A Surfeit of Studies: What Have We Learned from All the Box Turtle (Terrapene carolina and T. ornata) Home Range Studies?

Abstract
:1. Introduction
2. Method—Meta-Analysis Outline
Meta-Analysis
3. Results
3.1. Data Set
3.2. Analysis
4. Discussion
4.1. HR of T. Carolina and T. Ornata
4.2. Do the HRs of Male and Female Terrapene Differ in size?
4.3. Is HR Influenced by Ecoregion?
4.4. How Does Choice of Analysis Technique Affect HR Estimate?
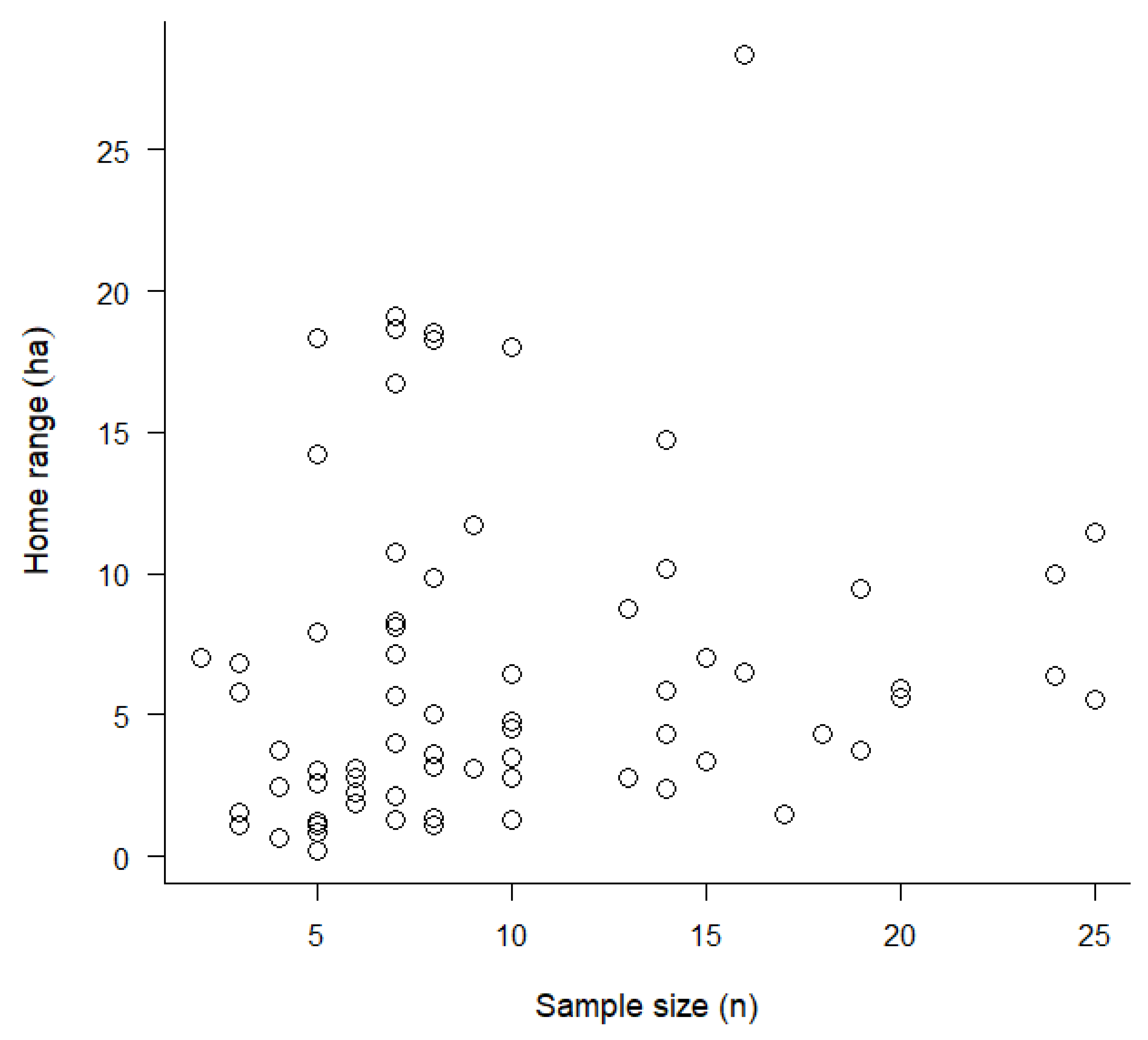
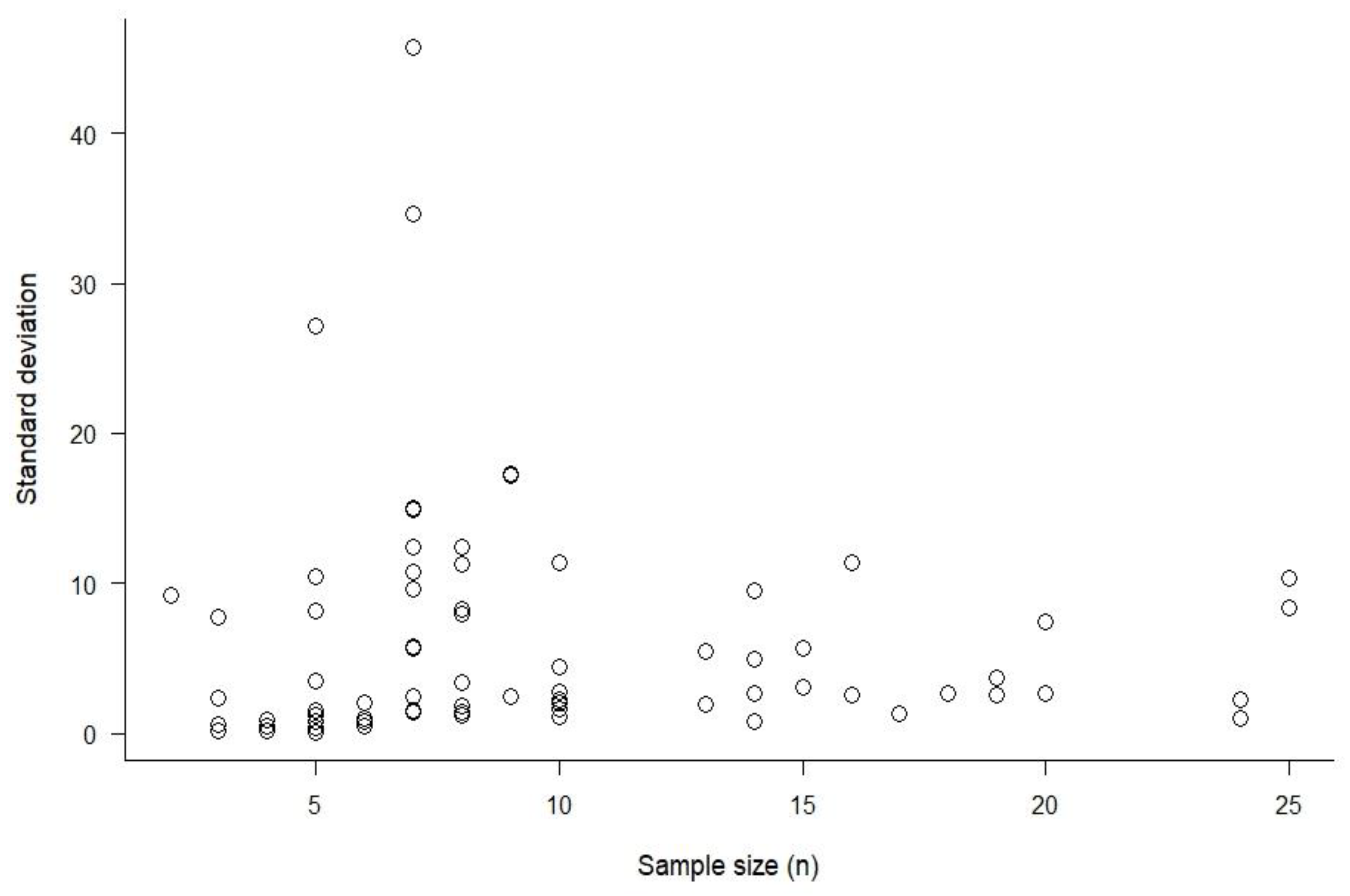
4.5. Does Sample Size Affect HR Size Estimation?
4.6. Do Relocated Terrapene Have Larger HR?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Burt, W.H. Territoriality and home range concepts as applied to mammals. J. Mammal. 1943, 24, 346. [Google Scholar] [CrossRef]
- Kernohan, B.J.; Gitzen, R.A.; Millspaugh, J.J. Analysis of animal space use and movements. In Radio Tracking and Animal Populations; Millspaugh, J.J., Marzluff, J.M., Eds.; Academic Press: London, UK, 2001; pp. 126–164. [Google Scholar]
- Van Winkle, W. Comparison of several probabilistic home-range models. J. Wildl. Manag. 1975, 39, 118–123. [Google Scholar] [CrossRef]
- Jones, R.L. Home range and seasonal movements of the Turtle Graptemys flavimaculata. J. Herpetol. 1996, 30, 376. [Google Scholar] [CrossRef]
- Morrow, J.L.; Howard, J.H.; Smith, S.A.; Poppel, D.K. Home range and movements of the bog turtle (Clemmys muhlenbergii) in Maryland. J. Herpetol. 2001, 35, 68–73. [Google Scholar] [CrossRef]
- Arvisais, M.; Bourgeois, J.-C.; Daigle, C.; Jutras, J.; Lévesque, E.; Masse, D. Home range and movements of a wood turtle (Clemmys insculpta) population at the northern limit of its range. Can. J. Zool. 2002, 80, 402–408. [Google Scholar] [CrossRef]
