Cross-Shelf Variation Among Juvenile and Adult Coral Assemblages on Australia’s Great Barrier Reef


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Design
2.2. Survey Methods for Juvenile Coral and Adult Coral Assemblages
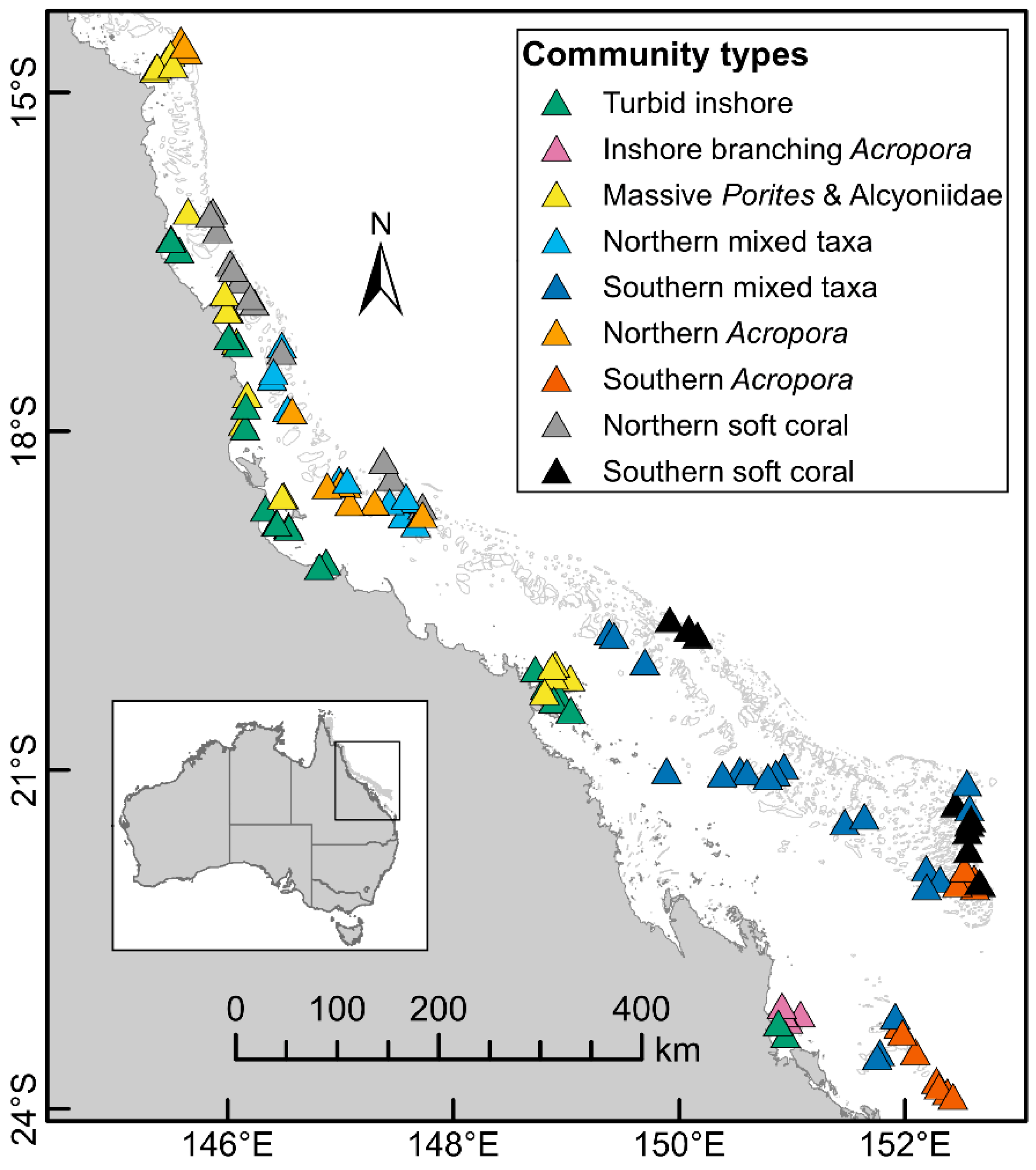
2.3. Community Types
2.4. Analysis of the Composition of Juvenile and Adult Coral Assemblages
2.5. Relationship between Juvenile and Adult Coral Assemblages
3. Results
3.1. Coral Community Types
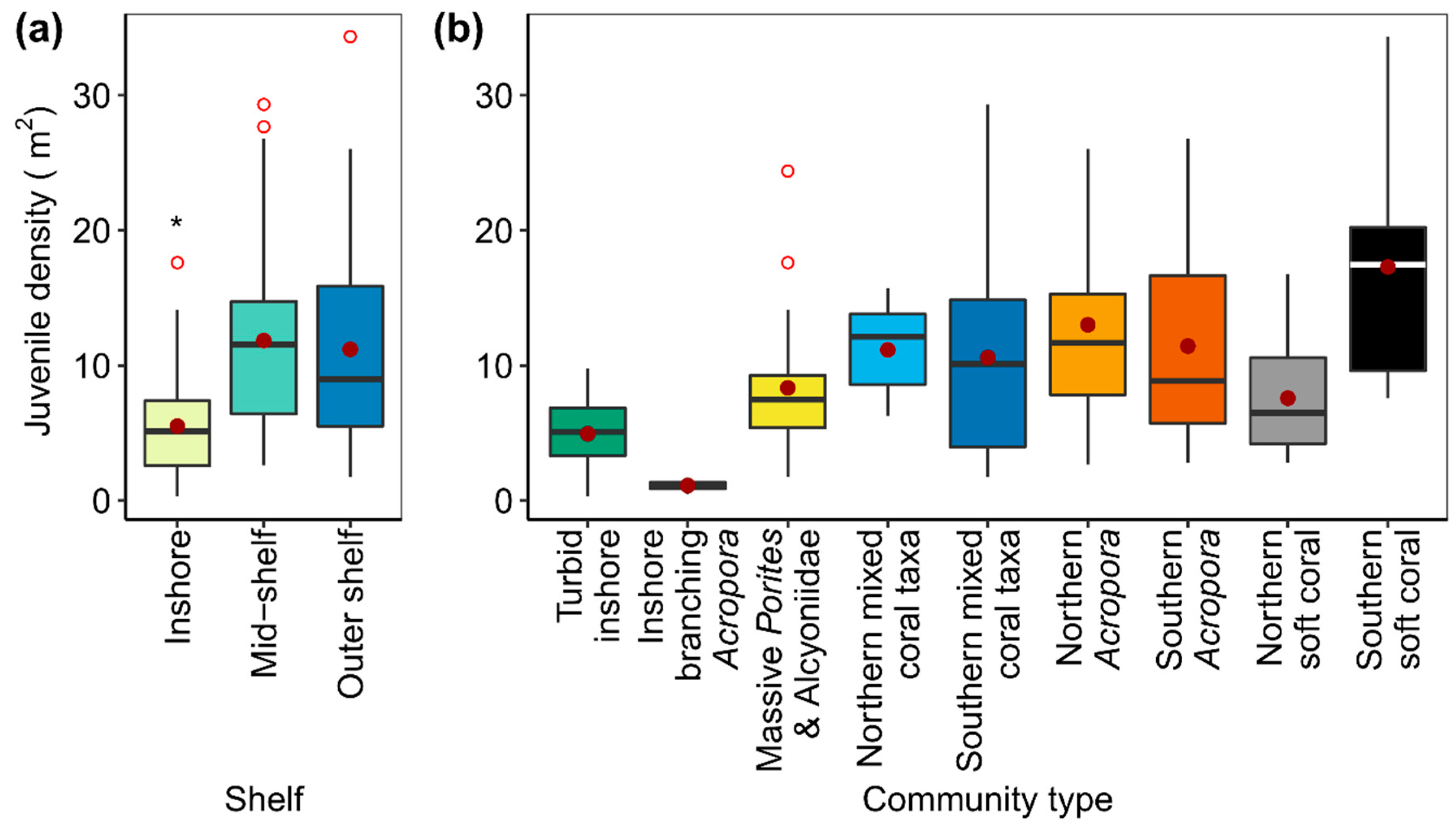
3.2. Juvenile Coral Densities
3.3. Composition of Juvenile Coral Assemblages on GBR
3.4. Relationship between Juvenile and Adult Coral Assemblages
3.4.1. Cross-Shelf
3.4.2. Community Types
3.4.3. Relative Abundance
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Inshore | Mid-Shelf | Outer Shelf | |||
|---|---|---|---|---|---|---|
| Juvenile | Adult | Juvenile | Adult | Juvenile | Adult | |
| Acropora | 14.82 ± 2.35 | 24.92 ± 4.43 | 21.84 ± 1.58 | 28.03 ± 1.89 | 29.26 ± 3.01 | 47.53 ± 5.12 |
| Porites | 10.12 ± 1.51 | 17.8 ± 3.31 | 24.74 ± 2.05 | 16.7 ± 1.49 | 30.29 ± 3.61 | 12.01 ± 1.98 |
| Montipora | 4.11 ± 0.71 | 7.81 ± 1.7 | 5.72 ± 1.02 | 8.51 ± 1.11 | 2.6 ± 0.5 | 4.71 ± 0.77 |
| Pocillopora | 2.39 ± 0.61 | 1.34 ± 0.43 | 6.68 ± 0.73 | 7.52 ± 1.1 | 8.05 ± 0.76 | 6.16 ± 0.88 |
| Goniopora | 5.18 ± 1.04 | 10.11 ± 2.33 | 0.42 ± 0.11 | 0.76 ± 0.16 | 0.23 ± 0.15 | 0.64 ± 0.21 |
| Echinopora | 0.74 ± 0.24 | 2.64 ± 0.98 | 1.28 ± 0.23 | 4.17 ± 0.68 | 0.5 ± 0.15 | 1.46 ± 0.23 |
| Isopora | 0.08 ± 0.05 | 0.18 ± 0.11 | 0.44 ± 0.14 | 1.88 ± 0.36 | 1.01 ± 0.24 | 6.93 ± 1.55 |
| Agariciidae | 1.74 ± 0.44 | 4.67 ± 0.85 | 0.89 ± 0.17 | 1.09 ± 0.16 | 0.5 ± 0.14 | 0.64 ± 0.18 |
| Stylophora | 0.26 ± 0.09 | 0.78 ± 0.24 | 2 ± 0.55 | 2.7 ± 0.35 | 4.79 ± 0.99 | 3.33 ± 0.51 |
| Favia | 7.59 ± 0.95 | 2.05 ± 0.42 | 4.93 ± 0.89 | 2.07 ± 0.22 | 2.81 ± 0.7 | 2.12 ± 0.28 |
| Pectiniidae | 3.35 ± 0.63 | 3.52 ± 0.8 | 0.97 ± 0.16 | 1.68 ± 0.26 | 0.1 ± 0.05 | 0.35 ± 0.11 |
| Goniastrea | 4.47 ± 1.1 | 2.11 ± 0.74 | 2.31 ± 0.3 | 1.64 ± 0.13 | 1.09 ± 0.24 | 1.62 ± 0.24 |
| Seriatopora | 0.69 ± 0.27 | 0.5 ± 0.19 | 3.64 ± 0.57 | 3.01 ± 0.53 | 4.27 ± 1.23 | 1.4 ± 0.51 |
| Galaxea | 4.37 ± 0.73 | 2.89 ± 0.86 | 1.23 ± 0.31 | 1.42 ± 0.31 | 0.77 ± 0.2 | 0.55 ± 0.12 |
| Fungiidae | 8.3 ± 1.76 | 1.88 ± 0.38 | 4.58 ± 0.53 | 2.26 ± 0.35 | 1.3 ± 0.41 | 0.27 ± 0.08 |
| Turbinaria | 8.57 ± 2.21 | 3.44 ± 0.98 | 0.22 ± 0.07 | 0.49 ± 0.19 | 0.15 ± 0.07 | 0.44 ± 0.19 |
| Lobophyllia | 4.77 ± 0.79 | 2.02 ± 0.34 | 3.37 ± 0.41 | 1.45 ± 0.23 | 1.46 ± 0.59 | 0.49 ± 0.19 |
| Platygyra | 1.37 ± 0.25 | 1.24 ± 0.22 | 1.2 ± 0.2 | 1.47 ± 0.17 | 0.87 ± 0.23 | 1.25 ± 0.25 |
| Other hard coral | 2.3 ± 0.72 | 0.15 ± 0.04 | 1.58 ± 0.27 | 2.39 ± 0.22 | 2.43 ± 0.56 | 1.12 ± 0.14 |
| Favites | 2.97 ± 0.48 | 1.05 ± 0.24 | 2.87 ± 0.34 | 1.51 ± 0.14 | 1.92 ± 0.54 | 1.18 ± 0.2 |
| Diploastrea | 0 ± 0 | 1.28 ± 0.42 | 0.02 ± 0.02 | 1.33 ± 0.48 | 0.01 ± 0.01 | 0.89 ± 0.37 |
| Merulina | 1.1 ± 0.22 | 1.3 ± 0.36 | 0.94 ± 0.25 | 1.31 ± 0.24 | 0.33 ± 0.16 | 0.54 ± 0.18 |
| Siderastreidae | 1.86 ± 0.55 | 1.58 ± 0.96 | 0.72 ± 0.2 | 1.08 ± 0.31 | 1.25 ± 0.36 | 0.52 ± 0.18 |
| Cyphastrea | 1.87 ± 0.5 | 1.37 ± 0.5 | 2.48 ± 0.32 | 0.81 ± 0.1 | 1.34 ± 0.27 | 0.38 ± 0.08 |
| Hydnophora | 0.43 ± 0.1 | 0.98 ± 0.33 | 0.65 ± 0.14 | 0.93 ± 0.13 | 0.16 ± 0.07 | 0.53 ± 0.1 |
| Other Faviidae | 1.48 ± 0.54 | 0.91 ± 0.4 | 0.53 ± 0.15 | 0.73 ± 0.12 | 0.29 ± 0.14 | 0.5 ± 0.09 |
| Symphyllia | 0.04 ± 0.03 | 0.11 ± 0.04 | 0.23 ± 0.07 | 0.97 ± 0.13 | 0.1 ± 0.04 | 0.63 ± 0.12 |
| Montastrea | 0.54 ± 0.16 | 0.09 ± 0.04 | 2.11 ± 0.29 | 0.82 ± 0.1 | 1.26 ± 0.31 | 0.72 ± 0.13 |
| Leptoria | 0.37 ± 0.12 | 0.34 ± 0.18 | 0.31 ± 0.11 | 0.64 ± 0.09 | 0.18 ± 0.09 | 0.47 ± 0.1 |
| Astreopora | 0.28 ± 0.14 | 0.42 ± 0.2 | 0.58 ± 0.15 | 0.37 ± 0.06 | 0.25 ± 0.14 | 0.47 ± 0.11 |
| Leptastrea | 0.81 ± 0.17 | 0.31 ± 0.11 | 0.19 ± 0.05 | 0.13 ± 0.03 | 0.16 ± 0.07 | 0.08 ± 0.06 |
| Euphyllidae | 0.54 ± 0.15 | 0.16 ± 0.06 | 0.21 ± 0.08 | 0.09 ± 0.03 | 0.05 ± 0.03 | 0.02 ± 0.02 |
| Acanthastrea | 0.52 ± 0.19 | 0.02 ± 0.01 | 0.13 ± 0.04 | 0.07 ± 0.02 | 0.23 ± 0.13 | 0.06 ± 0.02 |
