Effects of Land Cover Pattern Along Urban-Rural Gradient on Bird Diversity in Wetlands

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Bird Survey and Diversity Indices
2.3. Land Cover and Plant Community Survey
2.4. Statistical Analysis
3. Result
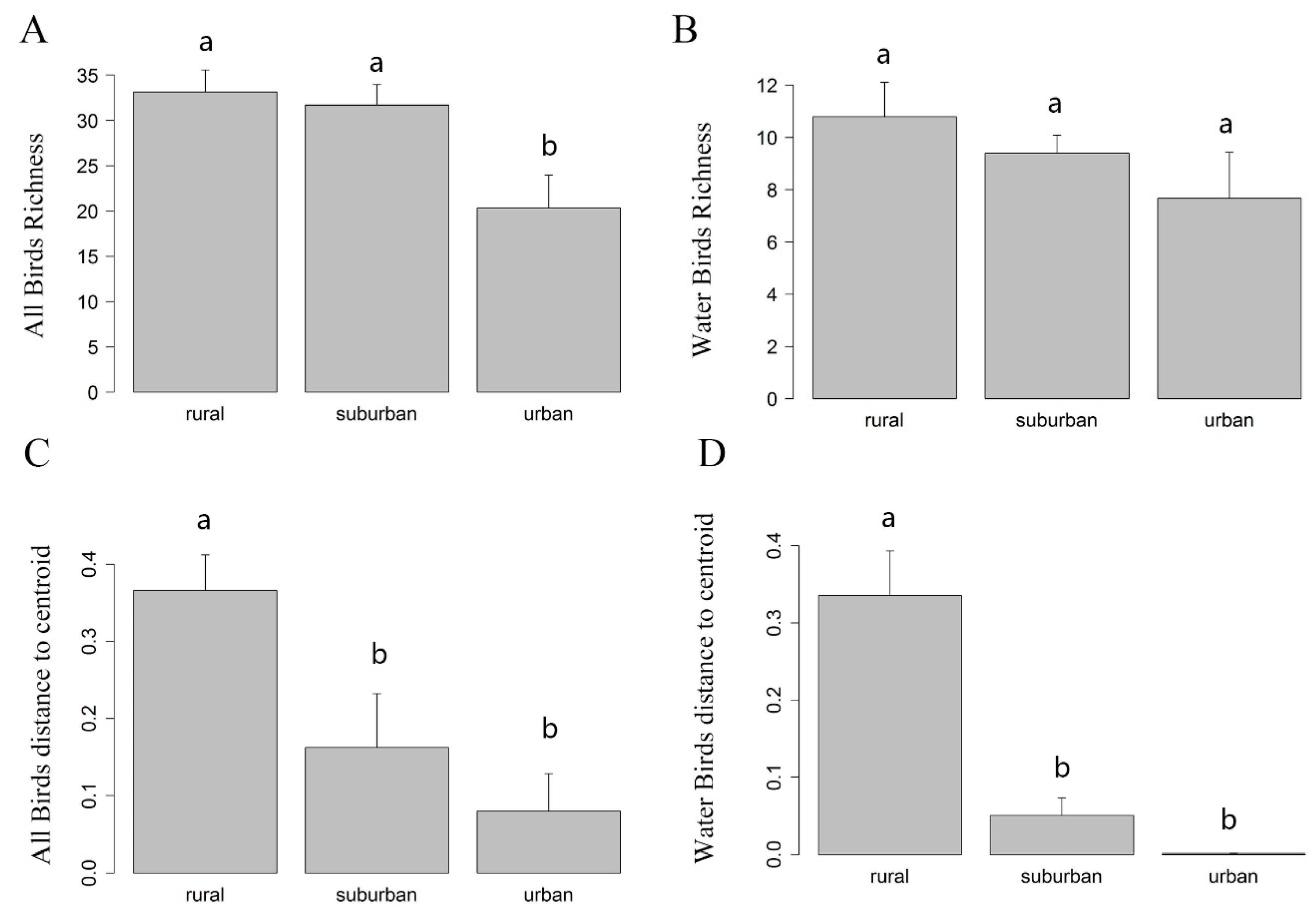
3.1. Differences of α Diversity in Urban-Rural Gradient
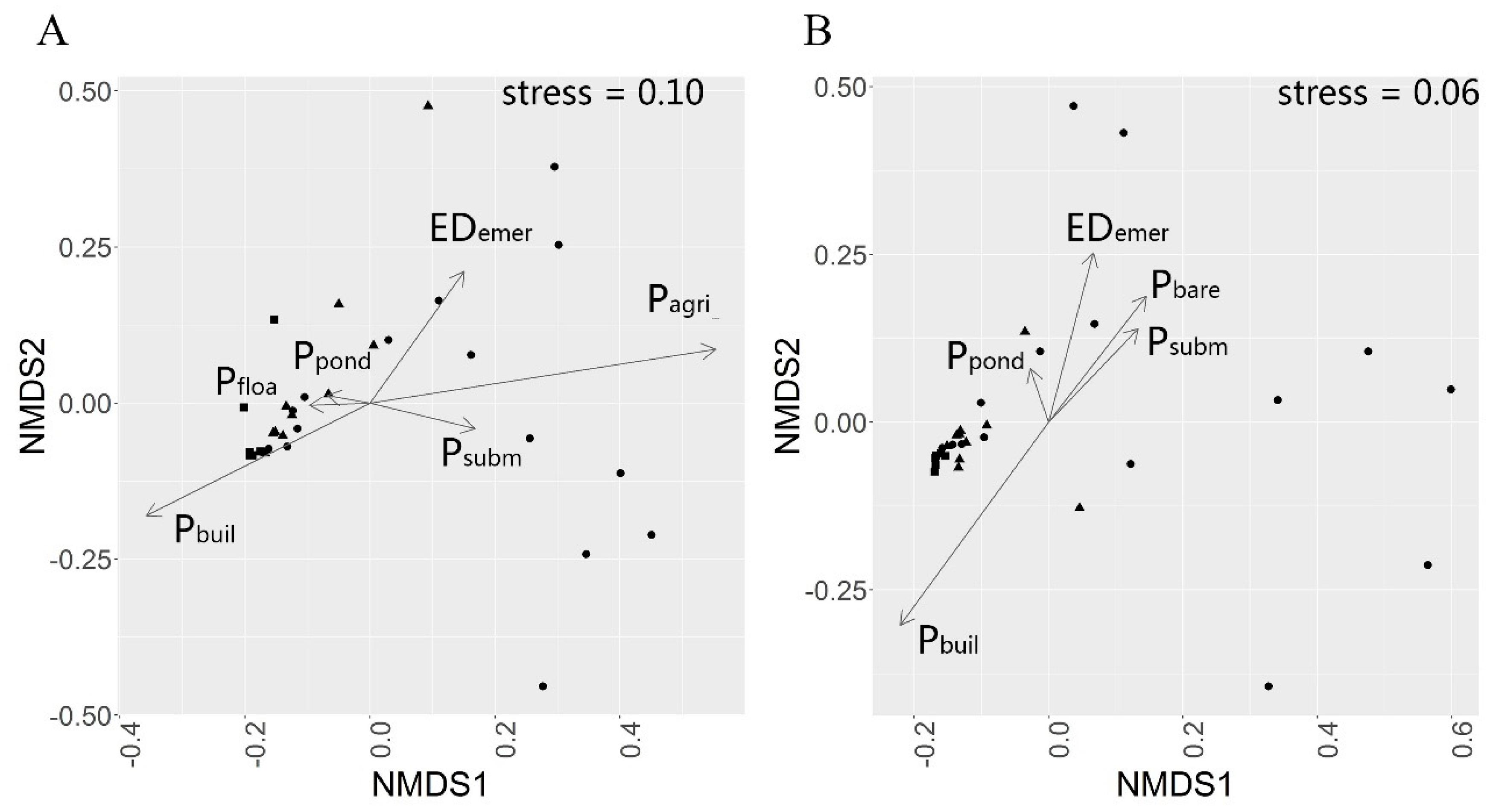
3.2. The Differences of β Diversities in Urban-Rural Gradient
3.3. Effects of Land Cover Patterns on Bird Species α Diversities
3.4. Effects of Land Cover Patterns on Bird Community Dispersion in Urban-Rural Gradient
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
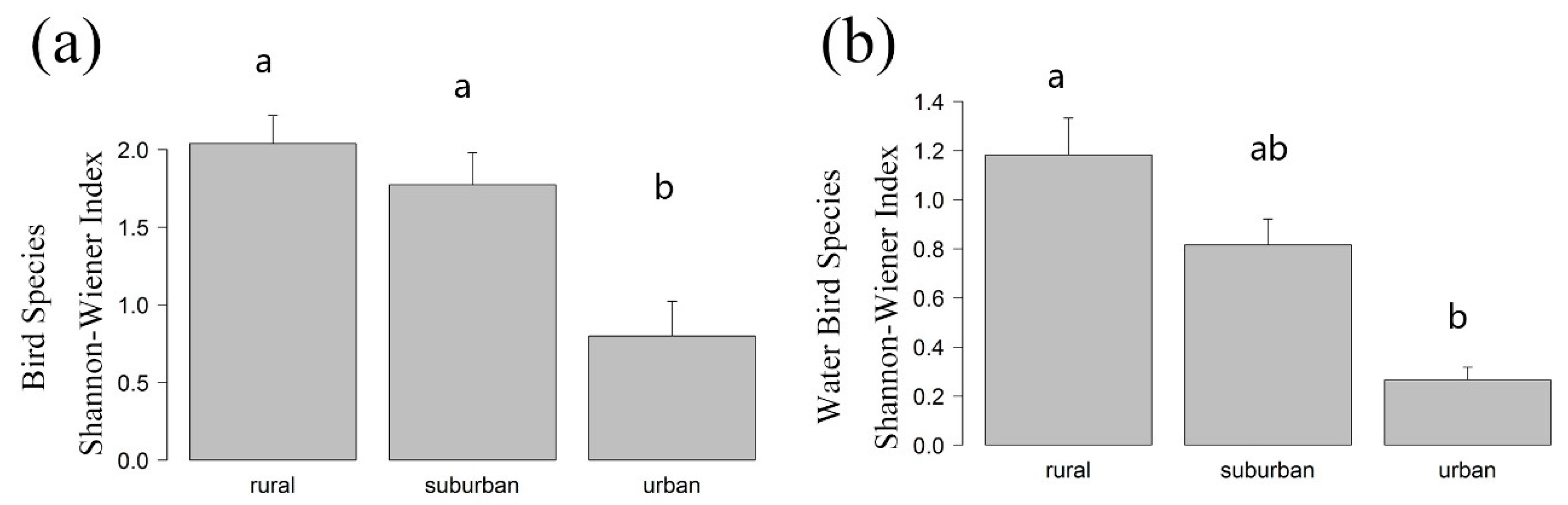
Appendix A. Differences in the Shannon-Wiener Index of the Urban-Rural Gradient


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | Type | Species Richness | Shannon-Wiener Index | ||
|---|---|---|---|---|---|
| Range | Mean ± S.E. | Range | Mean ± S.E. | ||
| Summer | All birds | 4–34 | 19.32 ± 1.32 | 0.42–2.98 | 1.97 ± 0.12 |
| Water birds | 0–15 | 5.00 ± 0.55 | 0–1.95 | 0.75 ± 0.09 | |
| Winter | All birds | 6–40 | 22.00 ± 1.35 | 0.10–2.71 | 1.27 ± 0.15 |
| Water birds | 1–15 | 7.74 ± 0.61 | 0–1.89 | 0.59 ± 0.10 | |
Appendix B
References
- Hu, S.; Niu, Z.; Chen, Y.; Li, L.; Zhang, H. Global wetlands: Potential distribution, wetland loss, and status. Sci. Total Environ. 2017, 586, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Davidson, N.C. How much wetland has the world lost? Long-term and recent trends in global wetland area. Mar. Freshw. Res. 2014, 65, 934–941. [Google Scholar] [CrossRef]
- Cohen, P.; Potchter, O.; Schnell, I. The impact of an urban park on air pollution and noise levels in the Mediterranean city of Tel-Aviv, Israel. Environ. Pollut. 2014, 195, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Song, J.; Yuan, H. Persistent organic pollutant residues in the sediments and mollusks from the Bohai Sea coastal areas, North China: An overview. Environ. Int. 2009, 35, 632–646. [Google Scholar] [CrossRef] [PubMed]