- Dodd, C.K. North American Box Turtles: A Natural History; Oklahoma University Press: Norman, OK, USA, 2001. [Google Scholar]
- Lindeman, P.V. The Map Turtle and Sawback Atlas; University of Oklahoma Press: Norman, OK, USA, 2013. [Google Scholar]
- Kimble, S.J.A.; Rhodes, O.E., Jr.; Williams, R.N. Unexpectedly Low Rangewide Population Genetic Structure of the Imperiled Eastern Box Turtle Terrapene c. carolina. PLoS ONE 2014, 9, e92274. [Google Scholar] [CrossRef]
- Breder, R.B. Turtle trailing: A new technique for studying the life habits of certain Testudinata. Zoologica 1927, 9, 231–243. [Google Scholar]
- Nichols, J.T. Range and homing of individual box turtles. Copeia 1939, 1939, 125–127. [Google Scholar] [CrossRef]
- Powell, R.; Conant, R.; Collins, J.T. Peterson Field Guide to Reptiles and Amphibians of Eastern and Central North America, 4th ed.; Houghton Mifflin Harcourt: Boston, MA, USA, 2016. [Google Scholar]
- Lue, K.-Y.; Chen, T.-H. Activity, movement patterns, and home range of the yellow-margined box turtle (Cuora flavomarginata) in Northern Taiwan. J. Herpetol. 1999, 33, 590–600. [Google Scholar] [CrossRef]
- Lawson, D.P. Habitat use, home range, and activity patterns of hingeback tortoises, Kinixys erosa and K. homeana, in Southwestern Cameroon. Chelonian Conser. Biol. 2006, 5, 48–56. [Google Scholar] [CrossRef]
- Fasola, M.; Mazzotti, S.; Pisapia, A. Activity and home range of Testudo hermanni in Northern Italy. Amphibia-Reptilia 2002, 23, 305–312. [Google Scholar] [CrossRef]
