Utility of Condition Indices as Predictors of Lipid Content in Slimy Sculpin (Cottus cognatus)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Condition Indices and Lipid Extraction
2.2. Statistical Analyses
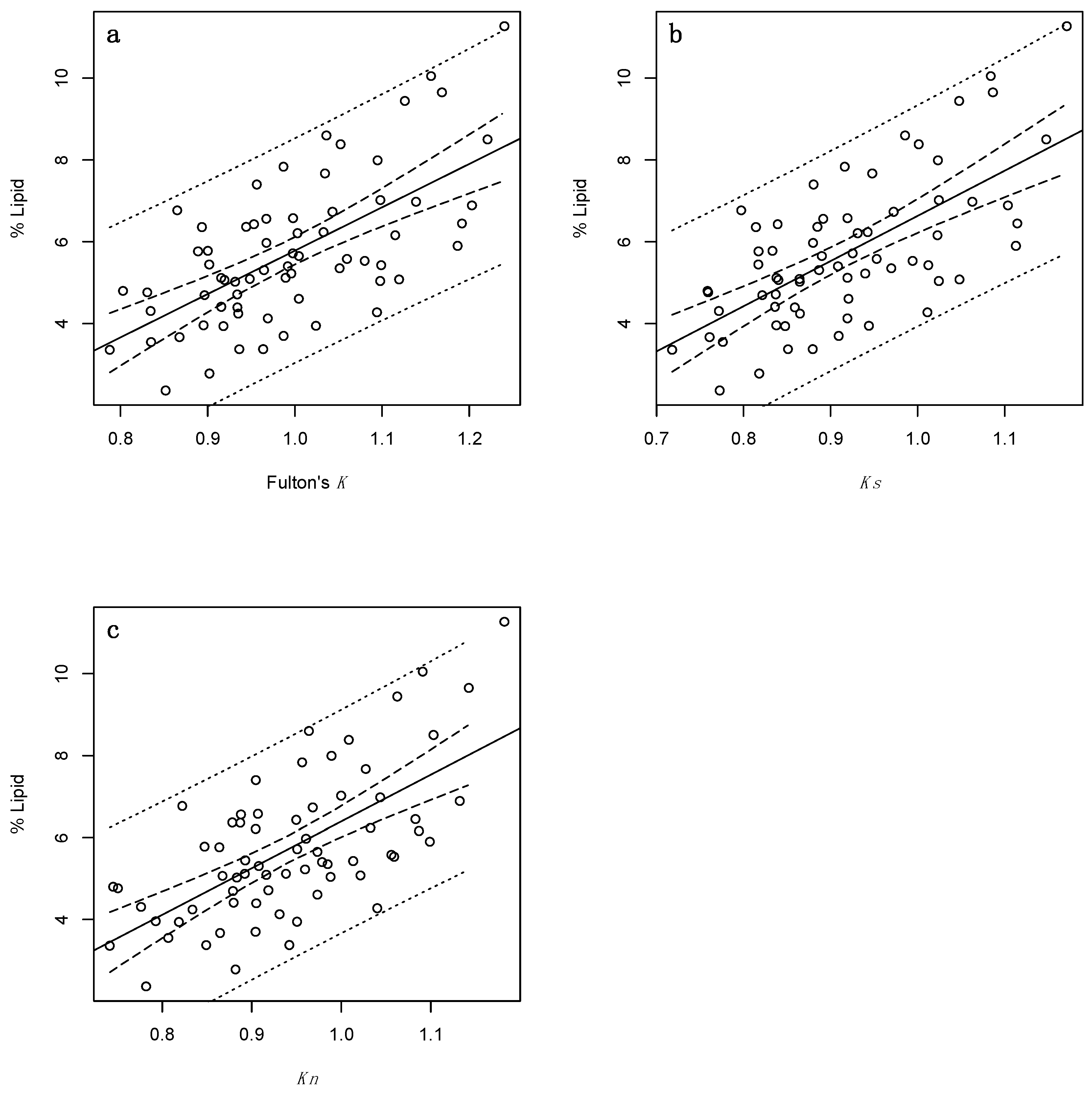
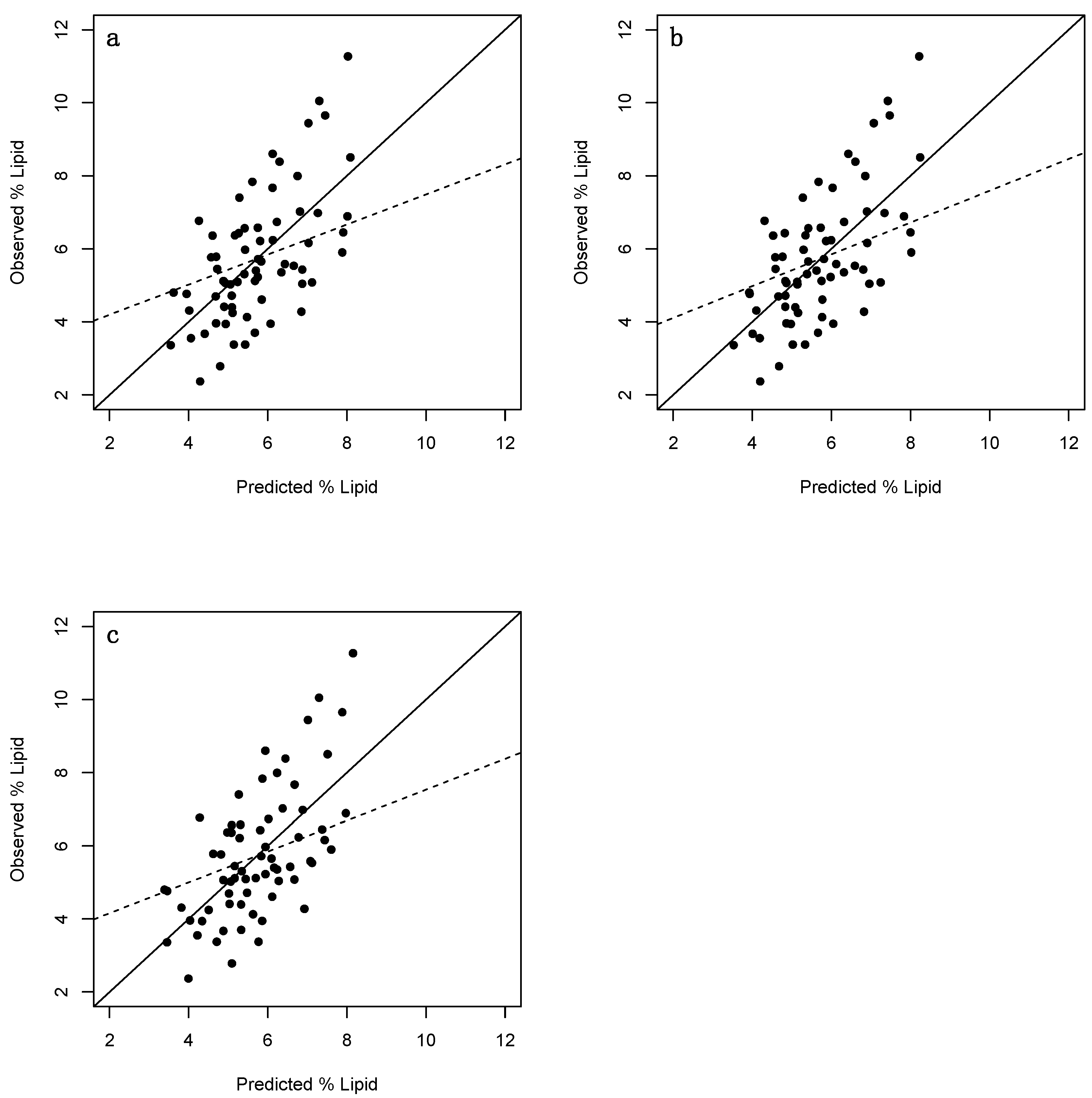
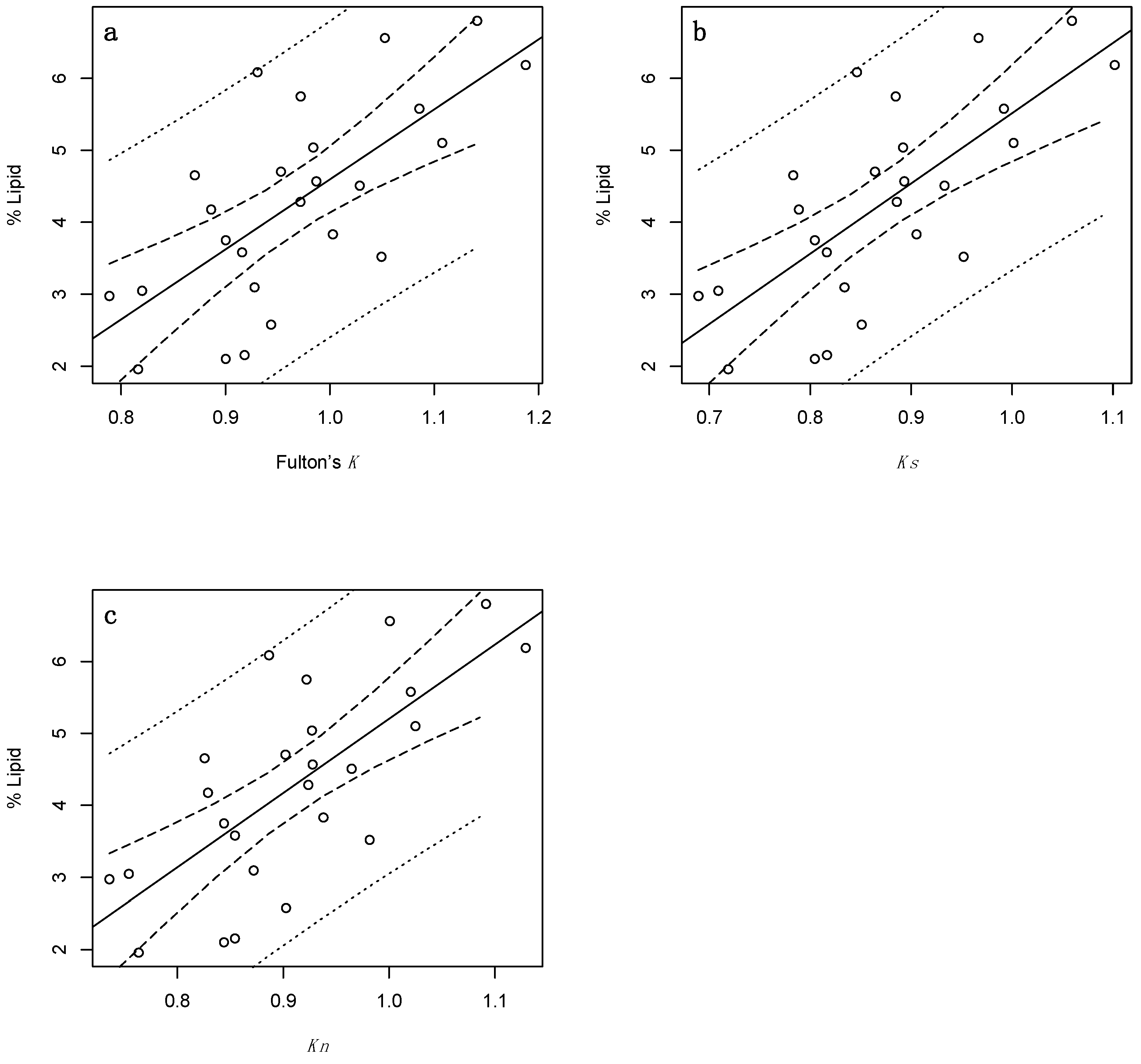
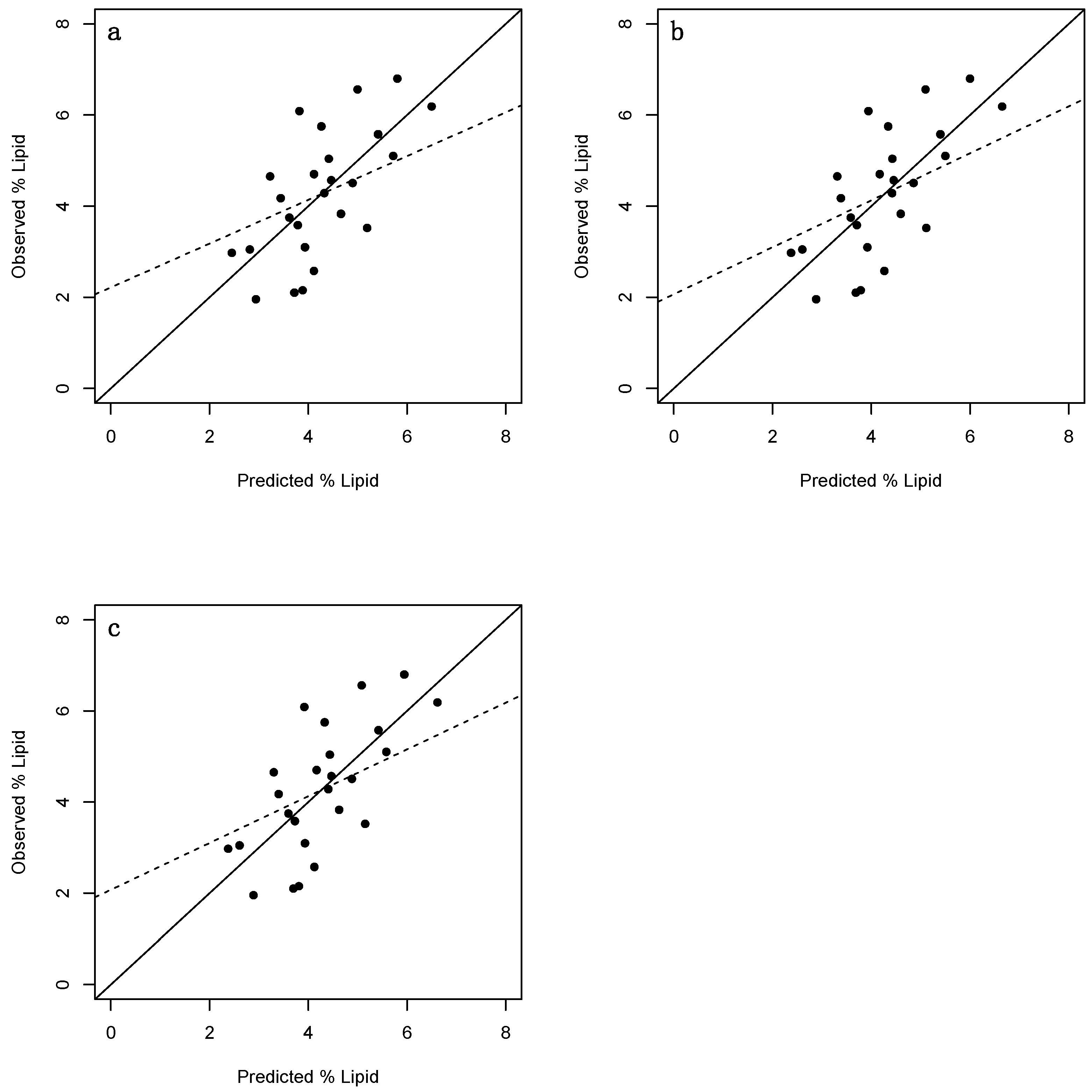
3. Results
3.1. Adults
3.2. Juveniles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scott, W.B.; Crossman, E.J. Freshwater Fishes of Canada. J. Fish. Res. Board Can. 1973, 184, 966. [Google Scholar]
- Gray, M.A.; Cunjak, R.A.; Munkittrick, K.R. Site fidelity of slimy sculpin (Cottus cognatus): Insights from stable carbon and nitrogen analysis. Can. J. Fish. Aquat. Sci. 2004, 61, 1717–1722. [Google Scholar] [CrossRef]
- Cunjak, R.A.; Roussel, J.M.; Gray, M.A.; Dietrich, J.P.; Cartwright, D.F.; Munkittrick, K.R.; Jardine, T.D. Using stable isotope analysis with telemetry or mark-recapture data to identify fish movement and foraging. Oecologia 2005, 144, 636–646. [Google Scholar] [CrossRef] [PubMed]
- Van Vliet, W.H. An ecological study of Cottus cognatus Richardson in northern Saskatchewan. Master’s Thesis, University of Saskatchewan, Saskatoon, SK, Canada, 1964. [Google Scholar]
- Keeler, R.A.; Cunjak, R.A. Reproductive ecology of slimy sculpin in small New Brunswick streams. Trans. Am. Fish. Soc. 2007, 136, 1762–1768. [Google Scholar] [CrossRef]