- Braga, E.S.; Bonetti, C.; Burone, L.; Bonetti, J. Eutrophication and bacterial pollution caused by industrial and domestic wastes at the Baixada Santista estuarine system - Brazil. Mar. Pollut. Bull. 2000, 40, 165–173. [Google Scholar] [CrossRef]
- Verzijden, M.N.; Ripmeester, E.A.P.; Ohms, V.R.; Snelderwaard, P.; Slabbekoorn, H. Immediate spectral flexibility in singing chiffchaffs during experimental exposure to highway noise. J. Exp. Biol. 2010, 213, 2575–2581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamer, A.J.; McDonnell, M.J. Amphibian ecology and conservation in the urbanising world: A review. Biol. Conserv. 2008, 141, 2432–2449. [Google Scholar] [CrossRef]
- Sewell, S.R.; Catterall, C.P. Bushland modification and styles of urban development: Their effects on birds in south-east Queensland. Wildl. Res. 1998, 25, 41–63. [Google Scholar] [CrossRef]
- Newbold, T.; Hudson, L.N.; Hill, S.L.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Boerger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45. [Google Scholar] [CrossRef] [PubMed]
- Dennis, R.L.H.; Hardy, P.B. Loss rates of butterfly species with urban development. A test of atlas data and sampling artefacts at a fine scale. Biodivers. Conserv. 2001, 10, 1831–1837. [Google Scholar] [CrossRef]
- Shochat, E.; Lerman, S.B.; Anderies, J.M.; Warren, P.S.; Faeth, S.H.; Nilon, C.H. Invasion, Competition, and Biodiversity Loss in Urban Ecosystems. Bioscience 2010, 60, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Moffatt, S.F.; McLachlan, S.M.; Kenkel, N.C. Impacts of land use on riparian forest along an urban-rural gradient in southern Manitoba. Plant Ecol. 2004, 174, 119–135. [Google Scholar] [CrossRef]
