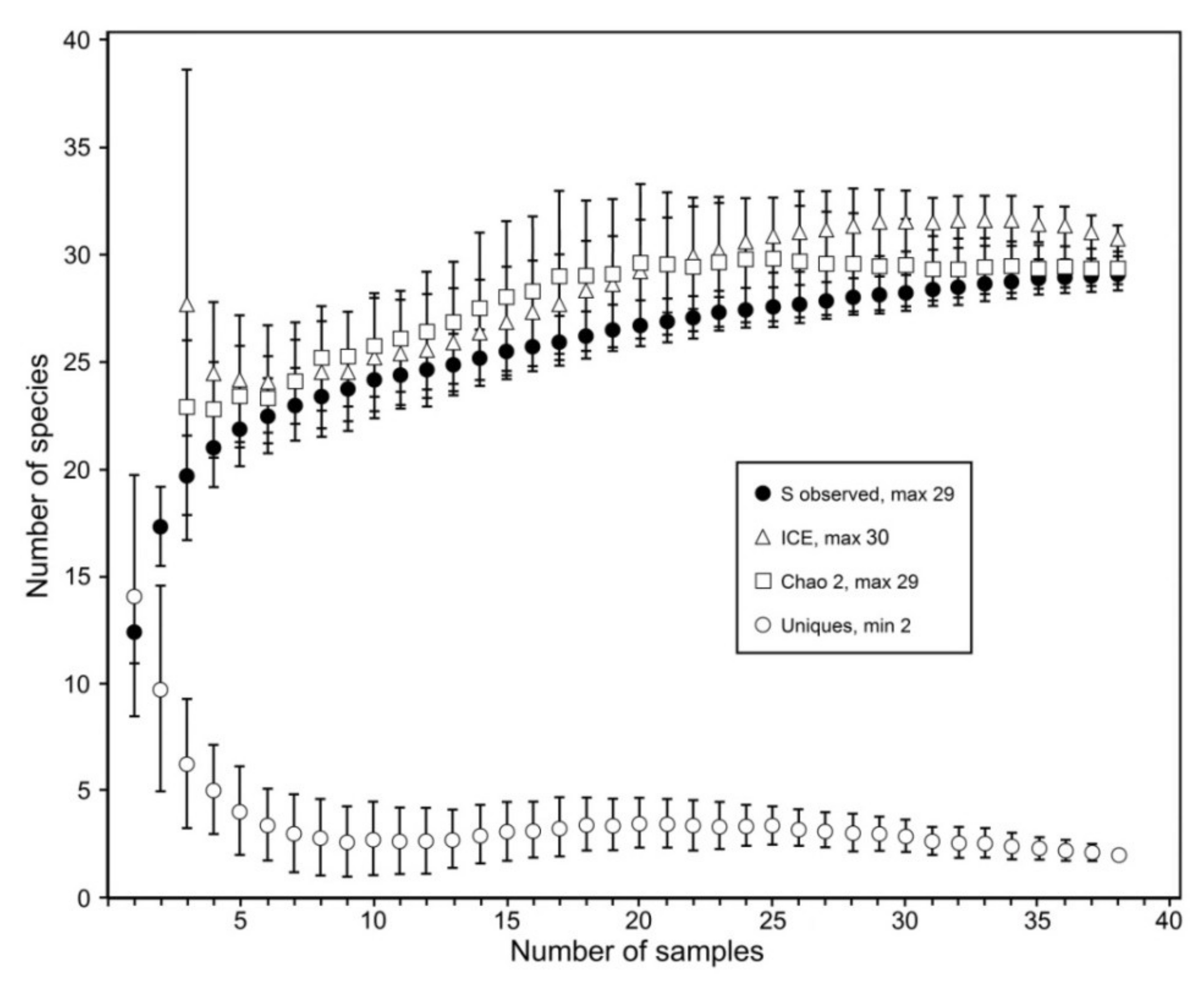
3. Results
3.1. Abiotic Parameters
3.2. Species Diversity Patterns
3.2.1. Entire Depth Range
3.2.2. Shallow Depths
3.2.3. Species Richness Estimators
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Heery, E.C.; Hoeksema, B.W.; Browne, N.K.; Reimer, J.D.; Ang, P.O.; Huang, D.; Friess, D.A.; Chou, L.M.; Loke, L.H.L.; Saksena-Taylor, P.; et al. Urban coral reefs: Degradation and resilience of hard coral assemblages in coastal cities of East and Southeast Asia. Mar. Pollut. Bull. 2018, 135, 654–681. [Google Scholar] [CrossRef]
- Cleary, D.F.R.; Becking, L.E.; de Voogd, N.J.; Renema, W.; de Beer, M.; van Soest, R.W.M.; Hoeksema, B.W. Variation in the diversity and composition of benthic taxa as a function of distance offshore, depth and exposure in the Spermonde Archipelago, Indonesia. Estuar. Coast. Shelf Sci. 2005, 65, 557–570. [Google Scholar] [CrossRef]
- Cleary, D.F.R.; DeVantier, L.; Vail, L.; Manto, P.; de Voogd, N.J.; Rachello-Dolmen, P.G.; Tuti, Y.; Budiyanto, A.; Wolstenholme, J.; Hoeksema, B.W.; et al. Relating variation in species composition to environmental variables: A multi-taxon study in an Indonesian coral reef complex. Aquat. Sci. 2008, 70, 419–431. [Google Scholar] [CrossRef]
- Huang, D.; Tun, K.P.P.; Chou, L.M.; Todd, P.A. An inventory of zooxanthellate scleractinian corals in Singapore, including 33 new records. Raffles Bull. Zool. 2009, 22, 69–80. [Google Scholar]
- Sawall, Y.; Teichberg, M.C.; Seemann, J.; Litaay, M.; Richter, C. Nutritional status and metabolism of the coral Stylophora subseriata along a eutrophication gradient in Spermonde Archipelago (Indonesia). Coral Reefs 2011, 30, 841–853. [Google Scholar] [CrossRef]
- Polónia, A.R.; Cleary, D.F.R.; de Voogd, N.J.; Renema, W.; Hoeksema, B.W.; Martins, A.; Gomes, N.C.M. Habitat and water quality variables as predictors of community composition in an Indonesian coral reef: A multi-taxon study in the Spermonde Archipelago. Sci. Total Environ. 2015, 537, 139–151. [Google Scholar] [CrossRef]
- Plass-Johnson, J.G.; Heiden, J.P.; Abu, N.; Lukman, M.; Teichberg, M. Experimental analysis of the effects of consumer exclusion on recruitment and succession of a coral reef system along a water quality gradient in the Spermonde Archipelago, Indonesia. Coral Reefs 2016, 35, 229–243. [Google Scholar] [CrossRef]
- Hopley, D. Continental shelf reef systems. In Coastal evolution, Late Quaternary Shoreline Morphodynamics; Carter, R.W.G., Woodroffe, C.D., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 303–340. [Google Scholar]
- Janßen, A.; Wizemann, A.; Klicpera, A.; Satari, D.Y.; Westphal, H.; Mann, T. Sediment composition and facies of coral reef islands in the Spermonde Archipelago, Indonesia. Front. Mar. Sci. 2017, 4, 144. [Google Scholar] [CrossRef]
- Fabricius, K.E. Effects of terrestrial runoff on the ecology of corals and coral reefs: Review and synthesis. Mar. Pollut. Bull. 2005, 50, 125–146. [Google Scholar] [CrossRef]
- Done, T.J. Patterns in the distribution of coral communities across the Central Great Barrier Reef. Coral Reefs 1982, 1, 95–107. [Google Scholar] [CrossRef]
- Williams, D.M.B. Patterns in the distribution of fish communities across the Great Barrier Reef. Coral Reefs 1982, 1, 35–43. [Google Scholar] [CrossRef]
- Dinesen, Z.D. Patterns in the distribution of soft corals across the Central Great Barrier Reef. Coral Reefs 1983, 1, 229–236. [Google Scholar] [CrossRef]
- Preston, N.P.; Doherty, P.J. Cross-shelf patterns in the community structure of coral-dwelling Crustacea in the central region of the Great Barrier Reef. II. Cryptofauna. Mar. Ecol. Prog. Ser. 1994, 104, 27–38. [Google Scholar] [CrossRef]
- Page, C.; Willis, B. Distribution, host range and large-scale spatial variability in black band disease prevalence on the Great Barrier Reef, Australia. Dis. Aquat. Organ. 2006, 69, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becking, L.E.; Cleary, D.F.R.; de Voogd, N.J.; Renema, W.; de Beer, M.; van Soest, R.W.M.; Hoeksema, B.W. Beta-diversity of tropical marine assemblages in the Spermonde Archipelago, Indonesia. Mar. Ecol. 2006, 27, 76–88. [Google Scholar] [CrossRef]
- Hoeksema, B.W. Distribution patterns of mushroom corals (Scleractinia:Fungiidae) across the Spermonde Shelf, Indonesia. Raffles Bull. Zool. 2012, 60, 183–212. [Google Scholar]
- Haya, L.O.M.; Fujii, M. Mapping the change of coral reefs using remote sensing and in situ measurements: A case study in Pangkajene and Kepulauan Regency, Spermonde Archipelago, Indonesia. J. Oceanogr. 2017, 73, 623–645. [Google Scholar] [CrossRef]