- Brasfield, S.M.; Tetreault, G.R.; McMaster, M.E.; Bennett, J.; Munkittrick, K.R. Seasonal patterns of gonad size, liver size, and in vitro gonadal steroidogenic capacity in slimy sculpin (Cottus cognatus). Water Qual. Res. J. Can. 2013, 48, 243–254. [Google Scholar] [CrossRef]
- Gray, M.A.; Curry, R.A.; Arciszewski, T.J.; Munkittrick, K.R.; Brasfield, S.M. The biology and ecology of slimy sculpin: A recipe for effective environmental monitoring. FACETS 2018, 3, 103–127. [Google Scholar] [CrossRef]
- Galloway, B.J.; Munkittrick, K.R.; Currie, S.; Gray, M.A.; Curry, R.A.; Wood, C.S. Examination of the responses of slimy sculpin (Cottus cognatus) and white sucker (Catostomus commersoni) collected on the Saint John River (Canada) downstream of pulp mill, paper mill, and sewage discharges. Environ. Toxicol. Chem. 2003, 22, 2898–2907. [Google Scholar] [CrossRef]
- Gray, M.A.; Munkittrick, K.R. An effects-based assessment of slimy sculpin (Cottus cognatus) populations in agricultural regions of northwestern New Brunswick. Water Qual. Res. J. Can. 2005, 40, 16–27. [Google Scholar] [CrossRef]
- Tetreault, G.R.; McMaster, M.E.; Dixon, D.G.; Parrott, J.L. Using reproductive endpoints in small forage fish species to evaluate the effects of Athabasca oil sands activities. Environ. Toxicol. Chem. 2003, 22, 2775–2782. [Google Scholar] [CrossRef]