- Kark, S.; Iwaniuk, A.; Schalimtzek, A.; Banker, E. Living in the city: Can anyone become an “urban exploiter”? J. Biogeogr. 2007, 34, 638–651. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- McKinney, R.A.; Raposa, K.B.; Cournoyer, R.M. Wetlands as habitat in urbanizing landscapes: Patterns of bird abundance and occupancy. Landsc. Urban Plan. 2011, 100, 144–152. [Google Scholar] [CrossRef]
- Morelli, F.; Benedetti, Y.; Diego Ibanez-Alamo, J.; Jokimaki, J.; Mand, R.; Tryjanowski, P.; Moller, A.P. Evidence of evolutionary homogenization of bird communities in urban environments across Europe. Glob. Ecol. Biogeogr. 2016, 25, 1284–1293. [Google Scholar] [CrossRef]
- Clergeau, P.; Croci, S.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.-L.; Dinetti, M. Avifauna homogenisation by urbanisation: Analysis at different European latitudes. Biol. Conserv. 2006, 127, 336–344. [Google Scholar] [CrossRef]
- Blair, R.B. Creating a homogeneous avifauna. In Avian Ecology and Conservation in an Urbanizing World; Marzluff, J.M., Bowman, R., Donnelly, R., Eds.; Springer US: Boston, MA, USA, 2001; pp. 459–486. ISBN 978-1-4615-1531-9. [Google Scholar]
- Kühn, I.; Klotz, S. Urbanization and homogenization—Comparing the floras of urban and rural areas in Germany. Biol. Conserv. 2006, 127, 292–300. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization, Biodiversity, and ConservationThe impacts of urbanization on native species are poorly studied, but educating a highly urbanized human population about these impacts can greatly improve species conservation in all ecosystems. BioScience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Fortel, L.; Henry, M.; Guilbaud, L.; Guirao, A.L.; Kuhlmann, M.; Mouret, H.; Rollin, O.; Vaissiere, B.E. Decreasing Abundance, Increasing Diversity and Changing Structure of the Wild Bee Community (Hymenoptera: Anthophila) along an Urbanization Gradient. PLoS ONE 2014, 9, e104679. [Google Scholar] [CrossRef]
- Bates, A.J.; Sadler, J.P.; Fairbrass, A.J.; Falk, S.J.; Hale, J.D.; Matthews, T.J. Changing Bee and Hoverfly Pollinator Assemblages along an Urban-Rural Gradient. PLoS ONE 2011, 6, e23459. [Google Scholar] [CrossRef]