| Moseleya | 1.96 ± 0.58 | 0.04 ± 0.02 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| Taxon | Turbid Inshore | Inshore Branching Acropora | Massive Porites & Alcyoniidae | Northern Mixed Coral Taxa | Southern Mixed Coral Taxa | Northern Acropora | Southern Acropora | Northern Soft Coral | Southern Soft Coral | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Juvenile | Adult | Juvenile | Adult | Juvenile | Adult | Juvenile | Adult | Juvenile | Adult | Juvenile | Adult | Juvenile | Adult | Juvenile | Adult | Juvenile | Adult | |
| Acropora | 9.17 ± 1.35 | 12.49 ± 3.85 | 49.97 ± 11.19 | 97.68 ± 0.84 | 14.34 ± 2.17 | 21.15 ± 2.95 | 14.79 ± 2.19 | 26.02 ± 4.23 | 22.3 ± 2.45 | 35.12 ± 3.6 | 26.38 ± 2.89 | 30.56 ± 4.76 | 44.02 ± 3.22 | 69.61 ± 8.97 | 25.3 ± 3.47 | 31.58 ± 4.58 | 20.08 ± 5.3 | 28.74 ± 4.05 |
| Porites | 10.15 ± 2.15 | 19.06 ± 5.87 | 1.46 ± 0.91 | 0.05 ± 0.05 | 12.19 ± 2.24 | 21.16 ± 3.15 | 39.04 ± 5.45 | 20.29 ± 2.73 | 22.17 ± 2.76 | 11.95 ± 1.87 | 26.32 ± 3.43 | 13.03 ± 2.91 | 14.92 ± 1.66 | 9.19 ± 3.8 | 34.08 ± 4.51 | 11.79 ± 1.74 | 34.79 ± 7.79 | 23.43 ± 3.16 |
| Montipora | 3.11 ± 0.73 | 3.77 ± 0.91 | 4.06 ± 2.5 | 0.66 ± 0.19 | 5.06 ± 1.16 | 12.29 ± 3 | 2.19 ± 0.54 | 4.96 ± 0.58 | 8.73 ± 2.03 | 13.63 ± 2.03 | 1.38 ± 0.43 | 5.71 ± 1.4 | 4.79 ± 1.14 | 3.92 ± 0.84 | 2.62 ± 0.63 | 4.29 ± 0.84 | 3.2 ± 0.86 | 4.49 ± 1.04 |
| Pocillopora | 0.76 ± 0.25 | 0.27 ± 0.1 | 3.33 ± 3.33 | 0.18 ± 0.18 | 4.42 ± 1 | 3.62 ± 1.05 | 5.47 ± 1.27 | 6.46 ± 1.18 | 7.3 ± 1.17 | 4.03 ± 0.51 | 7.79 ± 1.63 | 14.54 ± 3.82 | 9.08 ± 1.67 | 1.73 ± 0.58 | 6.94 ± 1.26 | 12.45 ± 1.41 | 7.37 ± 1.07 | 5.97 ± 1.03 |
| Goniopora | 7.81 ± 1.85 | 14.65 ± 3.86 | 1.56 ± 1.56 | 0 ± 0 | 2.37 ± 0.52 | 5.69 ± 2.3 | 0.22 ± 0.13 | 0.77 ± 0.25 | 0.76 ± 0.26 | 0.67 ± 0.23 | 0.48 ± 0.3 | 0.64 ± 0.41 | 0.13 ± 0.09 | 0.19 ± 0.1 | 0 ± 0 | 0.21 ± 0.07 | 0 ± 0 | 2 ± 0.56 |
| Echinopora | 0.95 ± 0.44 | 3.04 ± 1.86 | 0 ± 0 | 0 ± 0 | 1.13 ± 0.31 | 3.93 ± 1.01 | 0.68 ± 0.18 | 2.68 ± 0.56 | 1.22 ± 0.39 | 5.08 ± 1.18 | 0.6 ± 0.27 | 0.92 ± 0.23 | 0.13 ± 0.09 | 1.2 ± 0.82 | 1.23 ± 0.45 | 2.65 ± 0.38 | 0.9 ± 0.38 | 1.87 ± 0.35 |
| Isopora | 0.14 ± 0.11 | 0.35 ± 0.22 | 0 ± 0 | 0 ± 0 | 0.02 ± 0.02 | 0.02 ± 0.02 | 0.82 ± 0.44 | 1.35 ± 0.45 | 0.75 ± 0.28 | 3.85 ± 1.08 | 0.42 ± 0.21 | 2.14 ± 0.99 | 0.47 ± 0.25 | 0.91 ± 0.46 | 0.55 ± 0.32 | 9.7 ± 3.21 | 1.13 ± 0.47 | 5.95 ± 1.82 |
| Agariciidae | 2.66 ± 0.79 | 6.93 ± 1.43 | 0 ± 0 | 0 ± 0 | 1.11 ± 0.32 | 2.73 ± 0.61 | 0.89 ± 0.21 | 0.69 ± 0.23 | 0.48 ± 0.25 | 0.92 ± 0.19 | 1.12 ± 0.31 | 1.56 ± 0.65 | 0.25 ± 0.14 | 0.41 ± 0.14 | 0.59 ± 0.32 | 0.98 ± 0.41 | 1.16 ± 0.41 | 0.7 ± 0.15 |
| Stylophora | 0.24 ± 0.12 | 0.25 ± 0.12 | 0 ± 0 | 0 ± 0 | 2.24 ± 1.14 | 2.23 ± 0.52 | 3.08 ± 1.81 | 5.31 ± 1 | 1.11 ± 0.49 | 1.2 ± 0.3 | 2.98 ± 1.16 | 4.09 ± 0.66 | 1.56 ± 0.59 | 0.78 ± 0.28 | 2.94 ± 1.02 | 4.4 ± 0.8 | 6.48 ± 1.98 | 2.92 ± 0.41 |
| Favia | 6.88 ± 1.33 | 2.48 ± 0.74 | 3.25 ± 1.92 | 0.06 ± 0.06 | 8.83 ± 1.31 | 2.03 ± 0.36 | 2.66 ± 0.53 | 2.73 ± 0.45 | 7.8 ± 1.9 | 1.8 ± 0.33 | 1.84 ± 0.48 | 1.78 ± 0.52 | 1.48 ± 0.41 | 0.91 ± 0.2 | 3.65 ± 0.76 | 2.82 ± 0.37 | 2.53 ± 0.61 | 2.68 ± 0.48 |
| Pectiniidae | 4.12 ± 0.83 | 5.17 ± 1.46 | 0 ± 0 | 0.11 ± 0.11 | 2.8 ± 0.92 | 2.46 ± 0.53 | 1.28 ± 0.39 | 1.53 ± 0.36 | 0.53 ± 0.2 | 1.51 ± 0.32 | 0.63 ± 0.26 | 1.12 ± 0.56 | 0.34 ± 0.22 | 0.57 ± 0.4 | 0.85 ± 0.37 | 0.49 ± 0.13 | 0 ± 0 | 0.66 ± 0.27 |
| Goniastrea | 4.33 ± 1.66 | 3.06 ± 1.45 | 6.03 ± 5.48 | 0 ± 0 | 4.54 ± 1.21 | 1.57 ± 0.26 | 2.55 ± 0.42 | 1.76 ± 0.4 | 1.81 ± 0.38 | 1.48 ± 0.2 | 2.03 ± 0.63 | 1.55 ± 0.25 | 0.7 ± 0.33 | 0.63 ± 0.17 | 1.61 ± 0.43 | 2.1 ± 0.27 | 0.91 ± 0.31 | 2.22 ± 0.45 |
| Seriatopora | 0.28 ± 0.28 | 0.14 ± 0.11 | 0 ± 0 | 0 ± 0 | 1.26 ± 0.44 | 1.13 ± 0.35 | 1.95 ± 1.08 | 1.95 ± 0.85 | 3.66 ± 0.96 | 2.6 ± 0.81 | 5.46 ± 1.63 | 2.66 ± 0.52 | 3.57 ± 1 | 3.31 ± 1.6 | 2.65 ± 0.97 | 0.43 ± 0.18 | 7.97 ± 3.05 | 3.87 ± 1.37 |
| Galaxea | 5.04 ± 1.13 | 4.18 ± 1.65 | 0 ± 0 | 0 ± 0 | 3.99 ± 0.91 | 1.88 ± 0.37 | 1.93 ± 1.08 | 2.61 ± 1.15 | 0.64 ± 0.25 | 1.15 ± 0.33 | 1.57 ± 0.44 | 0.67 ± 0.27 | 0.54 ± 0.36 | 0.32 ± 0.11 | 1.15 ± 0.49 | 0.6 ± 0.14 | 0.73 ± 0.24 | 0.96 ± 0.27 |
| Fungiidae | 9.44 ± 2.67 | 2.5 ± 0.64 | 12.75 ± 8.88 | 0.17 ± 0.17 | 6.36 ± 1.7 | 2.11 ± 0.49 | 5.38 ± 1.53 | 1.42 ± 0.3 | 2.7 ± 0.52 | 2.45 ± 0.68 | 3.45 ± 0.77 | 1.07 ± 0.44 | 2.02 ± 1.26 | 0.88 ± 0.5 | 2 ± 0.63 | 0.31 ± 0.1 | 3.13 ± 1.24 | 0.71 ± 0.18 |
| Turbinaria | 11.94 ± 3.32 | 5.51 ± 1.72 | 3.22 ± 1.86 | 0 ± 0 | 4.74 ± 2.82 | 1.86 ± 0.81 | 0.22 ± 0.15 | 0.37 ± 0.24 | 0.12 ± 0.05 | 0.6 ± 0.24 | 0.04 ± 0.04 | 0.1 ± 0.06 | 0.08 ± 0.06 | 0.17 ± 0.08 | 0.15 ± 0.12 | 0.16 ± 0.05 | 0.38 ± 0.2 | 0.54 ± 0.26 |