- Teichberg, M.; Wild, C.; Bednarz, V.N.; Kegler, H.F.; Lukman, M.; Gärdes, A.A.; Heiden, J.P.; Weiand, L.; Abu, N.; Nasir, A.; et al. Spatio-temporal patterns in coral reef communities of the Spermonde Archipelago, 2012–2014, I: Comprehensive reef monitoring of water and benthic indicators reflect changes in reef health. Front. Mar. Sci. 2018, 5, 33. [Google Scholar] [CrossRef]
- Kunzmann, A. The coral reefs of West Sumatra. In The Ecology of the Indonesian Seas; Tomasick, T., Mah, A.J., Nontji, A., Moosa, M.K., Eds.; Periplus: Singapore, 1997; Part II; pp. 1249–1262. [Google Scholar]
- Johan, O.; Prasetio, A.B.; Ardi, I.; Syam, A.R.; Quinn, N.J. The abundance of ornamental corals after mass die off in 1997 on the Padang Shelf Reef System, West Sumatera, Indonesia. J. Trop. Mar. Sci. Technol. 2016, 8, 29–38. [Google Scholar] [CrossRef]
- Renema, W. Habitat variables determining the occurrence of large benthic foraminifera in the Berau area (East Kalimantan, Indonesia). Coral Reefs 2006, 25, 351–359. [Google Scholar] [CrossRef]
- Tarya, A.; Hoitink, A.J.F.; van der Vegt, M. Tidal and subtidal flow patterns on a tropical continental shelf semi-insulated by coral reefs. J. Geophys. Res. 2010, 115, C09029. [Google Scholar] [CrossRef]
- Tarya, A.; Hoitink, A.J.F.; van der Vegt, M.; van Katwijk, M.M.; Hoeksema, B.W.; Bouma, T.J.; Lamers, L.P.M.; Christianen, M.J.A. Exposure of coastal ecosystems to river plume spreading across a near-equatorial continental shelf. Cont. Shelf Res. 2018, 153, 1–15. [Google Scholar] [CrossRef]
- Waheed, Z.; Hoeksema, B.W. A tale of two winds: Species richness patterns of reef corals around the Semporna peninsula, Malaysia. Mar. Biodivers. 2013, 43, 37–51. [Google Scholar] [CrossRef]
- DeVantier, L.; Turak, E. Coral Reefs of Brunei Darussalam; Fisheries Department, Ministry of Industry and Primary Resources, Government of Brunei Darussalam: Bandar Seri Begawan, Brunei Darussalam, 2009; pp. 1–99.
- Hoeksema, B.W.; Lane, D.J.W. The mushroom coral fauna (Scleractinia Fungiidae) of Brunei Darussalam (South China Sea) and its relation to the Coral Triangle. Raffles Bull. Zool. 2014, 62, 566–580. [Google Scholar]
- Brown, C.; Browne, N.; McIlwain, J.L.; Zinke, Z. Inshore, turbid coral reefs from northwest Borneo exhibiting low diversity, but high cover show evidence of resilience to various environmental stressors. PeerJ Preprints 2018. [Google Scholar] [CrossRef]
- Wild, C.; Jantzen, C.; Kremb, S.G. Turf algae-mediated coral damage in coastal reefs of Belize, Central America. PeerJ 2014, 2, e571. [Google Scholar] [CrossRef] [PubMed]
- Baumann, J.H.; Townsend, J.E.; Courtney, T.A.; Aichelman, H.E.; Davies, S.W.; Lima, F.P.; Castillo, K.D. Temperature regimes impact coral assemblages along environmental gradients on lagoonal reefs in Belize. PLoS ONE 2016, 11, e0162098. [Google Scholar] [CrossRef] [PubMed]
- Hoeksema, B.W. Mushroom corals (Scleractinia: Fungiidae) of Madang Lagoon, northern Papua New Guinea: An annotated checklist with the description of Cantharellus jebbi spec. nov. Zool. Meded. 1993, 67, 1–19. [Google Scholar]
- Langer, M.R.; Lipps, J.H. Foraminiferal distribution and diversity, Madang Reef and Lagoon, Papua New Guinea. Coral Reefs 2003, 22, 143–154. [Google Scholar] [CrossRef]
- Adjeroud, M.; Fernandez, J.M.; Carroll, A.G.; Harrison, P.L.; Penin, L. Spatial patterns and recruitment processes of coral assemblages among contrasting environmental conditions in the southwestern lagoon of New Caledonia. Mar. Pollut. Bull. 2010, 61, 375–386. [Google Scholar] [CrossRef]
- Gilbert, A.; Heintz, T.; Hoeksema, B.W.; Benzoni, F.; Fernandez, J.M.; Fauvelot, C.; Andréfouët, S. Endangered New Caledonian endemic mushroom coral Cantharellus noumeae in turbid, metal-rich, natural and artificial environments. Mar. Pollut. Bull. 2015, 10, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Furby, K.A.; Bouwmeester, J.; Berumen, M.L. Susceptibility of central Red Sea corals during a major bleaching event. Coral Reefs 2013, 32, 505–513. [Google Scholar] [CrossRef]
- Ellis, J.; Anlauf, H.; Kürten, S.; Lozano-Cortés, D.; Alsaffar, Z.; Cúrdia, J.; Jones, B.; Carvalho, S. Cross shelf benthic biodiversity patterns in the southern Red Sea. Sci. Rep. 2017, 7, 437. [Google Scholar] [CrossRef] [PubMed]
- Monroe, A.A.; Ziegler, M.; Roik, A.; Röthig, T.; Hardenstine, R.S.; Emms, M.A.; Jensen, T.; Voolstra, C.R.; Berumen, M.L. In situ observations of coral bleaching in the central Saudi Arabian Red Sea during the 2015/2016 global coral bleaching event. PLoS ONE 2018, 13, e0195814. [Google Scholar] [CrossRef] [PubMed]