- Row, J.R.; Blouin-Demers, G. Kernels are not accurate estimators of home-range size for herpetofauna. Copeia 2006, 2006, 797–802. [Google Scholar] [CrossRef]
- Harless, M.L.; Walde, A.D.; Delaney, D.K.; Pater, L.L.; Hayes, W.K. Sampling considerations for improving home range estimates of desert tortoises: Effects of estimator, sampling regime, and sex. Herpetol. Conserv. Biol. 2010, 5, 374–387. [Google Scholar]
- Worton, B.J. A review of models of home range for animal movement. Ecol. Model. 1987, 38, 277–298. [Google Scholar] [CrossRef]
- Boyle, S.A.; Lourenço, W.C.; Da Silva, L.R.; Smith, A.T. Home range estimates vary with sample size and methods. Folia Primatol. 2009, 80, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Powell, R.A. Animal home ranges and territories and home range estimators. In Research Techniques in Animal Ecology: Controversies and Consequences; Boitani, L., Fuller, T.K., Eds.; Columbia University Press: New York, NY, USA, 2000; pp. 65–110. [Google Scholar]
- Farnsworth, S.D.; Seigel, R.A. Responses, movements, and survival of relocated box turtles during construction of the intercounty connector highway in Maryland. Transp. Res. Rec. J. Transp. Res. Board 2013, 2362, 1–8. [Google Scholar] [CrossRef]
- Sosa, J.A.; Perry, G. Site fidelity, movement, and visibility following translocation of Ornate box turtles (Terrapene ornata ornata) from a wildlife rehabilitation center in the high plains of Texas. Herpetol. Conserv. Biol. 2015, 10, 255–262. [Google Scholar]
- EPA. Environmental Protection Agency Ecoregions. 2018. Available online: https://www.epa.gov/eco-research/ecoregions (accessed on 30 March 2018).