| Lobophyllia | 4.77 ± 1.36 | 1.86 ± 0.43 | 2.79 ± 2.79 | 0.23 ± 0.23 | 4.88 ± 0.69 | 2.74 ± 0.52 | 3.76 ± 0.85 | 0.66 ± 0.15 | 4.07 ± 1 | 1.76 ± 0.42 | 2.82 ± 0.79 | 0.47 ± 0.2 | 1.74 ± 0.54 | 0.42 ± 0.2 | 0.92 ± 0.3 | 0.39 ± 0.14 | 0.78 ± 0.33 | 1.33 ± 0.51 |
| Platygyra | 0.9 ± 0.26 | 1.32 ± 0.39 | 0.93 ± 0.93 | 0.12 ± 0.07 | 1.93 ± 0.4 | 1.29 ± 0.2 | 1.33 ± 0.27 | 2.12 ± 0.4 | 1.36 ± 0.43 | 1.31 ± 0.2 | 0.33 ± 0.13 | 0.94 ± 0.35 | 1.01 ± 0.36 | 0.41 ± 0.15 | 1.36 ± 0.33 | 2.26 ± 0.52 | 0.49 ± 0.18 | 1.43 ± 0.34 |
| Other hard coral | 1.82 ± 0.56 | 0.04 ± 0.02 | 0 ± 0 | 0 ± 0 | 3.22 ± 1.34 | 0.66 ± 0.22 | 0.82 ± 0.19 | 2.22 ± 0.36 | 1.86 ± 0.52 | 1.66 ± 0.19 | 1.43 ± 0.4 | 4.14 ± 0.65 | 3.55 ± 1.31 | 1.13 ± 0.38 | 2.6 ± 0.66 | 1.54 ± 0.18 | 0.89 ± 0.32 | 0.98 ± 0.16 |
| Favites | 2.74 ± 0.65 | 1.17 ± 0.39 | 2.34 ± 2.34 | 0.06 ± 0.06 | 3.48 ± 0.7 | 1.15 ± 0.27 | 2.6 ± 0.51 | 1.89 ± 0.37 | 3.42 ± 0.68 | 1.2 ± 0.21 | 2.51 ± 0.55 | 1.52 ± 0.33 | 1.37 ± 0.51 | 0.61 ± 0.16 | 2.48 ± 0.95 | 1.88 ± 0.37 | 1.14 ± 0.51 | 1.39 ± 0.28 |
| Diploastrea | 0 ± 0 | 1.37 ± 0.76 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 1.84 ± 0.46 | 0.07 ± 0.07 | 2.83 ± 1.48 | 0 ± 0 | 0.14 ± 0.09 | 0 ± 0 | 2.15 ± 1.47 | 0 ± 0 | 0 ± 0 | 0.03 ± 0.03 | 1.69 ± 0.86 | 0 ± 0 | 0.34 ± 0.31 |
| Merulina | 1.52 ± 0.37 | 1.81 ± 0.65 | 0 ± 0 | 0 ± 0 | 1.03 ± 0.29 | 1.45 ± 0.44 | 0.2 ± 0.11 | 0.45 ± 0.15 | 1.08 ± 0.33 | 1.6 ± 0.35 | 0.07 ± 0.07 | 0.83 ± 0.33 | 0.26 ± 0.16 | 0.21 ± 0.12 | 1.21 ± 0.8 | 0.36 ± 0.11 | 0.5 ± 0.28 | 1.19 ± 0.47 |
| Siderastreidae | 1.84 ± 0.88 | 2.77 ± 1.91 | 3.32 ± 2.66 | 0.32 ± 0.22 | 1.36 ± 0.52 | 0.5 ± 0.1 | 0.78 ± 0.18 | 1.04 ± 0.28 | 0.2 ± 0.1 | 0.21 ± 0.05 | 2.8 ± 0.73 | 3.19 ± 1.33 | 1.55 ± 0.7 | 0.16 ± 0.07 | 0.19 ± 0.09 | 1.03 ± 0.41 | 1.17 ± 0.72 | 0.34 ± 0.1 |
| Cyphastrea | 1.78 ± 0.48 | 2.11 ± 0.96 | 4.67 ± 4.67 | 0.24 ± 0.24 | 1.56 ± 0.34 | 0.65 ± 0.18 | 2.97 ± 0.65 | 1.21 ± 0.26 | 1.59 ± 0.43 | 0.47 ± 0.11 | 3.54 ± 0.9 | 1.14 ± 0.22 | 1.54 ± 0.5 | 0.45 ± 0.19 | 0.88 ± 0.28 | 0.51 ± 0.15 | 2.22 ± 0.53 | 0.39 ± 0.09 |
| Hydnophora | 0.6 ± 0.17 | 1.16 ± 0.6 | 0 ± 0 | 0 ± 0 | 0.35 ± 0.1 | 1.1 ± 0.29 | 0.64 ± 0.2 | 1.53 ± 0.33 | 0.68 ± 0.25 | 0.65 ± 0.11 | 0.38 ± 0.19 | 0.44 ± 0.27 | 0.3 ± 0.18 | 0.2 ± 0.07 | 0.41 ± 0.25 | 0.69 ± 0.17 | 0.08 ± 0.05 | 0.96 ± 0.21 |
| Other Faviidae | 1.25 ± 0.49 | 1.42 ± 0.79 | 0 ± 0 | 0 ± 0 | 1.82 ± 0.97 | 0.5 ± 0.11 | 0.31 ± 0.11 | 1.22 ± 0.41 | 0.54 ± 0.21 | 0.5 ± 0.08 | 0.07 ± 0.07 | 0.84 ± 0.31 | 0.49 ± 0.34 | 0.35 ± 0.1 | 0.9 ± 0.47 | 0.5 ± 0.15 | 0.05 ± 0.05 | 0.67 ± 0.14 |
| Symphyllia | 0 ± 0 | 0.07 ± 0.05 | 0 ± 0 | 0 ± 0 | 0.1 ± 0.06 | 0.24 ± 0.06 | 0.3 ± 0.19 | 1.16 ± 0.31 | 0.08 ± 0.04 | 0.51 ± 0.12 | 0.22 ± 0.1 | 0.8 ± 0.19 | 0.2 ± 0.12 | 0.39 ± 0.21 | 0.37 ± 0.21 | 1.62 ± 0.35 | 0 ± 0 | 0.95 ± 0.16 |
| Montastrea | 0.29 ± 0.14 | 0.06 ± 0.05 | 0 ± 0 | 0 ± 0 | 0.88 ± 0.27 | 0.22 ± 0.09 | 1.28 ± 0.23 | 1.13 ± 0.23 | 1.86 ± 0.47 | 0.58 ± 0.14 | 1.87 ± 0.68 | 0.66 ± 0.15 | 3.02 ± 0.59 | 0.47 ± 0.23 | 1.42 ± 0.6 | 1 ± 0.21 | 1.67 ± 0.65 | 1.11 ± 0.26 |
| Leptoria | 0.13 ± 0.1 | 0.12 ± 0.07 | 0 ± 0 | 0 ± 0 | 1.05 ± 0.28 | 0.61 ± 0.36 | 0.08 ± 0.08 | 0.63 ± 0.14 | 0.09 ± 0.04 | 0.64 ± 0.17 | 0.11 ± 0.06 | 0.36 ± 0.18 | 0.24 ± 0.2 | 0.23 ± 0.08 | 0.27 ± 0.16 | 0.79 ± 0.15 | 0.14 ± 0.14 | 0.82 ± 0.21 |
| Astreopora | 0.23 ± 0.17 | 0.59 ± 0.39 | 0 ± 0 | 0 ± 0 | 0.43 ± 0.24 | 0.3 ± 0.09 | 0.63 ± 0.2 | 0.68 ± 0.2 | 0.74 ± 0.3 | 0.57 ± 0.12 | 0.45 ± 0.27 | 0.25 ± 0.11 | 0.11 ± 0.11 | 0.04 ± 0.03 | 0.37 ± 0.31 | 0.45 ± 0.14 | 0.06 ± 0.06 | 0.32 ± 0.11 |
| Leptastrea | 0.85 ± 0.21 | 0.15 ± 0.06 | 0.31 ± 0.31 | 0 ± 0 | 0.71 ± 0.27 | 0.54 ± 0.2 | 0.57 ± 0.17 | 0.06 ± 0.03 | 0.12 ± 0.06 | 0.06 ± 0.03 | 0.29 ± 0.16 | 0.11 ± 0.05 | 0.03 ± 0.03 | 0.14 ± 0.08 | 0.15 ± 0.1 | 0.23 ± 0.15 | 0.04 ± 0.04 | 0 ± 0 |
| Euphyllidae | 0.69 ± 0.19 | 0.07 ± 0.04 | 0 ± 0 | 0 ± 0 | 0.43 ± 0.22 | 0.27 ± 0.11 | 0.45 ± 0.21 | 0.23 ± 0.13 | 0.17 ± 0.13 | 0.06 ± 0.03 | 0.19 ± 0.1 | 0.02 ± 0.02 | 0 ± 0 | 0 ± 0 | 0.03 ± 0.03 | 0 ± 0 | 0 ± 0 | 0.02 ± 0.02 |
| Acanthastrea | 0.56 ± 0.32 | 0 ± 0 | 0 ± 0 | 0.11 ± 0.06 | 0.51 ± 0.23 | 0.05 ± 0.03 | 0.04 ± 0.04 | 0.05 ± 0.03 | 0.05 ± 0.04 | 0.09 ± 0.02 | 0.42 ± 0.17 | 0 ± 0 | 0.53 ± 0.34 | 0.05 ± 0.02 | 0.11 ± 0.11 | 0.09 ± 0.04 | 0 ± 0 | 0.05 ± 0.03 |