- Umbgrove, J.H.F. De koraalriffen der Duizend Eilanden (Java-Zee). Wetensch. Meded. Dienst Mijnb. Nederl. Ind. 1929, 12, 1–47. [Google Scholar]
- Umbgrove, J.H.F. Coral reefs of the East Indies. Bull. Geol. Soc. Am. 1947, 58, 729–778. [Google Scholar] [CrossRef]
- Suyarso. Lingkungan fisik pantai dan dasar perairan Teluk Jakarta. In Atlas Oseanologi Teluk Jakarta; Suyarso, Ed.; Pusat Penelitian dan Pengembangan Oseanalogi, Lembaga Ilmu Pengetahuan Indonesia: Jakarta, Indonesia, 1995; pp. 21–27. [Google Scholar]
- Jordan, C.F. The Sedimentology of Kepulauan Seribu: A Modern Patch Reef Complex in the Java Sea, Indonesia; Indonesian Petroleum Association: Jakarta, Indonesia, 1998; pp. 1–81. [Google Scholar]
- Hoeksema, B.W. Delineation of the Indo-Malayan centre of maximum marine biodiversity: The Coral Triangle. In Biogeography, Time, and Place: Distributions, Barriers, and Islands; Renema, W., Ed.; Springer: Dordrecht, The Netherlands, 2007; pp. 117–178. [Google Scholar]
- Umbgrove, J.H.F. De koraalriffen in de Baai van Batavia. Wetensch. Meded. Dienst Mijnb. Nederl. Ind. 1928, 7, 1–68. [Google Scholar]
- Umbgrove, J.H.F.; Verwey, J. The coral reefs in the Bay of Batavia. I. Geology. In Proceedings of the 4th Pacific Science Congress, Bandung, Indonesia, 16–25 May 1929; Excursion A2. pp. 1–30. [Google Scholar]
- Yates, B.F. Implementing coastal zone management policy: Kepulauan Seribu Marine National Park, Indonesia. Coast. Manag. 1994, 22, 235–249. [Google Scholar] [CrossRef]
- UNESCO. Reducing Megacity Impacts on the Coastal Environment—Alternative Livelihoods and Waste Management in Jakarta and the Seribu Islands; Coastal Region and Small Islands Paper 6; UNESCO: Paris, France, 2000; pp. 1–59. [Google Scholar]
- Fauzi, A.; Buchary, E.A. A Socioeconomic perspective of environmental degradation at Kepulauan Seribu Marine National Park, Indonesia. Coast. Manag. 2002, 30, 167–181. [Google Scholar] [CrossRef]
- Farhan, A.R.; Lim, S. Integrated vulnerability assessment on small island regions towards integrated coastal zone management (ICZM): A case study of Thousand Islands, Indonesia. Int. J. Geoinform. 2014, 10, 1–16. [Google Scholar]
- Baum, G.; Kusumanti, I.; Breckwoldt, A.; Ferse, S.C.A.; Glaser, M.; Dwiyitno; Adrianto, L.; van der Wulp, S.; Kunzmann, A. Under pressure: Investigating marine resource-based livelihoods in Jakarta Bay and the Thousand Islands. Mar. Pollut. Bull. 2016, 110, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Zaneveld, J.S.; Verstappen, H.T. A recent investigation about the geomorphology and the flora of some coral islands in the Bay of Djakarta. J. Sci. Res. 1952, 2–3, 58–68. [Google Scholar]
- Verstappen, H.T. Djakarta Bay. A Geomorphological Study on Shoreline Development. Ph.D. Thesis, Utrecht University, Utrecht, The Netherlands, 1953; pp. 1–101. [Google Scholar]
- Verstappen, H.T. Old and new observations on coastal changes of Jakarta Bay: An example of trends in urban stress on coastal environments. J. Coast. Res. 1988, 4, 573–587. [Google Scholar]
- Stoddart, D.R. Umbgrove’s islands revisited. UNESCO Rep. Mar. Sci. 1986, 40, 80–98. [Google Scholar]
- Han, Q.; Schaefer, W.; Barry, N. Land reclamation using waste as fill material: A case study in Jakarta. Int. J. Environ. Ecol. Eng. 2013, 7, 318–327. [Google Scholar]
- Van der Wulp, S.A.; Dsikowitzky, L.; Hesse, K.J.; Schwarzbauer, J. Master Plan Jakarta, Indonesia: The Giant Seawall and the need for structural treatment of municipal waste water. Mar. Pollut. Bull. 2016, 110, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Erftemeijer, P.L.A.; Riegl, B.; Hoeksema, B.W.; Todd, P.A. Environmental impacts of dredging and other sediment disturbances on corals: A review. Mar. Pollut. Bull. 2012, 64, 1737–1765. [Google Scholar] [CrossRef] [PubMed]
- Cleary, D.F.R.; Polónia, A.R.M.; Renema, W.; Hoeksema, B.W.; Wolstenholme, J.; Tuti, Y.; de Voogd, N.J. Coral reefs next to a major conurbation: A study of temporal change (1985–2011) in coral cover and composition in the reefs of Jakarta, Indonesia. Mar. Ecol. Prog. Ser. 2014, 501, 89–98. [Google Scholar] [CrossRef]
- Baum, G.; Januar, H.I.; Ferse, S.C.A.; Kunzmann, A. Local and regional impacts of pollution on coral reefs along the thousand islands north of the megacity Jakarta, Indonesia. PLoS ONE 2015, 10, e0138271. [Google Scholar] [CrossRef] [PubMed]
- Breckwoldt, A.; Dsikowitzky, L.; Baum, G.; Ferse, S.C.A.; van der Wulp, S.; Kusumanti, I.; Ramadhan, A.; Adrianto, L. A review of stressors, uses and management perspectives for the larger Jakarta Bay Area, Indonesia. Mar. Pollut. Bull. 2016, 110, 790–794. [Google Scholar] [CrossRef]