- Viechtbauer, W. Conducting Meta-Analyses in R with the metafor Package. J. Stat. Softw. 2010, 36, 1–48. [Google Scholar] [CrossRef]
- R Core Team. 2015 R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: http://www.R-project.org/ (accessed on 11 October 2018).
- R Studio Team. 2016 R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: http://www.R-project.org/ (accessed on 11 October 2018).
- Nakagawa, S.; Santos, E.S.A. Methodological issues and advances in biological meta-analysis. Evol. Ecol. 2012, 26, 1253–1274. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P.; Heiberger, R.M.; Schuetzenmeister, A.; Scheibe, S. Package “Multcomp”. 2019. Available online: http://cran.stat.sfu.701ca/web/packages/multcomp/multcomp.pdf/ (accessed on 11 October 2018).
- Hedges, L.V.; Olkin, I. Statistical Methods for Meta-Analysis; Academic Press: Orlando, FL, USA, 1985. [Google Scholar]
- Egger, M.; Smith, G.D.; Schneider, M.; Minder, C. Bias in meta-analysis detected by a simple, graphical test. BMJ 1997, 315, 629–634. [Google Scholar] [CrossRef] [Green Version]
- Sterne, J.A.C.; Egger, M. Regression methods to detect publication and other bias in meta-analysis. In Publication Bias in Meta-Analysis: Prevention, Assessment, and Adjustments; Rothstein, H.R., Sutton, A.J., Borenstein, M., Eds.; John Wiley & Sons: Chichester, UK, 2005; pp. 99–110. [Google Scholar]
- Viechtbauer, W.; Cheung, M.W.-L. Outlier and influence diagnostics for meta-analysis. Synth. Methods 2010, 1, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Habeck, C.W.; Schultz, A.K. Community-level impacts of white-tailed deer on understory plants in North American forests: A meta-analysis. AoB Plants 2015, 7, plv119. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.P. Outliers and influential data points in regression analysis. Psychol. Bull. 1984, 95, 334–344. [Google Scholar] [CrossRef]
- Aguinis, H.; Gottfredson, R.K.; Joo, H. Best-practice recommendations for defining, identifying, and handling outliers. Organ Methods 2013, 16, 270–301. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-137. 2018. Available online: https://CRAN.R-project.org/package=nlme (accessed on 11 October 2018).
- Madden, R.C. Home Range, Movements, and Orientation in the Eastern Box Turtle, Terrapene carolina carolina. Ph.D. Thesis, City University of New York, New York, NY, USA, 1975. [Google Scholar]
- Davis, M. Aspects of the Social and Spatial Experience of Eastern Box Turtles, Terrapene carolina carolina. Ph.D. Thesis, University of Tennessee, Knoxville, TN, USA, 1981. [Google Scholar]
- Bayless, J.W. Home Range Studies of the Eastern Box Turtle (Terrapene carolina carolina) Using Radio Telemetry. Master’s Thesis, George Mason University, Fairfax, VA, USA, 1984. [Google Scholar]
- Holy, L.L. Home Range, Homing Ability, Orientation and Navigational Mechanisms of the Western Box Turtle (Terrapene ornata) from Western Nebraska. Ph.D. Thesis, University of Nebraska—Lincoln, Lincoln, NE, USA, 1995. [Google Scholar]
- Trail, C.D. Natural History and Habitat Use of the Ornate Box Turtle Terrapene ornata ornata at a Mixed-Grass Prairie in Southwest Nebraska. Master’s Thesis, University Nebraska, Omaha, NE, USA, 1995. [Google Scholar]
- Nieuwolt, P.M. Movement, activity, and microhabitat selection in the western box turtle, Terrapene ornata luteola, in New Mexico. Herpetologica 1996, 52, 487–495. [Google Scholar]
- Kaye, D.R.J.; Walsh, K.M.; Ross, C.M. Seasonal movements and habitat preferences for the spotted turtle and eastern box turtle in Massachusetts. In Proceedings of the 2001 International Conference on Ecology and Transportation, Keystone, CO, USA, 24–28 September 2001; Irwin, C.L., Garrett, P., McDermott, K.P., Eds.; Center for Transportation and the Environment, North Carolina State University: Raleigh, NC, USA, 2001; pp. 66–82. [Google Scholar]
- Hall, D.H.; Steidl, R.J. Ecology and Conservation of Desert Box Turtles (Terrapene ornata luteola) in Arizona; Project No. I02003; Arizona Game and Fish Department: Phoenix, AZ, USA, 2003.