- Clucas, B.; Marzluff, J.M. A cross-continental look at the patterns of avian species diversity and composition across an urbanisation gradient. Wildl. Res. 2015, 42, 554–562. [Google Scholar] [CrossRef]
- Turner, M.G. Landscape ecology in North America: Past, present, and future. Ecology 2005, 86, 1967–1974. [Google Scholar] [CrossRef]
- Thornton, D.H.; Branch, L.C.; Sunquist, M.E. The influence of landscape, patch, and within-patch factors on species presence and abundance: A review of focal patch studies. Landsc. Ecol. 2011, 26, 7–18. [Google Scholar] [CrossRef]
- Colwell, M.A. Shorebird ecology, conservation, and management; Univ of California Press: Berkley, CA, USA, 2010; pp. 134–138. [Google Scholar]
- Bara, M.; Segura, L.N. Effect of Air Temperature and Water Depth on Bird Abundance: A Case Study of Rallidae and Anatidae in the Northeastern Algerian Garaet Hadj Tahar. Pak. J. Zool. 2019, 51, 211–217. [Google Scholar] [CrossRef]
- Xia, S.; Wang, Y.; Lei, G.; Liu, Y.; Lei, J.; XiuboYu; Wen, L.; Zhou, Y. Restriction of Herbivorous Waterbird Distributions in the Middle and Lower Yangtze River Floodplain in View of Hydrological Isolation. Wetlands 2017, 37, 79–88. [Google Scholar] [CrossRef]
- Petersen, K.L.; Westmark, A.S. Bird Use of Wetlands in a Midwestern Metropolitan Area in Relation to Adjacent Land Cover. Am. Midl. Nat. 2013, 169, 221–228. [Google Scholar] [CrossRef]
- Rosselli, L.; Stiles, F.G. Wetland habitats of the Sabana de Bogota Andean Highland Plateau and their birds. Aquat. Conserv. Mar. Freshw. Ecosyst. 2012, 22, 303–317. [Google Scholar] [CrossRef]
- Rodrigues, A.G.; Borges-Martins, M.; Zilio, F. Bird diversity in an urban ecosystem: The role of local habitats in understanding the effects of urbanization. Iheringia Ser. Zool. 2018, 108, e2018017. [Google Scholar] [CrossRef]