- Dsikowitzky, L.; Ferse, S.; Schwarzbauer, J.; Vogt, T.S.; Irianto, H.E. Impacts of megacities on tropical coastal ecosystems—The case of Jakarta, Indonesia. Mar. Pollut. Bull. 2016, 110, 621–623. [Google Scholar] [CrossRef] [PubMed]
- Van der Wulp, S.A.; Hesse, K.J.; Ladwig, N.; Damar, A. Numerical simulations of river discharges, nutrient flux and nutrient dispersal in Jakarta Bay, Indonesia. Mar. Pollut. Bull. 2016, 110, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Dsikowitzky, L.; Schäfer, L.; Ariyani, F.; Irianto, H.E.; Schwarzbauer, J. Evidence of massive river pollution in the tropical megacity Jakarta as indicated by faecal steroid occurrence and the seasonal flushing out into the coastal ecosystem. Environ. Chem. Lett. 2017, 15, 703–708. [Google Scholar] [CrossRef]
- Van der Meij, S.E.T.; Moolenbeek, R.G.; Hoeksema, B.W. Decline of the Jakarta Bay molluscan fauna linked to human impact. Mar. Pollut. Bull. 2009, 59, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Van der Meij, S.E.T.; Suharsono; Hoeksema, B.W. Long-term changes in coral assemblages under natural and anthropogenic stress in Jakarta Bay (1920–2005). Mar. Pollut. Bull. 2010, 60, 1442–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, B.E. Damage and recovery of coral reefs affected by El Niño related seawater warming in the Thousand Islands, Indonesia. Coral Reefs 1990, 8, 163–170. [Google Scholar] [CrossRef]
- Hoeksema, B.W. Control of bleaching in mushroom coral populations (Scleractinia, Fungiidae) in the Java Sea: Stress tolerance and interference by life history strategy. Mar. Ecol. Prog. Ser. 1991, 74, 225–237. [Google Scholar] [CrossRef]
- Aziz, A.; Sukarno. Preliminary observation on living habits of Acanthaster planci (Linnaeus) at Palau Tikus, Seribu Islands. Mar. Res. Indon. 1977, 17, 121–132. [Google Scholar]
- Aziz, A. Beberapa catatan tentang kehadiran bintang laut jenis Acanthaster planci di perairan Indonesia. Oseana 1995, 20, 23–31. [Google Scholar]
- Baird, A.H.; Pratchett, M.S.; Hoey, A.S.; Herdiana, Y.; Campbell, S.J. Acanthaster planci is a major cause of coral mortality in Indonesia. Coral Reefs 2013, 32, 803–812. [Google Scholar] [CrossRef]
- Sheppard, C.R.C. Coral cover, zonation and diversity on reef slopes of Chagos Atolls, and population structures of the major species. Mar. Ecol. Prog. Ser. 1980, 2, 193–205. [Google Scholar] [CrossRef]
- Sheppard, C.R.C. The reef and soft-substrate coral fauna of Chagos, Indian Ocean. J. Nat. Hist. 1981, 15, 607–621. [Google Scholar] [CrossRef]
- Graus, R.R.; Macintyre, I.G. The zonation patterns of Caribbean coral reefs as controlled by wave and light energy input, bathymetric setting and reef morphology: Computer simulation experiments. Coral Reefs 1989, 8, 9–18. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Moka, W. Species assemblages and phenotypes of mushroom corals (Scleractinia: Fungiidae) related to coral reef habitats in the Flores Sea. Neth. J. Sea Res. 1989, 23, 149–160. [Google Scholar] [CrossRef]
- Hoeksema, B.W. Evolutionary trends in onshore-offshore distribution patterns of mushroom coral species (Scleractinia: Fungiidae). Contrib. Zool. 2012, 81, 199–221. [Google Scholar]
- Lane, D.J.W.; Hoeksema, B.W. Mesophotic mushroom coral records at Brunei Darussalam support westward extension of the Coral Triangle to the South China Sea waters of Northwest Borneo. Raffles Bull. Zool. 2016, 64, 566–580. [Google Scholar]
- Hoeksema, B.W. Latitudinal species diversity gradient of mushroom corals off eastern Australia: A baseline from the 1970s. Estuar. Coast. Shelf Sci. 2015, 165, 190–198. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Sellanes, J.; Easton, E.E. A high-latitude, mesophotic Cycloseris field at 85 m depth off Rapa Nui (Easter Island). Bull. Mar. Sci. 2019, 95, 101–102. [Google Scholar] [CrossRef]
- Goreau, T.F.; Yonge, C.M. Coral community on muddy sand. Nature 1968, 217, 421–423. [Google Scholar] [CrossRef]
- Pichon, M. Free living scleractinian coral communities in the coral reefs of Madagascar. In Proceedings of the 2nd International Coral Reef Symposium, Brisbane, Australian, 2 June–22 July 1974; Volume 2, pp. 173–181. [Google Scholar]
- Fisk, D.A. Free-living corals: Distributions according to plant cover, sediments, hydrodynamics, depth and biological factors. Mar. Biol. 1983, 74, 287–294. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Cairns, S. World List of Scleractinia. Accessed through: World Register of Marine Species. Available online: http://www.marinespecies.org/scleractinia/ (accessed on 31 January 2019).