- Cook, R.P. Dispersal, home range establishment, survival, and reproduction of translocated eastern box turtles, Terrapene c. carolina. Appl. Herpetol. 2004, 1, 197–228. [Google Scholar] [CrossRef]
- Capitano, W. Home Range, Spatial Patterning and Reproductive Ecology of Female Eastern Box Turtles in a Long Island Population. Master’s Thesis, Hofstra University, Hempstead, NY, USA, 2005. [Google Scholar]
- Donaldson, B.M.; Echternacht, A.C. Aquatic Habitat Use Relative to Home Range and Seasonal Movement of Eastern Box Turtles (Terrapene carolina carolina: Emydidae) in Eastern Tennessee. J. Herpetol. 2005, 39, 278–284. [Google Scholar] [CrossRef]
- Lentz, J. Home Range and Habitat Preferences of Terrapene carolina carolina at Jug Bay Wetlands Sanctuary, Maryland; Technical Report; Jug Bay Wetlands Sanctuary: Lothian, MD, USA, 2005. [Google Scholar]
- Bernstein, N.P.; Richtsmeier, R.J.; Black, R.W.; Montgomery, B.R. Home range and philopatry in the ornate box turtle, Terrapene ornata ornata, in Iowa. Am. Midl. Nat. 2007, 157, 162–174. [Google Scholar] [CrossRef]
- Rittenhouse, C.D.; Millspaugh, J.J.; Hubbard, M.W.; Sheriff, S.L. Movements of translocated and resident three-toed box turtles. J. Herpetol. 2007, 41, 115–121. [Google Scholar] [CrossRef]
- Hester, J.M.; Price, S.J.; Dorcas, M.E. Effects of relocation on movements and home ranges of eastern box turtles. J. Wildl. Manag. 2008, 72, 772–777. [Google Scholar] [CrossRef]
- Quinn, D.P. A Radio-Telemetric Study of the Eastern Box Turtle (Terrapene carolina carolina): Home-Range, Habitat Use, and Hibernacula Selection in Connecticut. Ph.D. Thesis, Central Connecticut State University, New Britain, CT, USA, 2008. [Google Scholar]
- Baker, J.M. Home Range and Movement of the Eastern Box Turtle (Terrapene carolina) in East Central Illinois. Master’s Thesis, University of Illinois at Urbana-Champaign, Champaign, IL, USA, 2009. [Google Scholar]
- Aall, N.C. Influence of Season and Sex on Terrapene c. carolina (Eastern Box Turtle) Movements: An Observation of a Population in West Virginia. Master’s Thesis, Marshall University, Huntington, WV, USA, 2011. [Google Scholar]
- Currylow, A.F.; MacGowan, B.J.; Williams, R.N. Short-term forest management effects on a long-lived ectotherm. PLoS ONE 2012, 7, e40473. [Google Scholar] [CrossRef] [PubMed]
- Refsnider, J.M.; Strickland, J.; Janzen, F.J. Home range and site fidelity of imperiled ornate box turtles (Terrapene ornata) in northwestern Illinois. Chelonian Conser. Biol. 2012, 11, 78–83. [Google Scholar] [CrossRef]
- Samuelson, C.S. Movement Patterns in Resident and Translocated Three-Toed Box Turtles (Terrapene carolina triunguis). Master’s Thesis, The University of Texas at Tyler, Tyler, TX, USA, 2012. [Google Scholar]
- Wilson, S.D. Movement and Ecology of the Eastern Box Turtle (Terrapene carolina carolina) in a Heterogeneous Landscape. Master’s Thesis, Bowling Green State University, Bowling Green, OH, USA, 2012. [Google Scholar]
- Kapfer, J.M.; Muñoz, D.J.; Groves, J.D.; Kirk, R.W. Home range and habitat preferences of Eastern Box Turtles (Terrapene carolina Linnaeus, 1758) in the Piedmont Ecological Province of North Carolina (USA). Herpetol. Notes 2013, 6, 251–260. [Google Scholar]