- Whited, D.; Galatowitsch, S.; Tester, J.R.; Schik, K.; Lehtinen, R.; Husveth, J. The importance of local and regional factors in predicting effective conservation - Planning strategies for wetland bird communities in agricultural and urban landscapes. Landsc. Urban Plan. 2000, 49, 49–65. [Google Scholar] [CrossRef]
- Paz Silva, C.; Garcia, C.E.; Estay, S.A.; Barbosa, O. Bird Richness and Abundance in Response to Urban Form in a Latin American City: Valdivia, Chile as a Case Study. PLoS ONE 2015, 10, e0138120. [Google Scholar]
- Albanese, G.; Davis, C.A. Characteristics within and around stopover wetlands used by migratory shorebirds: Is the neighborhood important? Condor 2015, 117, 328–340. [Google Scholar] [CrossRef]
- Shwartz, A.; Shirley, S.; Kark, S. How do habitat variability and management regime shape the spatial heterogeneity of birds within a large Mediterranean urban park? Landsc. Urban Plan. 2008, 84, 219–229. [Google Scholar] [CrossRef]
- Menon, M.; Devi, P.M.; Rangaswamy, M. Avifaunal Richness and Abundance along an Urban Rural Gradient with Emphasis on Vegetative and Anthropogenic Attributes in Tiruchirappalli, India. Landsc. Res. 2016, 41, 131–148. [Google Scholar] [CrossRef]
- Chong, K.Y.; Teo, S.; Kurukulasuriya, B.; Chung, Y.F.; Rajathurai, S.; Tan, H.T.W. Not all green is as good: Different effects of the natural and cultivated components of urban vegetation on bird and butterfly diversity. Biol. Conserv. 2014, 171, 299–309. [Google Scholar] [CrossRef]
- Jost, L.; Chao, A.; Chazdon, R.L. Compositional similarity and β (beta) diversity. Biol. Divers. Front. Meas. Assess. 2011, 66–84. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community ecology package. R package version 1.17-4. R-Proj. Org Acess. Em 2010, 23, 2010. [Google Scholar]
- Agisoft Agisoft PhotoScan Unser Manual: Professional Edition, Version 1.2. 2016. Available online: https://www.agisoft.com/pdf/photoscan-pro_1_2_en.pdf (accessed on 28 April 2019).