- Hoeksema, B.W. Attached mushroom corals (Scleractinia: Fungiidae) in sediment-stressed reef conditions at Singapore, including a new species and a new record. Raffles Bull. Zool. 2009, Suppl. 22, 81–90. [Google Scholar]
- Benzoni, F.; Arrigoni, R.; Stefani, F.; Reijnen, B.T.; Montano, S.; Hoeksema, B.W. Phylogenetic position and taxonomy of Cycloseris explanulata and C. wellsi (Scleractinia: Fungiidae): Lost mushroom corals find their way home. Contrib. Zool. 2012, 81, 125–146. [Google Scholar]
- Hoeksema, B.W. Evolution of body size in mushroom corals (Scleractinia: Fungiidae) and its ecomorphological consequences. Neth. J. Zool. 1991, 41, 122–139. [Google Scholar] [CrossRef]
- Gittenberger, A.; Reijnen, B.T.; Hoeksema, B.W. A molecularly based phylogeny reconstruction of mushroom corals (Scleractinia: Fungiidae) with taxonomic consequences and evolutionary implications for life history traits. Contrib. Zool. 2011, 80, 107–132. [Google Scholar]
- Hoeksema, B.W. The “Fungia patella group” (Scleractinia, Fungiidae) revisited with a description of the mini mushroom coral Cycloseris boschmai sp. n. ZooKeys 2014, 371, 57–84. [Google Scholar] [CrossRef]
- Umbgrove, J.H.F. Madreporaria from the Bay of Batavia. Zool. Meded. 1939, 22, 1–64. [Google Scholar]
- Moll, H.; Suharsono. Distribution, diversity and abundance of reef corals in Jakarta Bay and Kepulauan Seribu. UNESCO Rep. Mar. Sci. 1986, 40, 112–125. [Google Scholar]
- Soemodihardjo, S. (Ed.) Proceedings: Coral Reef Evaluation Workshop Pulau Seribu, Jakarta, Indonesia, 11–20 September 1995; Contending with Global Change 10; UNESCO: Jakarta, Indonesia, 1998; pp. 1–129. [Google Scholar]
- Hoeksema, B.W. Taxonomy, phylogeny and biogeography of mushroom corals (Scleractinia: Fungiidae). Zool. Verh. 1989, 254, 1–295. [Google Scholar]
- Moll, H. Zonation and Diversity of Scleractinia on Reefs off S.W. Sulawesi, Indonesia. Ph.D. Thesis, Leiden University, Leiden, The Netherlands, 1983. [Google Scholar]
- Schuhmacher, H.; Mergner, H. Quantitative Analyse von Korallengemeinschaften des Sanganeb-Atolls (mittleres Rotes Meer). II. Vergleich mit einem Riffareal bei Aqaba (nördliches Rotes Meer) am Nordrande des indopazifischen Riffgürtels. Helgol. Meeresunters. 1985, 39, 419–440. [Google Scholar] [CrossRef] [Green Version]
- Waheed, Z.; Hoeksema, B.W. Diversity patterns of scleractinian corals at Kota Kinabalu, Malaysia, in relation to exposure and depth. Raffles Bull. Zool. 2014, 62, 66–82. [Google Scholar]
- Waheed, Z.; van Mil, H.G.J.; Syed Hussein, M.A.; Jumin, R.; Golam Ahad, B.; Hoeksema, B.W. Coral reefs at the northernmost tip of Borneo: An assessment of scleractinian species richness patterns and benthic reef assemblages. PLoS ONE 2015, 10, e0146006. [Google Scholar] [CrossRef]
- Muir, P.R.; Wallace, C.C.; Bridge, T.C.L.; Bongaerts, P. Diverse staghorn coral fauna on the mesophotic reefs of North-East Australia. PLoS ONE 2015, 10, e0117933. [Google Scholar] [CrossRef] [PubMed]
- Muir, P.R.; Wallace, C.C.; Pichon, M.; Bongaerts, P. High species richness and lineage diversity of reef corals in the mesophotic zone. Proc. R. Soc. B 2018, 285, 20181987. [Google Scholar] [CrossRef]
- Muir, P.R.; Pichon, M.; Squire, L.; Wallace, C.C. Acropora tenella, a zooxanthellate coral extending to 110-m depth in the northern Coral Sea. Mar. Biodivers. 2018. [Google Scholar] [CrossRef]
- Larcombe, P.; Woolfe, K.J. Increased sediment supply to the Great Barrier Reef will not increase sediment accumulation at most coral reefs. Coral Reefs 1999, 18, 163–169. [Google Scholar] [CrossRef]
- Larcombe, P.; Costen, A.; Woolfe, K.J. The hydrodynamic and sedimentary setting of nearshore coral reefs, central Great Barrier Reef shelf, Australia: Paluma Shoals, a case study. Sedimentology 2001, 48, 811–835. [Google Scholar] [CrossRef]
- Tuti, M.I.Y.; Soemodihardjo, S. (Eds.) Coral Reef Ecosystem in the Seribu Islands; LIPI Press: Jakarta, Indonesia, 2006; pp. 1–179. [Google Scholar]
- Brown, B.E. (Ed.) Human Induced Damage to Coral Reefs. UNESCO Rep. Mar. Sci. 1986, 40, 1–180. [Google Scholar]
- Cleary, D.F.R.; Suharsono; Hoeksema, B.W. Coral diversity across a disturbance gradient in the Pulau Seribu reef complex off Jakarta, Indonesia. Biodivers. Conserv. 2006, 15, 3653–3674. [Google Scholar] [CrossRef]
- TNI-AL. Pulau-Pulau Seribu. Pulau Rambut Hingga Karang Jalan, Sekala 1:20.000, 2nd ed.; Tentara Nasional Indonesia—Ankatan Laut, Jawatan Hidro-Oseanografi: Jakarta, Indonesia, 1997; 410-KK. [Google Scholar]
- TNI-AL. Teluk Jakarta. Pulau Rambut Hingga Pelabuhan Tanjungpriok, Sekala 1:50.000, 2nd ed.; Tentara Nasional Indonesia—Ankatan Laut, Jawatan Hidro-Oseanografi: Jakarta, Indonesia, 1996; 411-KK. [Google Scholar]
- TNI-AL. Pulau-Pulau Seribu. Karang Tohorjantan Hingga Pulau Damar Besar, Sekala 1:50.000; Tentara Nasional Indonesia—Ankatan Laut, Jawatan Hidro-Oseanografi: Jakarta, Indonesia, 1985; 412-KK. [Google Scholar]
- TNI-AL. Pulau-Pulau Seribu. Pulau Tidung Besar Hingga Pulau Bokor, Sekala 1:50.000, 2nd ed.; Tentara Nasional Indonesia—Ankatan Laut, Jawatan Hidro-Oseanografi: Jakarta, Indonesia, 1998; 413-KK. [Google Scholar]
- TNI-AL. Pulau-Pulau Seribu. Pulau Jukung Hingga Pulau Piniki, Sekala 1:50.000, 2nd ed.; Tentara Nasional Indonesia—Ankatan Laut, Jawatan Hidro-Oseanografi: Jakarta, Indonesia, 1995; 414-KK. [Google Scholar]
- TNI-AL. Pulau-Pulau Seribu. Pulau Dua Barat hingga Pulau Tondan Timur, sekala 1:50.000, 2nd ed.; Tentara Nasional Indonesia—Ankatan Laut, Jawatan Hidro-Oseanografi: Jakarta, Indonesia, 1998; 415-KK. [Google Scholar]
- TNI-AL. Pulau-Pulau Seribu. Pulau Pramuka hingga Pulau Kotok Kecil, sekala 1:20.000; Tentara Nasional Indonesia—Ankatan Laut, Jawatan Hidro-Oseanografi: Jakarta, Indonesia, 1986; 416-KK. [Google Scholar]
- TNI-AL. Pulau-Pulau Seribu. Pulau Opak Kecil hingga Pulau Jukung, sekala 1:20.000, 2nd ed.; Tentara Nasional Indonesia—Ankatan Laut, Jawatan Hidro-Oseanografi: Jakarta, Indonesia, 1998; 417-KK. [Google Scholar]
- English, S.; Wilkinson, C.; Baker, V. Survey Manual for Tropical Marine Resources, 2nd ed.; Australian Institute of Marine Science: Townsville, Australia, 1997; pp. 1–390. [Google Scholar]
- Google Maps. Available online: https://www.google.com/maps/ (accessed on 12 January 2019).
- GPS Coordinates. Available online: https://gps-coordinates.org (accessed on 16 January 2019).