- Bond, M.J.; Jones, N.E.; Haxton, T.J. Growth and life history patterns of a small-bodied stream fish, Cottus cognatus, in hydropeaking and natural rivers of northern Ontario. River Res. Appl. 2016, 32, 721–733. [Google Scholar] [CrossRef]
- Kelly, B.; Smokorowski, K.E.; Power, M. Slimy Sculpin (Cottus cognatus) annual growth in contrasting regulated and unregulated riverine environments. Hydrobiologia 2016, 768, 239–253. [Google Scholar] [CrossRef]
- Tesch, F.W. Age and growth. In Methods for Assessment of Fish Production in Fresh Waters; Ricker, W.E., Ed.; Blackwell Scientific Publications: Oxford, UK, 1968; pp. 93–123. [Google Scholar]
- Moyes, C.D.; West, T.G. Exercise metabolism of fish. In Biochemistry and Molecular Biology of Fishes; Hochachka, P.W., Mommsen, T.P., Eds.; Elsevier: Amsterdam, The Netherlands, 1995; pp. 367–392. [Google Scholar]
- Wootton, R.J. Ecology of Teleost Fishes, 2nd ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998. [Google Scholar]
- Hayes, J.P.; Shonkwiler, J.S. Morphometric indicators of body condition: Worthwhile or wishful thinking? In Body Composition Analysis of Animals: A Handbook of Non-Destructive Methods; Speakman, J.R., Ed.; Cambridge University Press: Cambridge, UK, 2001; pp. 8–38. [Google Scholar]