| Moseleya | 3.03 ± 0.93 | 0.06 ± 0.03 | 0 ± 0 | 0 ± 0 | 0.88 ± 0.63 | 0.02 ± 0.02 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
References
- Gattuso, J.-P.; Allemand, D.; Frankignoulle, M. Photosynthesis and calcification at cellular, organismal and community levels in coral reefs: A review of interactions and control by carbonate chemistry. Am. Zool. 1999, 39, 160–183. [Google Scholar] [CrossRef]
- Stella, J.S.; Pratchett, M.S.; Hutching, P.A. Jones GP Coral-associated invertebrates: Diversity, ecological importance and vulnerability to disturbance. Oceanogr. Mar. Biol. Ann. Rev. 2011, 49, 43–104. [Google Scholar]
- Coker, D.J.; Wilson, S.K.; Pratchett, M.S. Importance of live coral habitat for reef fishes. Rev. Fish. Biol. Fish. 2014, 24, 89–126. [Google Scholar] [CrossRef]
- Emslie, M.J.; Cheal, A.J.; Johns, K.A. Retention of habitat complexity minimizes disassembly of reef fish communities following disturbance: A large- scale natural experiment. PLoS ONE 2014, 9, e105384. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Baird, A.H.; Dinsdale, E.A.; Moltschaniwskyj, N.A.; Pratchett, M.S.; Tanner, J.E.; Willis, B.L. Assembly rules of reef corals are flexible along a steep climatic gradient. Curr. Biol. 2012, 22, 736–741. [Google Scholar] [CrossRef]
- Dornelas, M.; Connolly, S.R.; Hughes, T.P. Coral reef diversity refutes the neutral theory of biodiversity. Nature 2006, 440, 80–82. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Jackson, J.B.C. Do corals lie about their age? Some demographic consequences of partial mortality, fission and fusion. Science 1980, 209, 713–715. [Google Scholar] [CrossRef]
- Smith, L.D.; Hughes, T.P. An experimental assessment of survival, re-attachment and fecundity of coral fragmentation. J. Exp. Mar. Biol. Ecol. 1999, 235, 147–164. [Google Scholar] [CrossRef]
- Baird, A.H.; Guest, J.R.; Willis, B.L. Systematic and biogeological patterns in the reproductive biology of scleractinian corals. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 551–571. [Google Scholar] [CrossRef]
- Smith, L.D.; Devlin, M.; Haynes, D.; Gilmour, J.P. A Demographic approach to monitoring the health of coral reefs. Mar. Pollut. Bull. 2005, 51, 399–407. [Google Scholar] [CrossRef]
- Gilmour, J.P.; Smith, L.D.; Brinkman, R.M. Biannual spawning, rapid larval development and evidence of self-seeding for scleractinian corals at an isolated system of reefs. Mar. Biol. 2009, 156, 1297–1309. [Google Scholar] [CrossRef]
- Pratchett, M.S.; Anderson, K.D.; Hoogenboom, M.O.; Widman, E.; Baird, A.H.; Pandolfi, J.M.; Edmunds, P.J.; Lough, J.M. Spatial, temporal and taxonomic variation in coral growth: implications for the structure and function of coral reef ecosystems. Oceanogr. Mar. Biol. Ann. Rev. 2015, 53, 215–295. [Google Scholar]
- Roth, M.S.; Knowlton, N. Distribution, abundance, and microhabitat characterization of small juvenile coral at Palmyra Atoll. Mar. Ecol. Prog. Ser. 2009, 376, 133–142. [Google Scholar] [CrossRef]
- Graham, E.M.; Baird, A.H.; Connolly, S.R. Survival dynamics of scleractinian coral larvae and implications for dispersal. Coral Reefs 2008, 27, 529–539. [Google Scholar] [CrossRef]
- Graham, E.M.; Baird, A.H.; Willis, B.L.; Connolly, S.R. Effects of delayed settlement on post-settlement growth and survival of scleractinian coral larvae. Oecologia 2013, 173, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Penin, L.; Michonneau, F.; Baird, A.H.; Connolly, S.R.; Pratchett, M.S.; Kayal, M.; Adjeroud, M. Early post-settlement mortality and the structure of coral assemblages. Mar. Ecol. Prog. Ser. 2010, 408, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Martinez, S.; Abelson, A. Coral recruitment: the critical role of early post-settlement survival. ICES J. Mar. Sci. 2013, 70, 1294–1298. [Google Scholar] [CrossRef] [Green Version]
- Trapon, M.L.; Pratchett, M.S.; Hoey, A.S.; Baird, A.H. Influence of fish grazing and sedimentation on the early post-settlement survival of the tabular coral Acropora cytherea. Coral Reefs 2013, 32, 1051–1059. [Google Scholar] [CrossRef]
- Trapon, M.L.; Pratchett, M.S.; Hoey, A.S. Spatial variation in abundance, size and orientation of juvenile corals related to the biomass of parrotfishes on the Great Barrier Reef, Australia. PLoS ONE 2013, 8, e57788. [Google Scholar] [CrossRef]