- Giyanto; Tuti, Y.; Budiyanto, A. Preliminary analysis on the condition of coral reefs in the Kepulaan Seribu, Jakarta 2005. In Coral Reef Ecosystem in the Seribu Islands: Workshop Monitoring and Evaluation of Three Decades; Tuti, Y., Soemodihardjo, S., Eds.; LIPI Press: Jakarta, Indonesia, 2006; pp. 9–17. [Google Scholar]
- Hoeksema, B.W.; Koh, E.G.L. Depauperation of the mushroom coral fauna (Fungiidae) of Singapore (1860s–2006) in changing reef conditions. Raffles Bull. Zool. 2009, 22, 91–101. [Google Scholar]
- Veron, J.E.N. Corals of the World; Australian Institute of Marine Science: Townsville, Australia, 2000; Volume 2, pp. 1–429. [Google Scholar]
- Benzoni, F.; Stefani, F.; Stolarski, J.; Pichon, M.; Mitta, G.; Galli, P. Debating phylogenetic relationships of the scleractinian Psammocora: Molecular and morphological evidences. Contrib. Zool. 2007, 76, 35–54. [Google Scholar]
- MINITAB. Minitab 17.1.0; Minitab Inc.: State College, PA, USA, 2013. [Google Scholar]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples, Version 9.1; User’s Guide and Application. 2013. Available online: http://viceroy.eeb.uconn.edu/estimates (accessed on 20 December 2018).
- Verwey, J. Coral reef studies. II. The depth of coral reefs in relation to their oxygen consumption and the penetration of light in the water. Treubia 1931, 13, 169–198. [Google Scholar]
- Harger, J.R.E. Community displacement in stressed coral reef systems and the implication for a comprehensive management strategy for coastal and offshore productivity enrichment. Galaxea 1988, 7, 185–196. [Google Scholar]
- Tomascik, T.; Suharsono; Mah, A.J. Case histories: A historical perspective of the natural and anthropogenic impacts in the Indonesian archipelago with a focus on Kepulauan Seribu, Java Sea. In Proceedings of the Colloquium on Global Aspects of Coral Reefs, Health, Hazards and History; Ginsburg, R., Ed.; University of Miami, Rosenstiel School of Marine and Atmospheric Science: Miami, FL, USA, 1993; pp. 304–310. [Google Scholar]
- Johan, O.; Zamany, N.P.; Smith, D.; Sweet, M.J. Prevalence and incidence of black band disease of scleractinian corals in the Kepulauan Seribu Region of Indonesia. Diversity 2016, 8, 11. [Google Scholar] [CrossRef]
- Baum, G.; Januar, H.I.; Ferse, S.C.A.; Wild, C.; Kunzmann, A. Abundance and physiology of dominant soft corals linked to water quality in Jakarta Bay, Indonesia. PeerJ 2016, 4, e2625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleary, D.F.R.; Polónia, A.R.M.; Renema, W.; Hoeksema, B.W.; Rachello-Dolmen, P.G.; Moolenbeek, R.G.; Budiyanto, A.; Yahmantoro; Tuti, Y.; Draisma, S.G.A.; et al. Variation in the composition of corals, fishes, sponges, echinoderms, ascidians, molluscs, foraminifera and macroalgae across a pronounced in-to-offshore environmental gradient in the Jakarta Bay–Thousand Islands coral reef complex. Mar. Pollut. Bull. 2016, 110, 701–717. [Google Scholar] [CrossRef]
- Draisma, S.G.A.; Prud'homme van Reine, W.F.; Herandarudewi, S.M.C.; Hoeksema, B.W. Macroalgal diversity along an inshore-offshore environmental gradient in the Jakarta Bay–Thousand Islands reef complex, Indonesia. Estuar. Coast. Shelf Sci. 2018, 200, 258–269. [Google Scholar] [CrossRef]
- Madduppa, H.H.; Subhan, B.; Suparyani, E.; Siregar, A.M.; Arafat, D.; Tarigan, S.A.; Alimuddin; Khairudi, D.; Rahmawati, F.; Bramandito, A. Dynamics of fish diversity across an environmental gradient in the Seribu Islands reefs off Jakarta. Biodiversitas 2013, 14, 17–24. [Google Scholar] [CrossRef]
- De Voogd, N.J.; Cleary, D.F.R. An analysis of sponge diversity and distribution at three taxonomic levels in the Thousand Islands/Jakarta bay reef complex, West-Java, Indonesia. Mar. Ecol. 2008, 29, 205–215. [Google Scholar] [CrossRef]
- Siregar, T.H.; Priyanto, N.; Putri, A.K.; Rachmawati, N.; Triwibowo, R.; Dsikowitzky, L.; Schwarzbauer, J. Spatial distribution and seasonal variation of the trace hazardous element contamination in Jakarta Bay, Indonesia. Mar. Pollut. Bull. 2016, 110, 634–646. [Google Scholar] [CrossRef]
- Alder, J.; Sloan, N.A.; Uktolseya, H.A. comparison of management planning and implementation in three Indonesian marine protected areas. Ocean Coast. Manag. 1994, 24, 179–198. [Google Scholar] [CrossRef]
- Farhan, A.R.; Lim, S. Vulnerability assessment of ecological conditions in Seribu Islands, Indonesia. Ocean Coast. Manag. 2012, 65, 1–14. [Google Scholar] [CrossRef]
- Erdmann, M.V. Destructive fishing practices in the Pulau Seribu Archipelego. In Proceedings of the Coral Reef evaluation workshop Pulau Seribu, Jakarta, Indonesia, 11–20 September 1995; Soemodihardjo, S., Ed.; Contending with Global Change 10. UNESCO: Jakarta, Indonesia, 1998; pp. 84–89. [Google Scholar]
- Fox, H.E.; Caldwell, R.L. Recovery from blast fishing on coral reefs: A tale of two scales. Ecol. Appl. 2006, 16, 1631–1635. [Google Scholar] [CrossRef]
- Raymundo, L.J.; Maypa, A.P.; Gomez, E.D.; Cadiz, P. Can dynamite-blasted reefs recover? A novel, low-tech approach to stimulating natural recovery in fish and coral populations. Mar. Pollut. Bull. 2007, 54, 1009–1019. [Google Scholar] [CrossRef]