- Greenspan, S.E.; Condon, E.P.; Smith, L.L. Home range and habitat selection in the eastern box turtle (Terrapene carolina carolina) in a longleaf pine (Pinus palustris) reserve. Herpetol. Conserv. Biol. 2015, 10, 99–111. [Google Scholar]
- Omernik, J.M. Ecoregions of the conterminous United States. Ann. Assoc. Am. Geogr. 1987, 77, 118–125. [Google Scholar] [CrossRef]
- Clark, H.W. On the occurrence of a probable hybrid between the eastern and western box turtles, Terrapene carolina and T. ornata, near Lake Maxinkuckee, Indiana. Copeia 1935, 1935, 148–150. [Google Scholar] [CrossRef]
- Grobman, A.B. The effect of soil temperatures on emergence from hibernation of Terrapene carolina and T. ornata. Am. Midl. Nat. 1990, 124, 366. [Google Scholar] [CrossRef]
- Cureton, J.C.; Buchman, A.B.; Deaton, R.; Lutterschmidt, W.I. Molecular analysis of hybridization between the box turtles Terrapene carolina and T. ornata. Copeia 2011, 2011, 270–277. [Google Scholar] [CrossRef]
- Wanchai, P.; Stanford, C.B.; Thirakhupt, K.; Thanhikorn, S. Home range of the impressed tortoise, Manouria impressa (Günther, 1882) at Phu Luang Wildlife Sanctuary, Loei Province, Thailand. Trop. Nat. Hist. 2012, 12, 165–174. [Google Scholar]
- Barton, R.; Whiten, A.; Strum, S.; Byrne, R.; Simpson, A. Habitat use and resource availability in baboons. Anim. Behav. 1992, 43, 831–844. [Google Scholar] [CrossRef]
- Edwards, M.A.; DeRocher, A.E.; Nagy, J.A. Home range size variation in female arctic grizzly bears relative to reproductive status and resource availability. PLoS ONE 2013, 8, e68130. [Google Scholar] [CrossRef]
- Schoepf, I.; Schmohl, G.; König, B.; Pillay, N.; Schradin, C. Manipulation of population density and food availability affects home range sizes of African striped mouse females. Anim. Behav. 2015, 99, 53–60. [Google Scholar] [CrossRef]
- Laver, P.N.; Kelly, M.J. A critical review of home range studies. J. Wildl. Manag. 2008, 72, 290–298. [Google Scholar] [CrossRef]
- Kie, J.G.; Matthiopoulos, J.; Fieberg, J.; Powell, R.A.; Cagnacci, F.; Mitchell, M.S.; Gaillard, J.-M.; Moorcroft, P.R. The home-range concept: Are traditional estimators still relevant with modern telemetry technology? Philos. Trans. Soc. B Biol. Sci. 2010, 365, 2221–2231. [Google Scholar] [CrossRef]
- Powell, R.A.; Mitchell, M.S. What is a home range? J. Mammal. 2012, 93, 948–958. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.M. Repatriation and Health Assessment of Eastern Box Turtles (Terrapene c. carolina) at Caumsett State Park, New York. Master’s Thesis, Hofstra University, Hempstead, NY, USA, 2004. [Google Scholar]
- Tuberville, T.D.; Clark, E.E.; Buhlmann, K.A.; Gibbons, J.W. Translocation as a conservation tool: Site fidelity and movement of repatriated gopher tortoises (Gopherus polyphemus). Anim. Conserv. 2005, 8, 349–358. [Google Scholar] [CrossRef]
- Nussear, K.E.; Tracy, C.R.; Medica, P.A.; Wilson, D.S.; Marlow, R.W.; Corn, P.S. Translocation as a conservation tool for Agassiz’s desert tortoises: Survivorship, reproduction, and movements. J. Wildl. Manag. 2012, 76, 1341–1353. [Google Scholar] [CrossRef]