- Lloret, J.; Shulman, G.; Love, R.M. Condition and Health Indicators of Exploited Marine Fishes; John Wiley & Sons, Ltd.: Chichester, UK, 2013. [Google Scholar] [CrossRef]
- Adams, S.M. Ecological role of lipids in the health and success of fish populations. In Lipids in Freshwater Ecosystems; Arts, M.T., Wainman, B.C., Eds.; Springer: New York, NY, USA, 1999; pp. 132–160. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Phys. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Blackwell, B.G.; Brown, M.L.; Willis, D.W. Relative weight (Wr) status and current use in fisheries assessment and management. Rev. Fish. Sci. 2000, 8, 1–44. [Google Scholar] [CrossRef]
- Ricker, W.E. Computation and interpretation of biological statistics of fish populations. J. Fish. Res. Board Can. 1975, 191, 1–382. [Google Scholar]
- Le Cren, E.D. The length-weight relationship and seasonal cycle in gonad weight and condition in the perch (Perca fluviatilis). J. Anim. Ecol. 1951, 20, 201–219. [Google Scholar] [CrossRef]
- Herbinger, C.M.; Friars, G.W. Correlation between condition factor and total lipid content in Atlantic salmon, Salmo salar L., parr. Aquacult. Fish. Manag. 1991, 22, 527–529. [Google Scholar] [CrossRef]
- Stevenson, R.D.; Woods, W.A., Jr. Condition indices for conservation: New uses for evolving tools. Integr. Comp. Biol. 2006, 46, 1169–1190. [Google Scholar] [CrossRef]
