Two New Marine Free-Living Nematodes from Jeju Island Together with a Review of the Genus Gammanema Cobb 1920 (Nematoda, Chromadorida, Selachinematidae)



Abstract
:1. Introduction
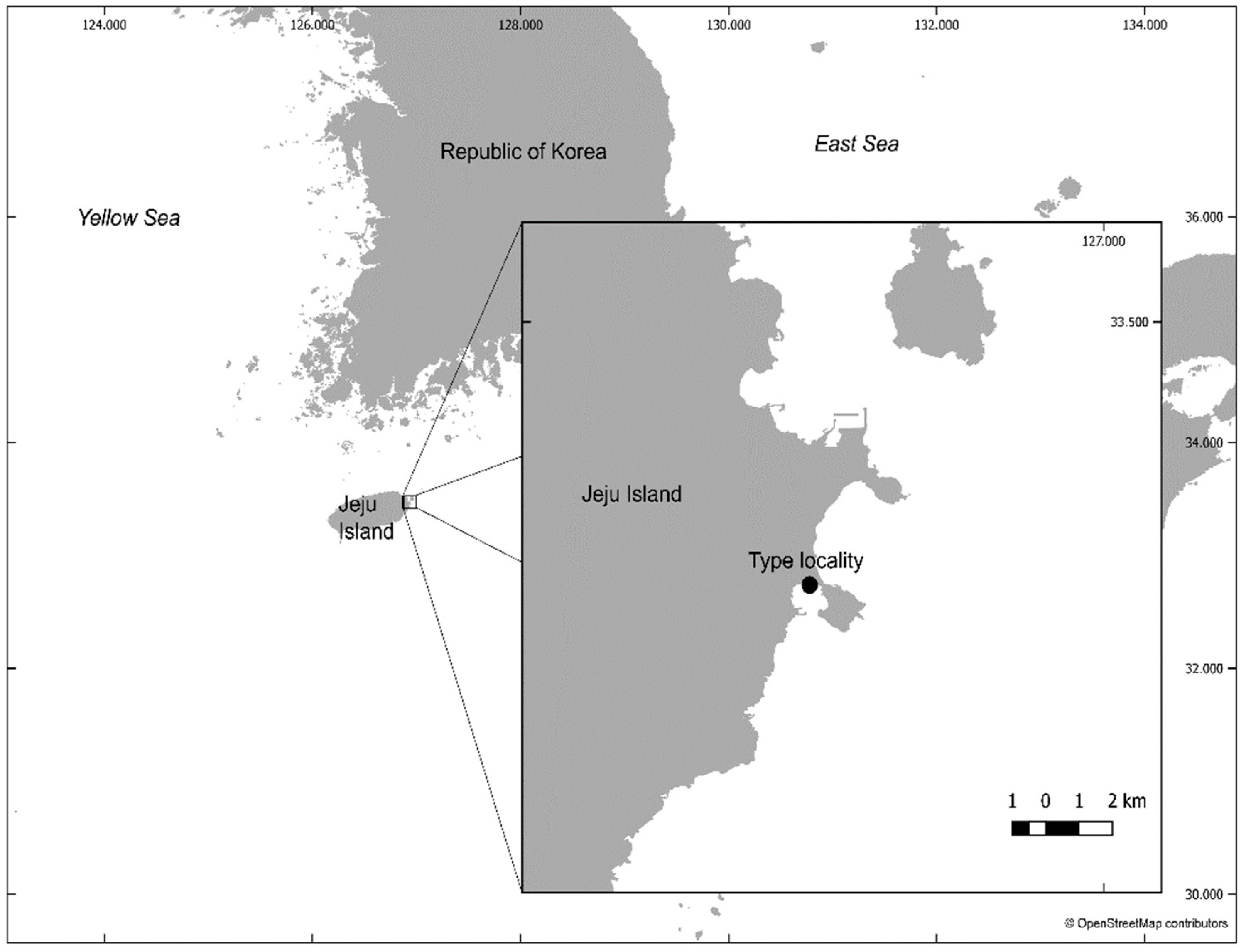
2. Materials and Methods
3. Results—Systematics