- Trimble ECognition Developer 8.7 Reference Book. 2011. Available online: http://www.ecognition.com/ (accessed on 28 April 2019).
- ESRI ArcGIS Desktop: Release 10. Environmental Systems Research Institute: Redlands, CA, USA, 2011.
- McGarigal, K.; Marks, B.J. FRAGSTATS: Spatial Pattern Analysis Program for Quantifying Landscape Structure; For. Serv. Gen. Tech. Rep. PNW; USDA, Forest Service, Pacific Northwest Research Station: Washington, DC, USA, 1995; Volume 351. [Google Scholar]
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.r-project.org/ (accessed on 28 April 2019).
- Anderson, M.J. Distance-Based Tests for Homogeneity of Multivariate Dispersions. Biometrics 2006, 62, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J.; Ellingsen, K.E.; McArdle, B.H. Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 2006, 9, 683–693. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference; Springer: New York, NY, USA, 2002; pp. 75,131,323,324. ISBN 978-1-280-00948-8. [Google Scholar]
- Bickford, D.; Ng, T.H.; Qie, L.; Kudavidanage, E.P.; Bradshaw, C.J. Forest fragment and breeding habitat characteristics explain frog diversity and abundance in Singapore. Biotropica 2010, 42, 119–125. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Multimodel Inference: Understanding AIC and BIC in Model Selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Turkheimer, F.E.; Hinz, R.; Cunningham, V.J. On the Undecidability among Kinetic Models: From Model Selection to Model Averaging. J. Cereb. Blood Flow Metab. 2003, 23, 490–498. [Google Scholar] [CrossRef] [Green Version]
- Grueber, C.E.; Nakagawa, S.; Laws, R.J.; Jamieson, I.G. Multimodel inference in ecology and evolution: Challenges and solutions. J. Evol. Biol. 2011, 24, 699–711. [Google Scholar] [CrossRef] [PubMed]
- Bartoń, K. Multi-Model Inference. R Package, Version 1.43.6. Available online: https://cran.r-project.org/web/packages/MuMIn/MuMIn.pdf (accessed on 28 April 2019).