- Baird, A.H.; Babcock, R.C.; Mundy, C.P. Habitat selection by larvae influences the depth distribution of six common coral species. Mar. Ecol. Prog. Ser. 2003, 252, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Done, T.J. Coral community adaptability to environmental change at the scales of regions, reefs and reef zones. Integr. Comp. Biol. 1999, 39, 66–79. [Google Scholar] [CrossRef]
- Glassom, D.; Chadwick, N.E. Recruitment, growth and mortality of juvenile corals at Eilat, northern Red Sea. Mar. Ecol. Prog. Ser. 2006, 318, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Penin, L.; Adjeroud, M.; Pratchett, M.S.; Hughes, T.P. Spatial distribution of juvenile and adult corals around Moorea (French Polynesia): Implications for population regulation. Bull. Mar. Sci. 2007, 80, 379–390. [Google Scholar]
- Hughes, T.P.; Baird, A.H.; Dinsdale, E.A.; Moltschaniwskyj, N.A.; Pratchett, M.S.; Tanner, J.E.; Willis, B.L. Supply-side ecology works both ways: the link between benthic adults, fecundity, and larval recruits. Ecology 2000, 81, 2241–2249. [Google Scholar] [CrossRef]
- Cetina-Heredia, P.; Connolly, S.R. A simple approximation for larval retention around reefs. Coral Reefs 2011, 30, 593–605. [Google Scholar] [CrossRef]
- Andutta, F.P.; Kingsford, M.J.; Wolanski, E. ‘Sticky water’ enables the retention of larvae in a reef mosaic. Estuar. Coast Shelf Sci. 2012, 101, 54–63. [Google Scholar] [CrossRef]
- Underwood, J.N.; Smith, L.D.; van Oppen, M.J.H.; Gilmour, J.P. Multiple scales of genetic connectivity in a brooding coral on isolated reefs following catastrophic bleaching. Mol. Ecol. 2007, 16, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Osborne, K.; Dolman, A.M.; Burgess, S.C.; Johns, K.A. Disturbance and the dynamics of coral cover on the Great Barrier Reef (1995–2009). PLoS ONE 2011, 6, e17516. [Google Scholar] [CrossRef]
- Osborne, K.; Thompson, A.A.; Cheal, A.J.; Johns, K.A.; Jonker, M.J.; Logan, M.; Miller, I.R.; Sweatman, H.P.A. Delayed coral recovery in a warming ocean. Glob. Chang. Biol. 2017, 23, 3869–3881. [Google Scholar] [CrossRef]
- Johns, K.A.; Osborne, K.O.; Logan, M. Contrasting rates of coral recovery and reassembly in coral communities on the Great Barrier Reef. Coral Reefs 2014, 33, 553–563. [Google Scholar] [CrossRef] [Green Version]
- Halford, A.; Cheal, A.J.; Ryan, D.; Williams, D. McB. Resilience to large-scale disturbance in coral and fish assemblages on the Great Barrier Reef. Ecology 2004, 85, 1892–1905. [Google Scholar] [CrossRef]
- Adjeroud, M.; Fernandez, J.M.; Carroll, A.G.; Harrison, P.L.; Penin, L. Spatial patterns and recruitment processes of coral assemblages among contrasting environmental conditions in the southwestern lagoon of New Caledonia. Mar. Pollut. Bull. 2010, 61, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Adjeroud, M.; Briand, M.J.; Kayal, M.; Dumas, P. Coral assemblages in Tonga: spatial patterns, replenishment capacities, and implications for conservation strategies. Environ. Monit. Assess 2013, 185, 5763–5773. [Google Scholar] [CrossRef]
- Edmunds, P.J.; Steneck, R.; Albright, R.; Carpenter, R.C.; Chui, A.P.Y.; Fan, T.-Y.; Harii, S.; Kitano, H.; Kurihara, H.; Legendre, L.; et al. Geographic variation in long-term trajectories of change in coral recruitment: A global-to-local perspective. Mar. Freshw. Res. 2015, 66, 609–622. [Google Scholar] [CrossRef]
- Edmunds, P.J. The hidden dynamics of low coral cover communities. Hydrobiologia 2018, 818, 193–209. [Google Scholar] [CrossRef]
- Doropoulos, C.; Ward, S.; Roff, G.; González-Rivero, M.; Mumby, P.J. Linking demographic processes of juvenile corals to benthic recovery trajectories in two common reef habitats. PLoS ONE 2015, 10, e0128535. [Google Scholar] [CrossRef]
- Thompson, A.; Schroeder, T.; Brando, V.E.; Schaffelke, B. Coral community responses to declining water quality; Whitsunday Islands, Great Barrier Reef, Australia. Coral Reefs 2014, 33, 923–938. [Google Scholar] [CrossRef]
- Bartlett, L.A.; Brinkhuis, V.I.P.; Ruzicka, R.R.; Colella, M.A.; Semon Lunz, K.; Leone, E.H.; Hallock, P. Dynamics of stony coral and octocoral juvenile assemblages following disturbance on patch reefs of the Florida Reef Tract. In Corals in a Changing World; Duque, C., Tello Camacho, E., Eds.; IntechOpen Limited: London, UK, 2018; ISBN 978-953-51-3910-2. [Google Scholar] [CrossRef]