- Williams, S.L.; Sur, C.; Janetski, N.; Hollarsmith, J.A.; Rapi, S.; Barron, L.; Heatwole, S.J.; Yusuf, A.M.; Yusuf, S.; Jompa, J.; et al. Large-scale coral reef rehabilitation after blast fishing in Indonesia. Restor. Ecol. 2018. [Google Scholar] [CrossRef]
- Fox, H.E.; Harris, J.L.; Darling, E.S.; Ahmadia, G.N.; Estradivari; Razak, T.B. Rebuilding coral reefs: Success (and failure) 16 years after low-cost, low-tech restoration. Restor. Ecol. 2019. [Google Scholar] [CrossRef]
- Hoeksema, B.W. Mobility of free-living fungiid corals (Scleractinia), a dispersion mechanism and survival strategy in dynamic reef habitats. In Proceedings of the 6th International Coral Reef Symposium, Townsville, Australia, 8–12 August 1988; Volume 2, pp. 715–720. [Google Scholar]
- Best, M.B.; Hoeksema, B.W.; Moka, W.; Moll, H.; Suharsono; Sutarna, I.N. Recent scleractinian coral species collected during the Snellius-II Expedition in eastern Indonesia. Neth. J. Sea Res. 1989, 23, 107–115. [Google Scholar] [CrossRef]
- Huang, D.; Licuanan, W.Y.; Hoeksema, B.W.; Chen, C.A.; Ang, P.O.; Huang, H.; Lane, D.J.W.; Vo, S.T.; Waheed, Z.; Amri, A.Y.; et al. Extraordinary diversity of reef corals in the South China Sea. Mar. Biodivers. 2015, 45, 157–168. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Waheed, Z. Initial phase of autotomy in fragmenting Cycloseris corals at Semporna, eastern Sabah, Malaysia. Coral Reefs 2011, 30, 1087. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Bouwmeester, J.; Range, P.; Ben-Hamadou, R.A. large aggregation of self-fragmenting mushroom corals in the Arabian/Persian Gulf. Ecology 2018, 99, 1236–1238. [Google Scholar] [CrossRef] [PubMed]
- Schuhmacher, H. Ability in fungiid corals to overcome sedimentation. In Proceedings of the 3rd International Coral Reef Symposium, Miami, 1977; pp. 503–509. [Google Scholar]
- Hoeksema, B.W.; Yeemin, T. Late detachment conceals serial budding by the free-living coral Fungia fungites in the Inner Gulf of Thailand. Coral Reefs 2011, 30, 975. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Waheed, Z. Onset of autotomy in an attached Cycloseris coral. Galaxea J. Coral Reef Stud. 2012, 14, 1–2. [Google Scholar] [CrossRef]
- Bongaerts, P.; Hoeksema, B.W.; Hay, K.B.; Hoegh-Guldberg, O. Mushroom corals overcome live burial through pulsed inflation. Coral Reefs 2012, 31, 399. [Google Scholar] [CrossRef]
- Chadwick-Furman, N.E.; Loya, Y. Migration, habitat use, and competition among mobile corals (Scleractinia: Fungiidae) in the Gulf of Eilat, Red Sea. Mar. Biol. 1992, 114, 617–623. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; de Voogd, N.J. On the run: Free-living mushroom corals avoiding interaction with sponges. Coral Reefs 2012, 31, 455–459. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Bongaerts, P. Mobility and self-righting by a freeliving mushroom coral through pulsed inflation. Mar. Biodivers. 2016, 46, 521–524. [Google Scholar] [CrossRef]
- Plusquellec, Y.; Webb, G.; Hoeksema, B.W. Automobility in Tabulata, Rugosa, and extant scleractinian analogues: Stratigraphic and paleogeographic distribution of Paleozoic mobile corals. J. Paleontol. 1999, 73, 985–1001. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Hassell, D.; Meesters, E.H.W.G.; van Duyl, F.C. Wave-swept coralliths of Saba Bank, Dutch Caribbean. Mar. Biodivers. 2018, 48, 2003–2016. [Google Scholar] [CrossRef]
- Denis, V.; De Palmas, S.; Benzoni, F.; Chen, C.C. Extension of the known distribution and depth range of the scleractinian coral Psammocora stellata: First record from a Taiwanese mesophotic reef. Mar. Biodivers. 2015, 45, 619–620. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Coordinates, DD | Shelf Zone | Number of Species | Shallow Records | Shelf Depth, m | Secchi Depth, m | Nearest Mainland, km | Nearest River Outlet, km | Distance to Harbor, km |
|---|---|---|---|---|---|---|---|---|---|
| Onrust NW | −6.0334, 106.7328 | 1 | 0 | 0 | 9 | 1.5 | 2.3 | 2.3 | 17.3 |
| Bidadari NW | −6.0321, 106.7463 | 1 | 5 | 5 | 11 | 1.5 | 3.6 | 3.7 | 16.0 |
| Nyamuk Besar NW | −6.0291, 106.8523 | 1 | 2 | 2 | 18 | 3.5 | 7.6 | 12.9 | 7.6 |
| Kelor NW | −6.0253, 106.7441 | 1 | 3 | 3 | 12 | 2.5 | 3.8 | 3.8 | 16.5 |
| Ayer Besar NW | −6.0017, 106.7801 | 1 | 1 | 1 | 18 | 3.5 | 8.6 | 8.6 | 14.8 |
| Ubi Besar NW | −5.9987, 106.7397 | 1 | 3 | 2 | 15 | 4.0 | 5.5 | 5.8 | 18.5 |
| Damar Kecil NW | −5.9834, 106.8453 | 1 | 5 | 4 | 19 | 3.5 | 12.7 | 15.9 | 12.7 |
| Untung Jawa NW | −5.9741, 106.7031 | 1 | 10 | 9 | 20 | 2.5 | 5.0 | 6.6 | 23.4 |
| Damar Besar NW | −5.8547, 106.8409 | 1 | 10 | 10 | 28 | 8.0 | 15.4 | 15.4 | 15.9 |
| Dapur NW | −5.9457, 106.7242 | 2 | 17 | 14 | 28 | 7.5 | 8.9 | 10.3 | 23.6 |
| Bokor NW | −5.9430, 106.6271 | 1 | 10 | 9 | 20 | 5.0 | 6.7 | 6.7 | 32.2 |
| Lancang NW | −5.9270, 106.5913 | 1 | 5 | 5 | 21 | 5.0 | 9.5 | 9.5 | 36.6 |
| Pari S shoal | −5.8808, 106.6352 | 2 | 15 | - | 29 | 8.0 | 12.0 | 13.6 | 35.7 |
| Tikus S | −5.8653, 106.5819 | 2 | 15 | 10 | 32 | 6.5 | 15.5 | 16.3 | 41.3 |
| Tikus W | −5.8589, 106.5700 | 2 | 14 | 9 | 37 | 8.0 | 16.8 | 17.4 | 42.8 |
| Pari E | −5.8544, 106.6389 | 2 | 17 | 8 | 37 | 12.5 | 14.8 | 16.6 | 37.4 |