- Peters, A.K.; Jones, M.L.; Honeyfield, D.C.; Bence, J.R. Monitoring energetic status of Lake Michigan Chinook salmon using water content as a predictor of whole-fish lipid content. J. Great Lakes Res. 2007, 33, 253–263. [Google Scholar] [CrossRef]
- Brosset, P.; Lloret, J.; Muñoz, M.; Fauvel, C.; Van Beveren, E.; Marques, V.; Fromentin, J.M.; Ménard, F.; Saraux, C. Body reserves mediate trade-offs between life-history traits: New insights from small pelagic fish reproduction. Roy. Soc. Open Sci. 2016, 3, 160–202. [Google Scholar] [CrossRef]
- Mion, M.; Thorsen, A.; Vitale, F.; Dierking, J.; Herrmann, J.P.; Huwer, B.; von Dewitz, B.; Casini, M. Effect of fish length and nutritional condition on the fecundity of distressed Atlantic cod Gadus morhua from the Baltic Sea. J. Fish Biol. 2018, 92, 1016–1034. [Google Scholar] [CrossRef]
- Brosset, P.; Fromentin, J.M.; Van Beveren, E.; Lloret, J.; Marques, V.; Basilone, G.; Bonanno, A.; Carpi, P.; De Felice, A.; Donato, F.; et al. Spatio-temporal patterns and environmental controls of small pelagic fish body condition from contrasted Mediterranean areas. Prog. Oceanogr. 2017, 151, 149–162. [Google Scholar] [CrossRef]
- Hoey, A.S.; McCormick, M.I. Selective predation for low body condition at the larval-juvenile transition of a coral reef fish. Oecologia 2004, 139, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Froese, R. Cube law, condition