- Álvarez-Noriega, M.; Baird, A.H.; Bridge, T.C.L.; Dornelas, M.; Fontoura, L.; Pizarro, O.; Precoda, K.; Torres-Pulliza, D.; Woods, R.M.; Zawada, K.; et al. Contrasting patterns of changes in abundance following a bleaching event between juvenile and adult scleractinian corals. Coral Reefs 2018, 37, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Bak, R.P.M.; Engel, M.S. Distribution, abundance and survival of juvenile hermatypic corals (Scleractinia) and the importance of life history strategies in the parental coral community. Mar. Biol. 1979, 54, 341–352. [Google Scholar] [CrossRef]
- Darling, E.S.; McClanahan, T.R.; Côté, I.M. Life histories predict coral community disassembly under multiple stressors. Glob. Chang. Biol. 2013, 19, 1930–1940. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, J.P.; Smith, L.D.; Heyward, A.J.; Baird, A.H.; Pratchett, M.S. Recovery of an isolated coral reef system following severe disturbance. Science 2013, 340, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Morri, C.; Montefalcone, M.; Lasagna, R.; Gatti, G.; Rovere, A.; Parravicini, V.; Baldelli, G.; Colantoni, P.; Bianchi, C.N. Through bleaching and tsunami: Coral reef recovery in the Maldives. Mar. Pollut. Bull. 2015, 98, 188–200. [Google Scholar] [CrossRef]
- Chong-Seng, K.M.; Graham, N.A.J.; Pratchett, M.S. Bottlenecks to coral recovery in the Seychelles. Coral Reefs 2014, 33, 449–461. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Chase, T.J.; Dietzel, A.; Hill, T.; Hoey, A.H.; Hoogenboom, M.O.; Jacobson, M.; et al. Global warming impairs stock-recruitment dynamics of corals. Nature 2019. [Google Scholar] [CrossRef]
- Pratchett, M.S.; Baird, A.H.; Bauman, A.G.; Burt, J.A. Abundance and composition of juvenile corals reveals divergent trajectories for coral assemblages across the United Arab Emirates. Mar. Pollut. Bull. 2017, 114, 1031–1035. [Google Scholar] [CrossRef]
- McClanahan, T.R. Decadal coral community reassembly on an African fringing reef. Coral Reefs 2014, 33, 939–950. [Google Scholar] [CrossRef]
- Adjeroud, M.; Kayal, M.; Iborra-Cantonnet, C.; Vercelloni, J.; Bosserelle, P.; Liao, V.; Chancerelle, Y.; Claudet, J.; Penin, L. Recovery of coral assemblages, despite acute and recurrent disturbances on a South Central Pacific reef. Sci. Rep. 2018, 8, 9680. [Google Scholar] [CrossRef] [PubMed]
- Graham, N.A.J.; Jennings, S.; MacNeil, M.A.; Mouillot, D.; Wilson, S.K. Predicting climate-driven regime shifts versus rebound potential in coral reefs. Nature 2015, 518, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Ninio, R.; Meekan, M. Spatial patterns in benthic communities and the dynamics of a mosaic ecosystem on the Great Barrier Reef, Australia. Coral Reefs. 2002, 21, 95–104. [Google Scholar] [CrossRef]
- Van Woesik, R.; Tomascik, T.; Blake, S. Coral assemblages and physico-chemical characteristics of the Whitsunday Islands: evidence of recent community changes. Mar. Freshw. Res. 1999, 50, 427–440. [Google Scholar] [CrossRef]
- Done, T. Patterns in the distribution of coral communities across the central Great Barrier Reef. Coral Reefs 1982, 1, 95–107. [Google Scholar] [CrossRef]
- Fabricius, K.E. Effects of terrestrial runoff on the ecology of corals and coral reefs: Review and synthesis. Mar. Pollut. Bull. 2005, 50, 125–166. [Google Scholar] [CrossRef]
- Fabricius, K.E.; Logan, M.; Weeks, S.; Brodie, J. The effects of river run-off on water clarity across the central Great Barrier Reef. Mar. Pollut. Bull. 2014, 84, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Anthony, K.R.N.; Connolly, S.R. Environmental limits to growth: physiological nice boundaries of corals along turbidity-light gradients. Oecologia 2004, 141, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Wolanski, E. Physical Oceanographic Processes of the Great Barrier Reef; CRC Press: Boca Raton, FL, USA, 1994. [Google Scholar]
- Choukroun, S.; Ridd, P.V.; Brinkman, R.; McKinna, L.I.W. On the surface circulation in the western Coral Sea and residence times in the Great Barrier Reef. J. Geophys. Res. 2010, 115, C06013. [Google Scholar] [CrossRef]
- Steinberg, C. Impacts of climate change on the physical oceanography of the Great Barrier Reef. In Climate Change and the Great Barrier Reef; Johnson, J.E., Marshall, P.A., Eds.; Great Barrier Reef Marine Park Authority and Australian Greenhouse Office: Townsville, Australia, 2007; pp. 51–74. [Google Scholar]