3.1. Review of Genus Gammanema
3.1.1. Diagnosis
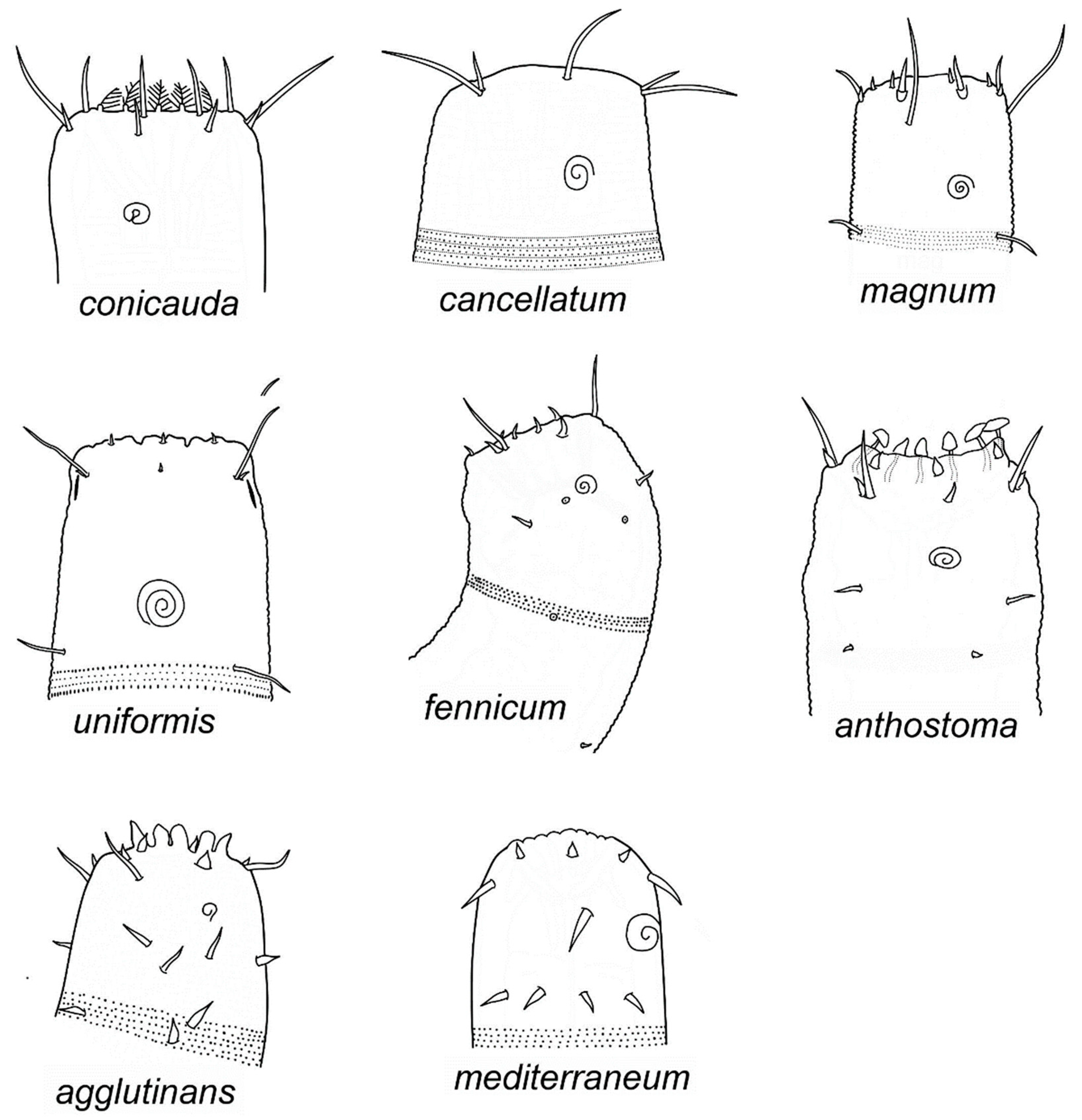
- Gammanema agglutinans Leduc, 2013. Leduc 2013:21‒25, Figures 11‒13, Table 2 [13] (males, females); SW Pacific, region to the east of New Zealand, Chatham Rise, depths 350‒1238 m.