factor and weight–length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Fulton, T.W. The rate of growth of fishes. In Part III of the Twenty-Second Annual Report of the Fishery Board for Scotland; Fishery Board for Scotland: Glasgow, UK, 1904. [Google Scholar]
- Marshall, C.T.; Needle, C.L.; Yaragina, N.A.; Ajiad, A.M.; Gusev, E. Deriving condition indices from standard fisheries databases and evaluating their sensitivity to variation in stored energy reserves. Can. J. Fish. Aquat. Sci. 2004, 61, 1900–1917. [Google Scholar] [CrossRef]
- Davidson, D.; Marshall, C.T. Are morphometric indices accurate indicators of stored energy in herring Clupea harengus? J. Fish Biol. 2010, 76, 913–929. [Google Scholar] [CrossRef]
- Wentworth, C.K. A scale of grade and class terms for clastic sediments. J. Geol. 1922, 30, 377–392. [Google Scholar] [CrossRef]
- Elson, P.F. The pollett apron seine. J. Fish. Res. Board Can. 1962, 19, 93–100. [Google Scholar] [CrossRef]
- Beverton, R.J.H.; Holt, S.J. On the Dynamics of Exploited Fish Populations; Chapman and Hall: London, UK, 1993. [Google Scholar]
- Austin, P.C.; Hux, J.E. A brief note on overlapping confidence intervals. J. Vasc. Surg. 2002, 36, 194–195. [Google Scholar] [CrossRef]
- Hards, A.R.; Cunjak, R.A. The growth of slimy sculpin downstream of storage reservoirs (working title). manuscript in preparation.