- Hopley, D. The Geomorphology of the Great Barrier Reef: Quaternary Development of Coral Reefs; Wiley: New York, NY, USA, 1982. [Google Scholar]
- Burgess, S.C.; Osborne, K.; Caley, M.J. Similar regional effects among local habitats on the structure of tropical reef fish and coral communities. Glob. Ecol. Biogeogr. 2010, 19, 363–375. [Google Scholar] [CrossRef]
- Emslie, M.J.; Pratchett, M.S.; Cheal, A.J.; Osborne, K. Great Barrier Reef butterflyfish community structure: the role of shelf position and benthic community type. Coral Reefs 2010, 29, 705–715. [Google Scholar] [CrossRef]
- Ninio, R.; Meekan, M.; Done, T.; Sweatman, H. Temporal patterns in coral assemblages on the Great Barrier Reef from local to large spatial scales. Mar. Ecol. Prog. Ser. 2000, 194, 65–74. [Google Scholar] [CrossRef]
- Trapon, M.L.; Pratchett, M.S.; Adjeroud, M.; Hoey, A.S.; Baird, A.H. Post-settlement growth and mortality rates of juvenile scleractinian coral in Moorea, French Polynesia versus Trunk Reef, Australia. Mar. Ecol. Prog. Ser. 2013, 488, 157–170. [Google Scholar] [CrossRef]
- Graham, N.A.J.; Chong-Seng, K.M.; Huchery, C.; Januchowski-Hartley, F.A.; Nash, K.L. Coral reef community composition in the context of disturbance history on the Great Barrier Reef, Australia. PLoS ONE 2014, 9, e101204. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; Knowlton, N.; Eakin, C.M.; Iglesias-Prieto, R.; et al. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef]
- Jones, R.; Ricardo, G.F.; Negri, A.P. Effects of sediments on the reproductive cycle of corals. Mar. Pollut. Bull. 2015, 100, 13–33. [Google Scholar] [CrossRef]
- Humanes, A.; Noonan, S.H.C.; Willis, B.L.; Fabricius, K.E.; Negri, A.P. Cumulative effects of nutrient enrichment and elevated temperature compromise the early life history stages of the coral Acropora tenuis. PLoS ONE 2016, 11, e0161616. [Google Scholar] [CrossRef] [PubMed]
- Humanes, A.; Fink, A.; Willis, B.L.; Fabricius, K.E.; de Beer, D.; Negri, A.P. Effects of suspended sediments and nutrient on juvenile corals. Mar. Pollut. Bull. 2017, 125, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Lam, V.Y.Y.; Chaloupka, M.; Thompson, A.; Doropoulos, C.; Mumby, P.J. Acute drivers influence recent inshore Great Barrier Reef dynamics. Proc. R. Soc. Lond. B Biol. Sci. 2018, 285, 1890. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Dietzel, A.; Eakin, C.M.; Heron, S.F.; Hoey, A.S.; Hoogenboom, M.O.; Liu, G.; et al. Global warming transforms coral reef assemblages. Nature 2018, 556, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Cheal, A.J.; MacNeil, M.A.; Emslie, M.J. Sweatman, H. The threat to coral reefs from more intense cyclones under climate change. Glob. Chang. Biol. 2017, 23, 1511–1524. [Google Scholar] [CrossRef]
- Uthicke, S.; Logan, M.; Liddy, M.; Francis, D.; Hardy, N.; Lamare, M. Climate change as an unexpected co-factor promoting coral eating seastar (Acanthaster planci) outbreaks. Sci. Rep. 2015, 5, 8402. [Google Scholar] [CrossRef]
- Jonker, M.; Johns, K.; Osborne, K. Surveys of Benthic Reef Communities using Underwater Digital Photography and Counts of Juvenile Corals; Long-term Monitoring of the Great Barrier Reef. Standard Operational Procedure Number 10; Australian Institute of Marine Science: Townsville, Australia, 2008.
- Veron, J.E.N. Corals of the world, 1st ed.; Australian Institute of Marine Science: Townsville, Australia, 2000; Volume 1–3.
- Fabricius, K.E.; Alderslade, P. Soft Corals and Sea Fans: A Comprehensive Guide to the Tropical Shallow Water Genera of the Central-West Pacific, the Indian Ocean and the Red Sea, 1st ed.; Australian Institute of Marine Science: Townsville, Australia, 2001; ISBN 064-232-210-4.
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Oxford, UK, 2012. [Google Scholar]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 30 May 2019).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R package version 2.5–2. 2018. Available online: https://CRAN.R-project.org/package=vegan (accessed on 30 May 2019).
- Wickham, H. Tidyverse: Easily Install and Load the ‘Tidyverse’. R package version 1.2.1. 2017. Available online: https://CRAN.R-project.org/package=tidyverse (accessed on 30 May 2019).