- Gammanema anthostoma Okhlopkov, 2002. Okhlopkov 2002:42‒45, Figures 2 and 3 [15] (males and females); White Sea (Northern Russia), Kandalaksha Bay, depth 14 m.
- Gammanema cancellatum Gerlach, 1955. Gerlach 1955:271‒272, Abbildung. 6 [16] (female); San Salvador, coastal groundwater. Remark: the species is retained here as valid, despite the absence of males in diagnosis, since it can be distinguished sound from other congeners by peculiar cuticle ornamentation with rods.
- Gammanema conicauda Gerlach, 1953. Gerlach 1953:553‒555, Abbildungen 17 a‒f [17] (males, females, juvenile); Tyrrhenian Sea, area of Naples, surf zone; North Sea coast.
- Gammanema curvata Gagarin et Klerman, 2007. Gagarin and Klerman 2007:780, Figure 2, Table 2 [18] (male, females); Mediterranean coast of Israel, depth 50 m, sandy sediment.
- Gammanema fennicum (Gerlach, 1953) Gerlach, 1964. Gerlach 1953:22–23, Abbildung 6 [19] (as Halichoanolaimus fennicus) (female, juvenile); east Baltic Sea. Gerlach 1964:37 [20] (as junior synonym of Gammanema rapax). Okhlopkov 2002:45‒46, Figures 4 and 5 [15] (males, females, juvenile); White Sea (Northern Russia), Kandalaksha Bay, depth 16 m.
- Gammanema ferox Cobb, 1920 sp. inq. Cobb 1920:291‒293, Figure 74 [21] (male, female); New Hebrides, coral sand. Though the species is described for both genders, the description lacks some necessary measurements, such as anterior setae and spicules; amphids are not depicted and not mentioned in the text (amphideal fovea possibly inconspicuous or reduced); the only illustration presents a head without an indication of gender and position in slide (possibly sublateral). Restitution of valid state is possible after redescription of new specimens from the type area; the species can be recognized through the prominent anterior projections of rugae.
- Gammanema kosswigi Gerlach, 1964. Gerlach 1964:38‒39, Abbildungen 11 a‒d [19] (male, juvenile); Maldive Islands, coastal groundwater, intertidal zone.
- Gammanema magnum Shi et Xu, 2018. Shi and Xu 2018, 3–9, Figures 1‒4, Table 1 [22] (males, females); East China Sea, intertidal sandy sediment.
- Gammanema mediterraneum Vitiello, 1970. Vitiello 1970:491‒493, planche XV, 30 a‒e [23] (as Gammanema mediterranea) (females, juveniles); West Mediterranean, depths 310‒650 m.
- Gammanema okhlopkoviTchesunov, Jeong et Lee sp. n. Present paper.
- Gammanema polydonta Murphy, 1965. Murphy 1965:176‒179, Figures 3A‒C and 4A‒H [25] (males, females); Chilean coast, depth 45 m.
- Gammanema rapax (Ssaweljev, 1912) Gerlach 1964 sp. inq. Ssaweljev 1912:122 [26] (as Halichoanolaimus rapax) (male, female); Barents Sea, Kolafjord, Palafjord, Mogilnojesee (lake with an intermediate layer of sea water on the Kildin Island in the Barents Sea), sand. Murphy (1965: 179 [25]) considered the species; dubious because the original description is not illustrated and type specimens are evidently lacking. Gerlach (1964 [20]) synonymized Halichoanolaimus menzeli Ditlevsen 1918, Gammanema ferox Cobb, 1920 and Gammanema fennicum (Gerlach, 1953) with Gammanema rapax.
- Gammanema smithi Murphy, 1964. Murphy 1964: 194‒198, Figure 3A,B,D,E, Figure 4, Figure 5B [27] (male); Puget Sound (NE Pacific), intertidal sand. Remark: there are some discrepancies in the original diagnosis. Caudal glands are described and depicted ([27], Figure 4) as incaudal (i.e., located entirely within tail), while the glands in Figure 5A are definitely excaudal (i.e., gland cell bodies protrude anteriorly and lie along the midgut). Supplementary organs are depicted from a medio-ventral view in Figure 3C, while the hind body in Figure 4 is shown laterally. Taking into account that the diagnosis is based on only one male (holotype), we consider Figures 3C and 5A as wrong and not pertaining to G. smithi.