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Burnham, K.P.; Anderson, D.R.; Huyvaert, K.P. AIC model selection and multimodel inference in behavioral ecology: Some background, observations, and comparisons. Behav. Ecol. Sociobiol. 2011, 65, 23–35. [Google Scholar] [CrossRef]
- Cook, R.D. Detection of influential observation in linear regression. Technometrics 1977, 19, 15–18. [Google Scholar] [CrossRef]
- Cook, R.D.; Weisberg, S. Residuals and Influence in Regression; Chapman and Hall: New York, NY, USA, 1982. [Google Scholar]
- Gershanovich, A.D.; Markevich, N.M.; Dergaleva, Z.T. Using the condition factor in ichthyological research. J. Ichthyol. 1984, 24, 78–90. [Google Scholar]
- Carlander, K.D. Handbook of Freshwater Fishery Biology; vol. 1; Iowa State University Press: Ames, IA, USA, 1969. [Google Scholar]
- Mozsár, A.; Boros, G.; Sály, P.; Antal, L.; Nagy, S.A. Relationship between Fulton’s condition factor and proximate body composition in three freshwater fish species. J. Appl. Ichthyol. 2015, 31, 315–320. [Google Scholar] [CrossRef]
- Schloesser, R.W.; Fabrizio, M.C. Condition indices as surrogates of energy density and lipid content in juveniles of three fish species. Trans. Am. Fish. Soc. 2017, 146, 1058–1069. [Google Scholar] [CrossRef]
- Brosset, P.; Fromentin, J.M.; Ménard, F.; Pernet, F.; Bourdeix, J.H.; Bigot, J.L.; Van Beveren, E.; Pérez Roda, M.A.; Choy, S.; Saraux, C. Measurement and analysis of small pelagic fish condition: A suitable method for rapid evaluation in the field. J. Exp. Mar. Biol. Ecol. 2015, 462, 90–97. [Google Scholar] [CrossRef]
- McPherson, L.R.; Slotte, A.; Kvamme, C.; Meier, S.; Marshall, C.T. Inconsistencies in measurement of fish condition: A comparison of four indices of fat reserves for Atlantic herring (Clupea harengus). ICES J. Mar. Sci. 2011, 68, 52–60. [Google Scholar] [CrossRef]
- Jonas, J.L.; Kraft, C.E.; Margenau, T.L. Assessment of seasonal changes in energy density and condition in age-0 and age-1 muskellunge. Trans. Am. Fish. Soc. 1996, 125, 203–210. [Google Scholar] [CrossRef]
- Chellappa, S.; Huntingford, F.A.; Strang, R.H.C.; Thomson, R.Y. Condition factor and hepatosomatic index as estimates of energy status in male three-spined stickleback. J. Fish Biol. 1995, 47, 775–787. [Google Scholar] [CrossRef]
- McCue, M.D. Starvation physiology: Reviewing the different strategies animals use to survive a common challenge. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2010, 156, 1–18. [Google Scholar] [CrossRef]
- Love, R.M. The Chemical Biology of Fishes; Academic Press: London, UK, 1970; Volume 1. [Google Scholar]
- Madenjian, C.P.; Elliott, R.F.; DeSorcie, T.J.; Stedman, R.M.; O’Connor, D.V.; Rottiers, D.V. Lipid concentrations in Lake Michigan fishes: Seasonal, spatial, ontogenetic, and long-term trends. J. Great Lakes Res. 2000, 26, 427–444. [Google Scholar] [CrossRef]
- Bennett, P.M.; Weber, L.P.; Janz, D.M. Comparison of chloroform-methanol-extracted and solvent-free triglyceride determinations in four fish species. J. Aquat. Anim. Health 2007, 19, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Sündermann, A.; Eggers, L.F.; Schwudke, D. Liquid Extraction: Bligh and Dyer. In Encyclopedia of Lipidomics; Wenk, M., Ed.; Springer: Dordrecht, The Netherlands, 2016. [Google Scholar]
- Dibble, K.L.; Yard, M.D.; Ward, D.L.; Yackulic, C.B. Does bioelectrical impedance analysis accurately estimate the physiological condition of threatened and endangered desert fish species? Trans. Am. Fish Soc. 2017, 146, 888–902. [Google Scholar] [CrossRef]