- Galili, T. Dendextend: An R package for visualizing, adjusting, and comparing trees of hierarchical clustering. Bioinformatics 2015, 31, 3718–3720. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Rigby, R.A.; Stasinopoulos, D.M. Generalized additive models for location, scale and shape (with discussion). Appl. Stat. 2005, 54, 507–544. [Google Scholar] [CrossRef]
- Shlesinger, T.; Loya, Y. Recruitment, mortality, and resilience potential of scleractinian corals at Eilat, Red Sea. Coral Reefs 2016, 35, 1357–1368. [Google Scholar] [CrossRef]
- Connell, J.H.; Hughes, T.P.; Wallace, C.C. A 30-year study of coral abundance, recruitment, and disturbance as several scales in space and time. Ecol. Monogr. 1997, 67, 461–488. [Google Scholar] [CrossRef]
- Caley, M.J.; Carr, M.H.; Hixon, M.A.; Hughes, T.P.; Jones, G.P.; Menge, B.A. Recruitment and the local dynamics of open marine populations. Annu. Rev. Ecol. Syst. 1996, 27, 477–500. [Google Scholar] [CrossRef]
- González-Rivero, M.; Beijom, O.; Rodriguez-Ramirez, A.; Holtrop, T.; González-Marrero, Y.; Ganase, A.; Roelfsema, C.; Phinn, S.; Hoegh-Guldberg, O. Scaling up ecological measurements of coral reefs using semi-automated field image collection and analysis. Remote Sens. 2016, 8, 30. [Google Scholar] [CrossRef]
- Anthony, K.R.N. Enhanced energy status of corals on coastal, high-turbidity reefs. Mar. Ecol. Prog. Ser. 2006, 319, 111–116. [Google Scholar] [CrossRef]
- Sofonia, J.J.; Anthony, K.R.N. High-sediment tolerance in the reef coral Turbinaria mesenterina from the inner Great Barrier Reef lagoon (Australia). Estuar Coast Shelf Sci. 2008, 78, 748–752. [Google Scholar] [CrossRef]
- Mellin, C.; Thompson, A.; Jonker, M.J.; Emslie, M.J. Cross-shelf variation in coral community response to disturbance. Diversity (Basel) 2019, 11, 38. [Google Scholar] [CrossRef]
- Baird, A.H.; Hughes, T.P. Competitive dominance by tabular corals: An experimental analysis of recruitment and survival of understorey assemblages. J. Exp. Mar. Biol. Ecol. 2000, 251, 117–132. [Google Scholar] [CrossRef]
- Torda, G.; Sambrook, K.; Cross, P.; Sato, Y.; Bourne, D.G.; Lukoschek, V.; Hill, T.; Torras Jorda, G.; Moya, A.; Willis, B.L.; et al. Decadal erosion of coral assemblages by multiple disturbances in the Palm Islands, central Great Barrier Reef. Sci. Rep. 2018, 8, 11885. [Google Scholar] [CrossRef]
- Loya, Y.; Sakai, K.; Yamazato, K.; Nakano, Y.; Sambali, H.; van Woesik, R. Coral bleaching: the winners and losers. Ecol. Lett. 2001, 4, 122–131. [Google Scholar] [CrossRef]
- Van Woesik, R.; Sakai, K.; Ganase, A.; Loya, Y. Revisiting the winners and losers a decade after coral bleaching. Mar. Ecol. Prog. Ser. 2011, 434, 67–76. [Google Scholar] [CrossRef]
- Done, T.J.; Potts, D.C. Influences of habitat and natural disturbances on contributions of massive Porites coral to reef communities. Mar. Biol. 1992, 114, 479–493. [Google Scholar] [CrossRef]
- Marimuthu, N.; Jerald Wilson, J.; Vinithkumar, N.V.; Kirubagaran, R. Coral reef recovery status in South Andaman Islands after the bleaching event 2010. J. Ocean Univ. China 2013, 12, 91–96. [Google Scholar] [CrossRef]
- Schuhmacher, H.; Loch, K.; Loch, W.; See, W.R. The aftermath of coral bleaching on a Maldivian reef- a quantitative study. Facies 2005, 51, 80–92. [Google Scholar] [CrossRef]
- Baird, A.H.; Marshall, P.A. Mortality, growth and reproduction in scleractinian corals following bleaching on the Great Barrier Reef. Mar. Ecol. Prog. Ser. 2002, 237, 133–141. [Google Scholar] [CrossRef]
- Darling, E.S.; Alvarez-Filip, L.; Oliver, T.A.; McClanahan, T.R.; Cote, I.M. Evaluation life-history strategies of reef corals from species traits. Ecol. Lett. 2012, 15, 1378–1386. [Google Scholar] [CrossRef]
- Frade, P.; Bongaerts, P.; Englebert, N.; Rogers, A.; Gonzalez-Rivero, M.; Hoegh-Guldberg, O. Deep reefs of the Great Barrier Reef offer limited thermal refuge during mass coral bleaching. Nat. Commun. 2018, 9, 3447. [Google Scholar] [CrossRef]
- Babcock, R.C.; Baird, A.H.; Piromvaragorn, S.; Thomson, D.P.; Willis, B.L. Identification of scleractinian coral recruits from Indo-Pacific reefs. Zool. Stud. 2003, 42, 211–226. [Google Scholar]
- Ritson-Williams, R.; Arnold, S.N.; Paul, V.J. Patterns of larval settlement preferences and post-settlement survival for seven Caribbean corals. Mar. Ecol. Prog. Ser. 2016, 548, 127–138. [Google Scholar] [CrossRef]
- Szmant, A. Reproductive ecology of Caribbean reef corals. Coral Reefs 1986, 5, 43–53. [Google Scholar] [CrossRef]
- Hughes, T.P.; Baird, A.H.; Dinsdale, E.A.; Moltschaniwskyj, N.; Pratchett, M.S.; Tanner, J.E.; Willis, B.L. Patterns of recruitment and abundance of corals along the Great Barrier Reef. Nature 1999, 397, 59–63. [Google Scholar] [CrossRef] [Green Version]
- Davidson, J.; Thompson, A.; Logan, M.; Schaffelke, B. High spatio-temporal variability in Acroporidae settlement to inshore reefs of the Great Barrier Reef. PLoS ONE 2019, 14, e0209771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mumby, P.J.; Hastings, A.; Edwards, H.J. Thresholds and the resilience of Caribbean coral reefs. Nature 2007, 450, 98–101. [Google Scholar] [CrossRef]
- Kayal, M.; Vercelloni, J.; Lison de Loma, T.; Bosserelle, P.; Chancerelle, Y.; Geoffroy, S.; Stievenart, C.; Michonneau, F.; Penin, L.; Planes, S.; et al. Predator crown-of-thorns starfish (Acanthaster planci) outbreak, mass mortality of corals, and cascading effects on reef fish and benthic communities. PLoS ONE 2012, 7, e47363. [Google Scholar] [CrossRef] [PubMed]
- Dajka, J.-C.; Wilson, S.K.; Robinson, J.P.W.; Chong-Seng, K.M.; Harris, A.; Graham, N.A.J. Uncovering drivers of juvenile coral density following mass bleaching. Coral Reefs 2019. [Google Scholar] [CrossRef]
- Madin, J.S.; Baird, A.H.; Bridge, T.C.L.; Connolly, S.R.; Zawada, K.J.A.; Dornelas, M. Cumulative effects of cyclones and bleaching on coral cover and species richness at Lizard Island. Mar. Ecol. Prog. Ser. 2018, 604, 263–268. [Google Scholar] [CrossRef] [Green Version]





| Elements of Sampling Design | LTMP | MMP |
|---|---|---|
| Depth * | 6–9 m | 5 m |
| Sites | 3 | 2 |
| Transects | 5 | 5 |
| Length of benthic transects | 50 m | 20 m |
| Images per transect | 40 | 32 |
| Total points identified per reef | 3000 | 1600 |
| Total area of juvenile transects per reef | 25.5 m2 | 68 m2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jonker, M.J.; Thompson, A.A.; Menéndez, P.; Osborne, K. Cross-Shelf Variation Among Juvenile and Adult Coral Assemblages on Australia’s Great Barrier Reef. Diversity 2019, 11, 85. https://doi.org/10.3390/d11060085
Jonker MJ, Thompson AA, Menéndez P, Osborne K. Cross-Shelf Variation Among Juvenile and Adult Coral Assemblages on Australia’s Great Barrier Reef. Diversity. 2019; 11(6):85. https://doi.org/10.3390/d11060085
Chicago/Turabian StyleJonker, Michelle J., Angus A. Thompson, Patricia Menéndez, and Kate Osborne. 2019. "Cross-Shelf Variation Among Juvenile and Adult Coral Assemblages on Australia’s Great Barrier Reef" Diversity 11, no. 6: 85. https://doi.org/10.3390/d11060085
APA StyleJonker, M. J., Thompson, A. A., Menéndez, P., & Osborne, K. (2019). Cross-Shelf Variation Among Juvenile and Adult Coral Assemblages on Australia’s Great Barrier Reef. Diversity, 11(6), 85. https://doi.org/10.3390/d11060085