- Gammanema tchesunovi Gagarin et Klerman, 2007. Gagarin and Klerman 2007:782‒785, Figure 3, Table 3 [18] (males, females); Mediterranean coast of Israel, depths 50‒55 m, sand.
- Gammanema uniformis (Cobb, 1920) Tchesunov et Okhlopkov, 2006. Cobb 1920:293‒294, Figure 75 [21] (as Trogolaimus uniformis) (seemingly, one male); Atlantic coast of the United States, New Hampshire. Tchesunov and Okhlopkov 2006:40‒43, Figure 12, Table 6 [28] (males, females); Atlantic coast of the United States, Maine.
3.1.2. Remarks on the Species List and Synonymy
3.1.3. Identification of Species of Gammanema
3.2. Description of New Species of Gammanema
3.2.1. Etymology
3.2.2. Material Examined
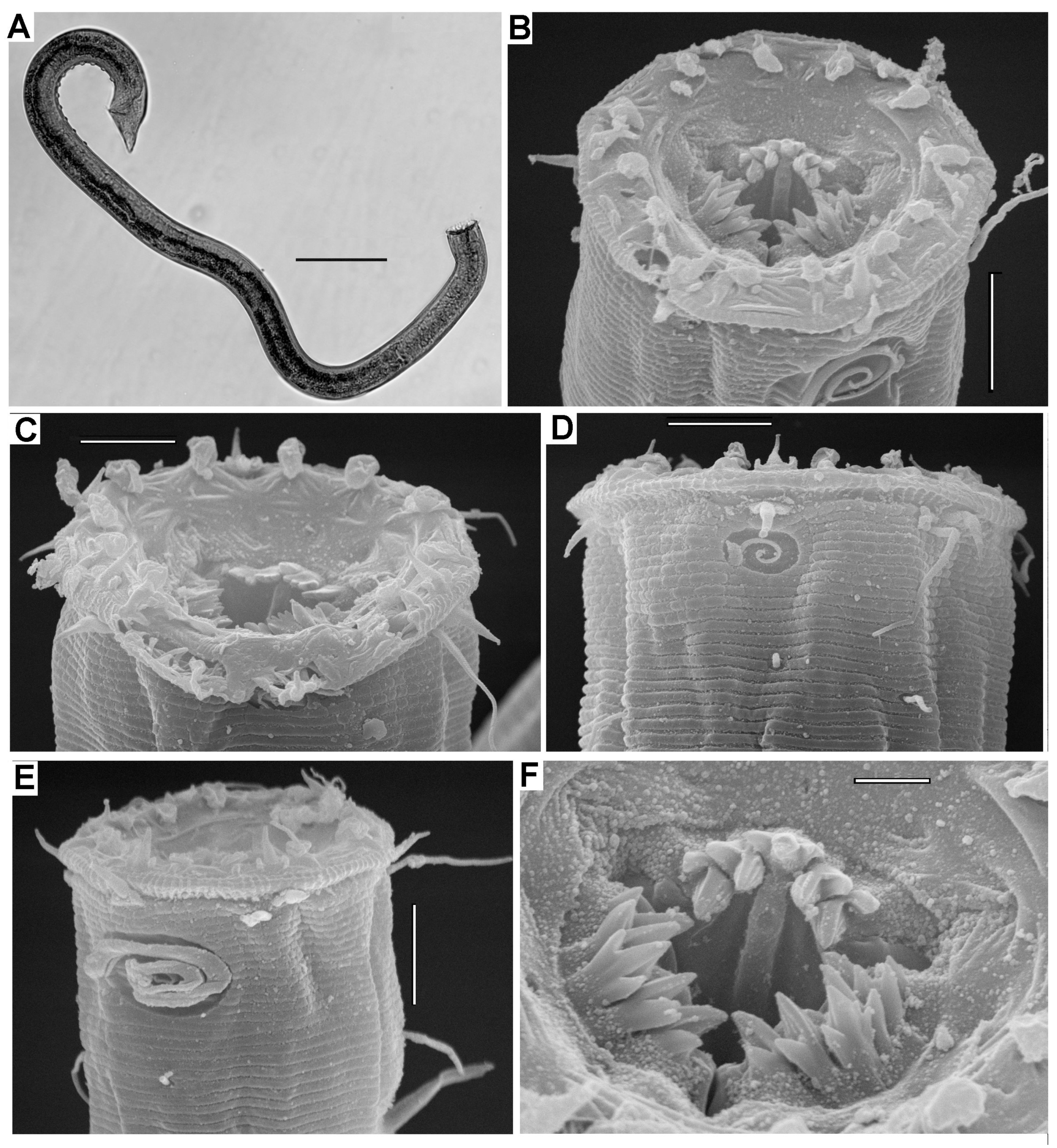
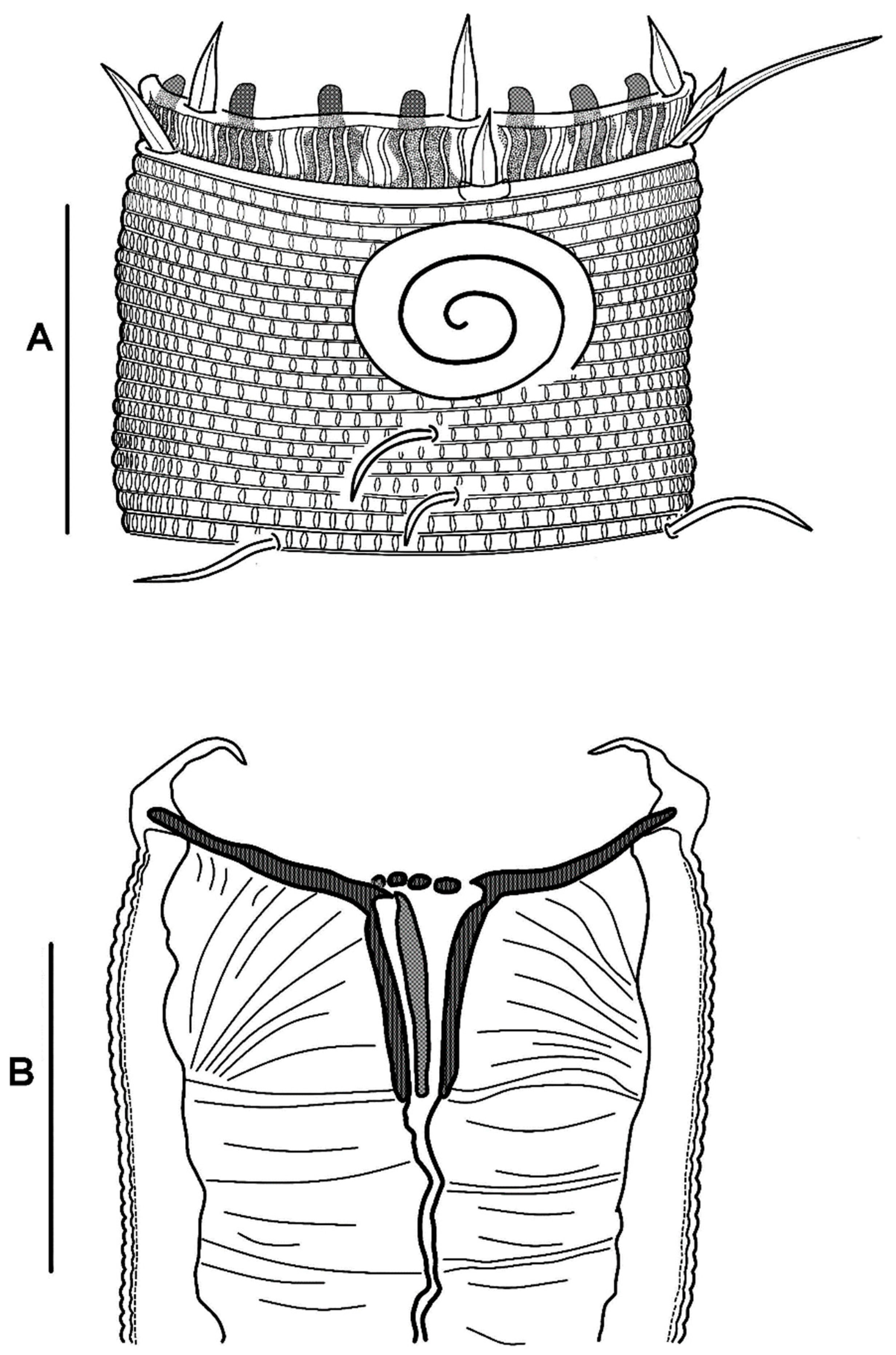