- Mazerolle, M.J. Model Selection and Multimodel Inference Based on (Q)AIC(c). R Package, Version 2.2-1. Available online: https://cran.r-project.org/web/packages/AICcmodavg/AICcmodavg.pdf (accessed on 28 April 2019).
- Tylianakis, J.M.; Klein, A.-M.; Tscharntke, T. Spatiotemporal variation in the diversity of Hymenoptera across a tropical habitat gradient. Ecology 2005, 86, 3296–3302. [Google Scholar] [CrossRef]
- Flohre, A.; Fischer, C.; Aavik, T.; Bengtsson, J.; Berendse, F.; Bommarco, R.; Ceryngier, P.; Clement, L.W.; Dennis, C.; Eggers, S.; et al. Agricultural intensification and biodiversity partitioning in European landscapes comparing plants, carabids, and birds. Ecol. Appl. 2011, 21, 1772–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paz Silva, C.; Sepulveda, R.D.; Barbosa, O. Nonrandom filtering effect on birds: Species and guilds response to urbanization. Ecol. Evol. 2016, 6, 3711–3720. [Google Scholar] [CrossRef] [PubMed]
- Luo, K. Communities and Habitat Selection of Wetland Birds in the Lakeshore-Wetland Around the Dian Lake. Master’s Thesis, Yunnan University, Kunming, China, May 2014. [Google Scholar]
- Niemuth, N.D.; Estey, M.E.; Reynolds, R.E.; Loesch, C.R.; Meeks, W.A. Use of wetlands by spring-migrant shorebirds in agricultural landscapes of North Dakota’s Drift Prairie. Wetlands 2006, 26, 30–39. [Google Scholar] [CrossRef]
- Buler, J.J.; Moore, F.R.; Woltmann, S. A multi-scale examination of stopover habitat use by birds. Ecology 2007, 88, 1789–1802. [Google Scholar] [CrossRef] [PubMed]
- Sushinsky, J.R.; Rhodes, J.R.; Possingham, H.P.; Gill, T.K.; Fuller, R.A. How should we grow cities to minimize their biodiversity impacts? Glob. Chang. Biol. 2013, 19, 401–410. [Google Scholar] [CrossRef] [PubMed]



| (Int) | Pagri | Pbuil | EDemer | Ppond | Psubm | Pfloa | AICc | ΔAICc | Weight |
|---|---|---|---|---|---|---|---|---|---|
| 28.20 | 0.217 | −0.192 | 0.071 | 218.6 | 0.00 | 0.290 | |||
| 30.58 | −0.265 | 0.463 | 2.570 | 219.5 | 0.89 | 0.186 | |||
| 30.43 | 0.212 | −0.266 | 0.424 | 220.0 | 1.48 | 0.138 | |||
| 29.63 | −0.206 | 0.633 | 2.041 | 220.1 | 1.51 | 0.136 | |||
| 31.13 | −0.229 | 0.070 | 220.1 | 1.54 | 0.134 | ||||
| 35.26 | −0.286 | 2.967 | −1.341 | 220.4 | 1.83 | 0.116 |
| (Int) | Pbare | Ppond | Psubm | EDemer | Pbuil | AICc | ΔAICc | Weight |
|---|---|---|---|---|---|---|---|---|
| 6.52 | 0.792 | 0.178 | 1.658 | 176.9 | 0.00 | 0.204 | ||