| Tikus N | −5.8536, 106.5786 | 2 | 17 | 12 | 43 | 11.0 | 16.9 | 17.6 | 42.4 |
| Karang Jong E | −5.8522, 106.6486 | 2 | 20 | 11 | 34 | 11.5 | 15.0 | 16.9 | 36.8 |
| Karang Jong NW | −5.8511, 106.6464 | 2 | 19 | 12 | 48 | 11.0 | 15.1 | 17.0 | 37.1 |
| Tikus NE | −5.8501, 106.5848 | 2 | 18 | 11 | 34 | 8.5 | 16.9 | 17.8 | 42.1 |
| Payung Besar E | −5.8219, 106.5631 | 2 | 15 | 11 | 54 | 10.5 | 19.4 | 21.5 | 46.0 |
| Payung Kecil NW | −5.8134, 106.5492 | 2 | 19 | 11 | 54 | 15.5 | 20.1 | 23.0 | 47.8 |
| Tidung Kecil NW | −5.7997, 106.5178 | 2 | 21 | 13 | 45 | 17.5 | 21.7 | 25.9 | 51.4 |
| Tidung Besar NW | −5.7910, 106.4812 | 2 | 17 | 10 | 43 | 15.0 | 23.3 | 28.8 | 55.2 |
| Air NW | −5.7606, 106.7456 | 3 | 15 | 11 | 29 | 12.0 | 27.3 | 28.9 | 39.7 |
| Semak Daun NW | −5.7322, 106.5731 | 3 | 16 | 11 | 28 | 11.5 | 29.6 | 30.8 | 52.4 |
| Kotok Besar NW | −5.6988, 106.5398 | 3 | 16 | 12 | 33 | 10.5 | 32.8 | 35.3 | 57.6 |
| Peniki NW | −5.6968, 106,7155 | 2 | 23 | 12 | 43 | 9.5 | 32.3 | 34.9 | 47.6 |
| Peniki E | −5.6922, 106.7174 | 2 | 21 | 11 | 41 | 9.5 | 33.6 | 35.4 | 48.0 |
| Kotok Kecil NW | −5.6888, 106.5336 | 3 | 19 | 15 | 25 | 10.0 | 33.9 | 36.6 | 58.9 |
| Kelapa, NW | −5.6544, 106.5589 | 3 | 15 | 13 | 28 | 11.0 | 37.8 | 39.6 | 60.2 |
| Panjang, NW | −5.6423, 106.5599 | 3 | 14 | 12 | 26 | 11.5 | 39.2 | 40.9 | 61.2 |
| Belanda, NW | −5.6037, 106.6035 | 3 | 19 | 12 | 31 | 10.0 | 43.0 | 44.5 | 62.3 |
| Putri, NW | −5.5904, 106.5673 | 3 | 15 | 11 | 26 | 10.5 | 45.0 | 46.4 | 65.6 |
| Sepa, NW | −5.5755, 106.5799 | 3 | 15 | 13 | 29 | 11.0 | 46.4 | 47.9 | 66.3 |
| Jukung, NW | −5.5669, 106.5272 | 3 | 11 | 10 | 30 | 9.5 | 47.5 | 49.9 | 70.2 |
| Hantu Kecil, NW | −5.5354, 106.5319 | 3 | 19 | 15 | 31 | 10.0 | 51.0 | 53.2 | 72.3 |
| Hantu Besar, NW | −5.5296, 106.5389 | 3 | 17 | 14 | 27 | 9.5 | 51.6 | 53.6 | 73.0 |
| Species | Zone 1 11 Sites | Zone 2 15 Sites | Zone 3 12 Sites | All Three Zones 38 Sites |
|---|---|---|---|---|
| Ctenactis albitentaculata (Hoeksema, 1989) | 1 (1) | 10 (2) | 8 (5) | 19 (8) |
| Ctenactis crassa (Dana, 1846) | 8 | 9 (6) | 17 (6) | |
| Ctenactis echinata (Pallas, 1766) | 4 (4) | 14 (13) | 12 (12) | 30 (29) |
| Cycloseris boschmai (Hoeksema, 2014) # | 2 | 2 | ||
| Cycloseris costulata (Ortmann, 1889) | 3 (2) | 11 | 1 | 15 (2) |
| Cycloseris cyclolites (Lamarck, 1816) # | 3 | 3 | ||
| Cycloseris explanulata (van der Horst, 1922) # | 3 | 1 | 4 | |
| Cycloseris fragilis (Alcock, 1893) | 10 | 1 | 11 | |
| Cycloseris mokai (Hoeksema, 1989) | 7 | 4 | 11 | |
| Cycloseris sinensis (Milne Edwards and Haime, 1851) # | 1 | 2 | 3 | |
| Cycloseris tenuis (Dana, 1846) # | 1 | 1 | ||
| Cycloseris vaughani (Boschma, 1923) # | 1 | 1 | ||
| Danafungia horrida (Dana, 1846) | 2 (2) | 15 (14) | 12 (12) | 29 (28) |
| Danafungia scruposa (Klunzinger, 1879) | 2 (2) | 13 (12) | 11 (11) | 26 (25) |
| Fungia fungites (Linnaeus, 1758) | 6 (6) | 15 (14) | 12 (12) | 33 (32) |
| Halomitra pileus (Linnaeus, 1758) | 2 (2) | 2 (1) | 4 (3) | |
| Heliofungia actiniformis (Quoy and Gaimard, 1833) | 2 (2) | 10 (2) | 8 (6) | 20 (10) |
| Herpolitha limax (Esper, 1797) | 3 (1) | 15 (14) | 12 (12) | 30 (27) |
| Lithophyllon concinna (Verrill, 1864) | 2 (2) | 15 (14) | 12 (12) | 29 (28) |
| Lithophyllon repanda (Dana, 1846) | 3 (3) | 15 (14) | 12 (12) | 30 (29) |
| Lithophyllon scabra (Döderlein, 1901) | 1 * | 1 * | ||
| Lithophyllon undulatum Rehberg, 1892 | 5 (5) | 7 (1) | 5 (1) | 17 (7) |
| Lobactis scutaria (Lamarck, 1801) | 3 (1) | 3 (1) | ||
| Pleuractis granulosa (Klunzinger, 1879) | 10 (5) | 6 (5) | 16 (10) | |
| Pleuractis moluccensis (van der Horst, 1919) | 3 (2) | 15 (1) | 10 | 28 (3) |
| Pleuractis paumotensis (Stutchbury, 1833) | 5 (5) | 15 (14) | 12 (12) | 32 (31) |
| Podabacia crustacea (Pallas, 1766) | 1 (1) | 13 (8) | 12 (12) | 26 (21) |
| Podabacia kunzmanni (Hoeksema, 2009) # | (1)** | (1)** | ||
| Polyphyllia talpina (Lamarck, 1801) | 4 (4) | 11 (11) | 7 (7) | 22 (22) |
| Sandalolitha dentata Quelch, 1884 | 9 | 8 (1) | 17 (1) | |
| Sandalolitha robusta (Quelch, 1886) | 8 (8) | 14 (12) | 12 (11) | 34 (31) |
| Total number of species per zone/three zones | 16 (16) | 29 (18) + 1 * | 24 (18) + 1 ** | 29 (21) + 2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoeksema, B.W.; Giyanto; Suharsono. The Role of Maximum Shelf Depth versus Distance from Shore in Explaining a Diversity Gradient of Mushroom Corals (Fungiidae) off Jakarta. Diversity 2019, 11, 46. https://doi.org/10.3390/d11030046
Hoeksema BW, Giyanto, Suharsono. The Role of Maximum Shelf Depth versus Distance from Shore in Explaining a Diversity Gradient of Mushroom Corals (Fungiidae) off Jakarta. Diversity. 2019; 11(3):46. https://doi.org/10.3390/d11030046
Chicago/Turabian StyleHoeksema, Bert W., Giyanto, and Suharsono. 2019. "The Role of Maximum Shelf Depth versus Distance from Shore in Explaining a Diversity Gradient of Mushroom Corals (Fungiidae) off Jakarta" Diversity 11, no. 3: 46. https://doi.org/10.3390/d11030046
APA StyleHoeksema, B. W., Giyanto, & Suharsono. (2019). The Role of Maximum Shelf Depth versus Distance from Shore in Explaining a Diversity Gradient of Mushroom Corals (Fungiidae) off Jakarta. Diversity, 11(3), 46. https://doi.org/10.3390/d11030046