- Cochrane, M.M.; Brown, D.J.; Moen, R.A. GPS technology for semi-aquatic turtle research. Diversity 2019, 11, 34. [Google Scholar] [CrossRef]
- Thompson, D.G.; Swystun, T.; Cross, J.; Cross, R.L.; Chartrand, D.; Edge, C.B. Fine- and coarse-scale movements and habitat use by Wood Turtles (Glyptemys insculpta) based on probabilistic modeling of radiotelemetry and GPS-telemetry data. Can. J. Zool. 2018, 96, 1153–1164. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | HR Estimate | SE | CI.L | CI.U |
|---|---|---|---|---|
| T. carolina | ||||
| Overall | 4.50 | 0.83 | 2.88 | 6.12 |
| Female a | 4.82 | 1.24 | 2.40 | 7.24 |
| Male b | 3.80 | 1.24 | 1.37 | 6.22 |
| T. ornata | ||||
| Overall | 4.61 | 1.37 | 1.93 | 7.29 |
| Female a | 4.37 | 1.63 | 1.18 | 7.56 |
| Male b | 4.67 | 1.63 | 1.48 | 7.86 |
| Ecoregions | HR Estimate | SE | CI.L | CI.U |
|---|---|---|---|---|
| Central USA plains | 18.32 | 12.43 | −6.05 | 42.69 |
| Mississippi alluvial and southeast USA coastal plains | 4.64 | 1.71 | 1.29 | 8.00 |
| Mixed wood plains | 3.51 | 2.06 | −0.54 | 7.55 |
| Ozark/Ouachita-Appalachian forests | 2.04 | 1.53 | −0.96 | 5.05 |
| Southeastern USA plains | 5.16 | 0.91 | 3.37 | 6.95 |
| Ecoregions | HR Estimate | SE | CI.L | CI.U |
|---|---|---|---|---|
| Central USA plains a | 2.55 | 1.00 | 0.58 | 4.52 |
| Cold desert a | 1.58 | 1.02 | −0.42 | 3.57 |
| South-central semi-arid prairies s | 2.32 | 1.01 | 0.35 | 4.30 |
| Temperate prairies a | 0.86 | 0.99 | −1.08 | 2.79 |
| West-central semi-arid prairies b | 18.81 | 3.02 | 12.90 | 24.72 |
| Western Sierra Madre piedmont c | 6.97 | 1.01 | 5.00 | 8.94 |
| HR Calculation Technique | HR Estimate | SE | CI.L | CI.U |
|---|---|---|---|---|
| Ellipse a | 8.29 | 0.91 | 6.50 | 10.08 |
| Harmonic mean b | 4.13 | 0.83 | 2.50 | 5.75 |
| Kernel bc | 3.48 | 0.72 | 2.08 | 4.88 |
| MCP bd | 4.59 | 0.70 | 3.21 | 5.96 |
| Relocated | HR Estimate | SE | CI.L | CI.U |
|---|---|---|---|---|
| Yes a | 6.13 | 0.80 | 4.56 | 7.70 |
| No b | 4.07 | 0.67 | 2.76 | 5.38 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Habeck, C.W.; Figueras, M.P.; Deo, J.E.; Burke, R.L. A Surfeit of Studies: What Have We Learned from All the Box Turtle (Terrapene carolina and T. ornata) Home Range Studies? Diversity 2019, 11, 68. https://doi.org/10.3390/d11050068
Habeck CW, Figueras MP, Deo JE, Burke RL. A Surfeit of Studies: What Have We Learned from All the Box Turtle (Terrapene carolina and T. ornata) Home Range Studies? Diversity. 2019; 11(5):68. https://doi.org/10.3390/d11050068
Chicago/Turabian StyleHabeck, Christopher W., Miranda P. Figueras, Jean E. Deo, and Russell L. Burke. 2019. "A Surfeit of Studies: What Have We Learned from All the Box Turtle (Terrapene carolina and T. ornata) Home Range Studies?" Diversity 11, no. 5: 68. https://doi.org/10.3390/d11050068
APA StyleHabeck, C. W., Figueras, M. P., Deo, J. E., & Burke, R. L. (2019). A Surfeit of Studies: What Have We Learned from All the Box Turtle (Terrapene carolina and T. ornata) Home Range Studies? Diversity, 11(5), 68. https://doi.org/10.3390/d11050068