| 7.73 | 0.749 | 1.572 | 177.0 | 0.10 | 0.193 | |||
| 6.94 | 0.654 | 1.358 | 0.022 | 177.5 | 0.64 | 0.148 | ||
| 7.63 | 1.192 | 0.027 | 178.1 | 1.27 | 0.108 | |||
| 10.15 | 1.172 | −0.064 | 178.4 | 1.55 | 0.094 | |||
| 8.87 | 0.616 | 1.368 | −0.044 | 178.5 | 1.57 | 0.093 | ||
| 8.75 | 1.433 | 178.7 | 1.79 | 0.083 | ||||
| 9.11 | 0.167 | 1.238 | −0.066 | 178.8 | 1.93 | 0.078 |
| Parameter | Estimate | Unconditional SE | Confidence Interval | Relative Importance | Pr(>|z|) |
|---|---|---|---|---|---|
| (Intercept) | 30.357 | <0.001 | |||
| Pbuil | −0.234 | 0.085 | (−0.400, −0.068) | 1.00 | 0.008 |
| EDemer | 0.069 | 0.031 | (0.007, 0.130) | 0.56 | 0.036 |
| Psubm | 2.511 | 1.302 | (−0.042, 5.063) | 0.44 | 0.065 |
| Pagri | 0.215 | 0.108 | (0.004, 0.426) | 0.43 | 0.056 |
| Ppond | 0.446 | 0.216 | (0.023, 0.870) | 0.32 | 0.048 |
| Pfloa | −1.341 | 0.696 | (−2.704, 0.022) | 0.12 | 0.066 |
| Parameter | Estimate | Unconditional SE | Confidence Interval | Relative Importance | Pr(>|z|) |
|---|---|---|---|---|---|
| (Intercept) | 7.878 | <0.001 | |||
| Psubm | 0.722 | 0.656 | (0.138, 2.708) | 1.00 | 0.037 |
| Pbare | 0.175 | 0.371 | (−0.005, 1.448) | 0.64 | 0.063 |
| Ppond | 1.423 | 0.109 | (−0.039, 0.389) | 0.28 | 0.126 |
| Pbuil | 0.024 | 0.040 | (−0.137, 0.021) | 0.26 | 0.168 |
| EDemer | −0.058 | 0.016 | (−0.006, 0.055) | 0.26 | 0.137 |
| Statistic Value | Pbuil | Psubm | EDemer | Ppond |
|---|---|---|---|---|
| F-value | 19.337 | 3.189 | 3.450 | 3.780 |
| p-value | <0.001 | 0.056 | 0.046 | 0.035 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, Q.; Liao, C.; Wu, Z.; Guan, W.; Yang, W.; Tang, Y.; Wu, G. Effects of Land Cover Pattern Along Urban-Rural Gradient on Bird Diversity in Wetlands. Diversity 2019, 11, 86. https://doi.org/10.3390/d11060086
Mao Q, Liao C, Wu Z, Guan W, Yang W, Tang Y, Wu G. Effects of Land Cover Pattern Along Urban-Rural Gradient on Bird Diversity in Wetlands. Diversity. 2019; 11(6):86. https://doi.org/10.3390/d11060086
Chicago/Turabian StyleMao, Qian, Chencan Liao, Zhaolu Wu, Wenbo Guan, Wenda Yang, Yuqin Tang, and Gang Wu. 2019. "Effects of Land Cover Pattern Along Urban-Rural Gradient on Bird Diversity in Wetlands" Diversity 11, no. 6: 86. https://doi.org/10.3390/d11060086
APA StyleMao, Q., Liao, C., Wu, Z., Guan, W., Yang, W., Tang, Y., & Wu, G. (2019). Effects of Land Cover Pattern Along Urban-Rural Gradient on Bird Diversity in Wetlands. Diversity, 11(6), 86. https://doi.org/10.3390/d11060086