- Wuenschel, M.J.; McElroy, W.D.; Oliveira, K.; McBride, R.S. Measuring fish condition: An evaluation of new and old metrics for three species with contrasting life histories. Can. J. Fish. Aquat. Sci. 2018, 999, 1–18. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Type | Variable | Sex/Size | Mean ± S.D. | Range |
|---|---|---|---|---|
| Lethal | Total wet mass (g) | Adults | 4.0 ± 2.4 | 1.2–9.5 |
| Juveniles | 0.5 ± 0.2 | 0.2–1.0 | ||
| Total Length (mm) | Adults | 71.0 ± 13.6 | 51–102 | |
| Juveniles | 37.6 ± 4.2 | 29–45 | ||
| Fulton’s K | Adults | 1.0 ± 0.1 | 0.8–1.2 | |
| Juveniles | 1.0 ± 0.1 | 0.8–1.2 | ||
| Ks | Adults | 0.9 ± 0.1 | 0.7–1.2 | |
| Juveniles | 0.9 ± 0.1 | 0.7–1.1 | ||
| Kn | Adults | 0.9 ± 0.1 | 0.7–1.2 | |
| Juveniles | 0.9 ± 0.1 | 0.7–1.1 | ||
| Percent Lipid | Adults | 5.7 ± 1.8 | 2.4–11.3 | |
| Juveniles | 4.3 ± 1.4 | 2.0–6.8 | ||
| Non-Lethal | Total wet mass (g) | Adults | 3.6 ± 2.0 | 1.1–16.2 |
| Juveniles | 0.6 ± 0.2 | 0.2–1.3 | ||
| Total Length (mm) | Adults | 67.7 ± 10.8 | 50–112 | |
| Juveniles | 38.7 ± 4.8 | 26–49 |
| Lethal/Non-Lethal | Maturity Status/Sex | Mass-Length Relationship | n | C.I. for b | R2 | RMSE |
|---|---|---|---|---|---|---|
| Non-lethal | Adult | loge(M) = −11.97 + 3.12 × loge(L) | 937 | 3.08–3.16 | 0.96 | 0.10 |
| Juvenile | loge(M) = −11.05 + 2.89 × logeL) | 509 | 2.81–2.96 | 0.92 | 0.11 | |
| Combined | loge(M) = −11.51 + 3.01 × loge(L) | 1446 | 2.99–3.03 | 0.99 | 0.10 | |
| Lethal | Adult Females | loge(M) = −11.66 + 3.03 × loge(L) | 36 | 2.85–3.22 | 0.97 | 0.10 |
| Adult Males | loge(M) = −12.03 + 3.12 × loge(L) | 28 | 2.85–3.39 | 0.95 | 0.11 | |
| Combined | loge(M) = −11.83 + 3.07 × loge(L) | 68 1 | 2.94–3.21 | 0.97 | 0.10 |
| Candidate Model | R2 | RMSE | AICc | ΔAICc | wi | ER | LOOCV R2 | LOOCV RMSE |
|---|---|---|---|---|---|---|---|---|
| Ks | 0.44 | 1.34 | 236.84 | 0.00 | 0.59 | - | 0.39 | 1.37 |
| Kn | 0.42 | 1.35 | 238.43 | 1.59 | 0.27 | 2.22 | 0.38 | 1.38 |
| Fulton’s K | 0.41 | 1.37 | 239.65 | 2.81 | 0.14 | 4.07 | 0.37 | 1.39 |
| Intercept-only | - | 5.99 | 438.49 | 201.65 | 0.00 | 6.1243 | - | - |
| Candidate Model | R2 | RMSE | AICc | ΔAICc | wi | ER | LOOCV R2 | LOOCV RMSE |
|---|---|---|---|---|---|---|---|---|
| Kn | 0.52 | 1.00 | 76.13 | 0.00 | 0.42 | - | 0.45 | 1.03 |
| Ks | 0.51 | 1.01 | 76.24 | 0.11 | 0.40 | 1.06 | 0.45 | 1.03 |
| Fulton’s K | 0.48 | 1.04 | 77.88 | 1.75 | 0.18 | 2.27 | 0.41 | 1.06 |
| Intercept-only | - | 4.48 | 148.12 | 71.99 | 0.00 | 4.0615 | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hards, A.R.; Gray, M.A.; Noël, S.C.; Cunjak, R.A. Utility of Condition Indices as Predictors of Lipid Content in Slimy Sculpin (Cottus cognatus). Diversity 2019, 11, 71. https://doi.org/10.3390/d11050071
Hards AR, Gray MA, Noël SC, Cunjak RA. Utility of Condition Indices as Predictors of Lipid Content in Slimy Sculpin (Cottus cognatus). Diversity. 2019; 11(5):71. https://doi.org/10.3390/d11050071
Chicago/Turabian StyleHards, Adrian R., Michelle A. Gray, Sophia C. Noël, and Rick A. Cunjak. 2019. "Utility of Condition Indices as Predictors of Lipid Content in Slimy Sculpin (Cottus cognatus)" Diversity 11, no. 5: 71. https://doi.org/10.3390/d11050071
APA StyleHards, A. R., Gray, M. A., Noël, S. C., & Cunjak, R. A. (2019). Utility of Condition Indices as Predictors of Lipid Content in Slimy Sculpin (Cottus cognatus). Diversity, 11(5), 71. https://doi.org/10.3390/d11050071