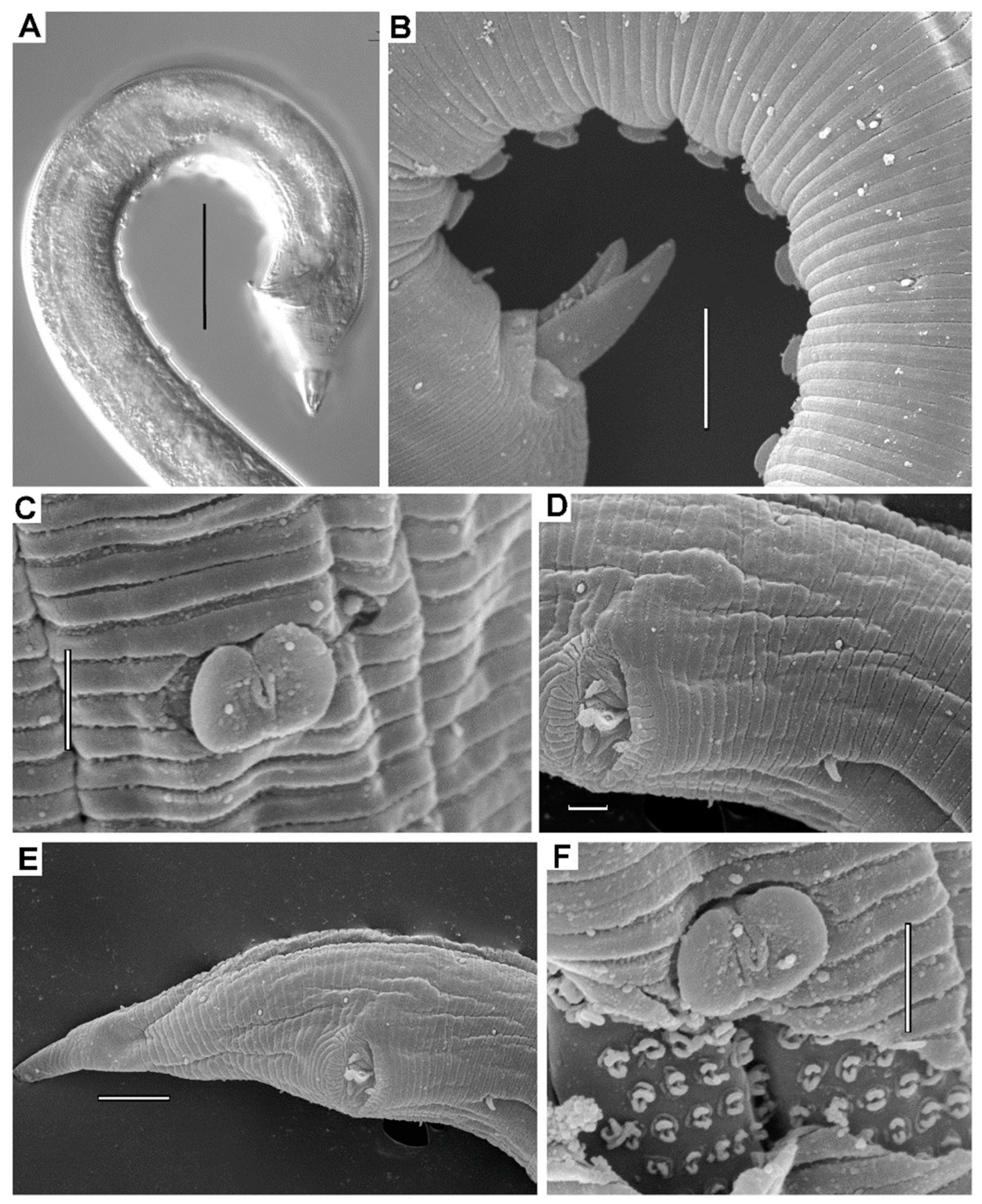
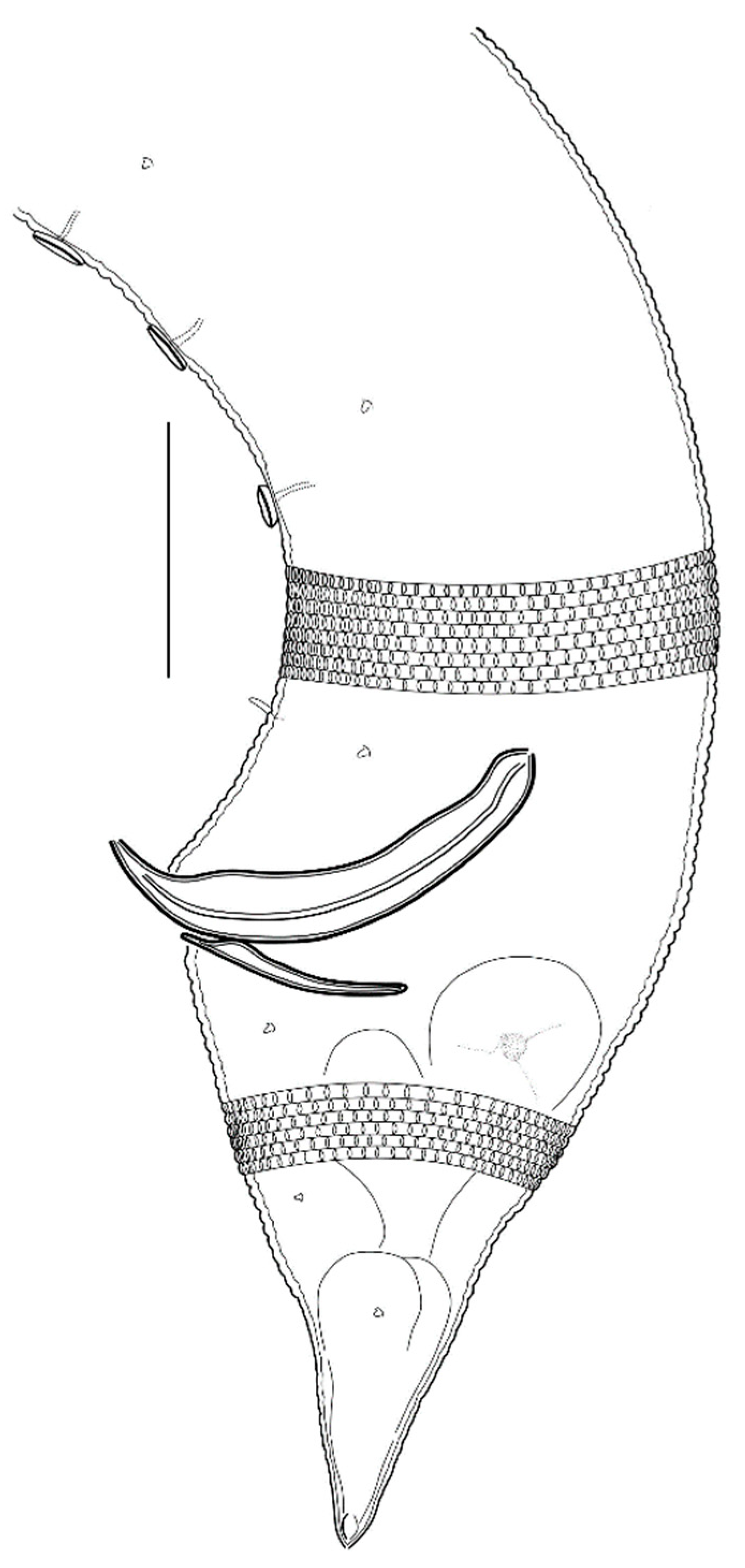
3.2.3. Description
3.2.4. Diagnosis
3.2.5. Differential Diagnosis
3.3. Description of New Species of Latronema
3.3.1. Etymology
3.3.2. Material Examined
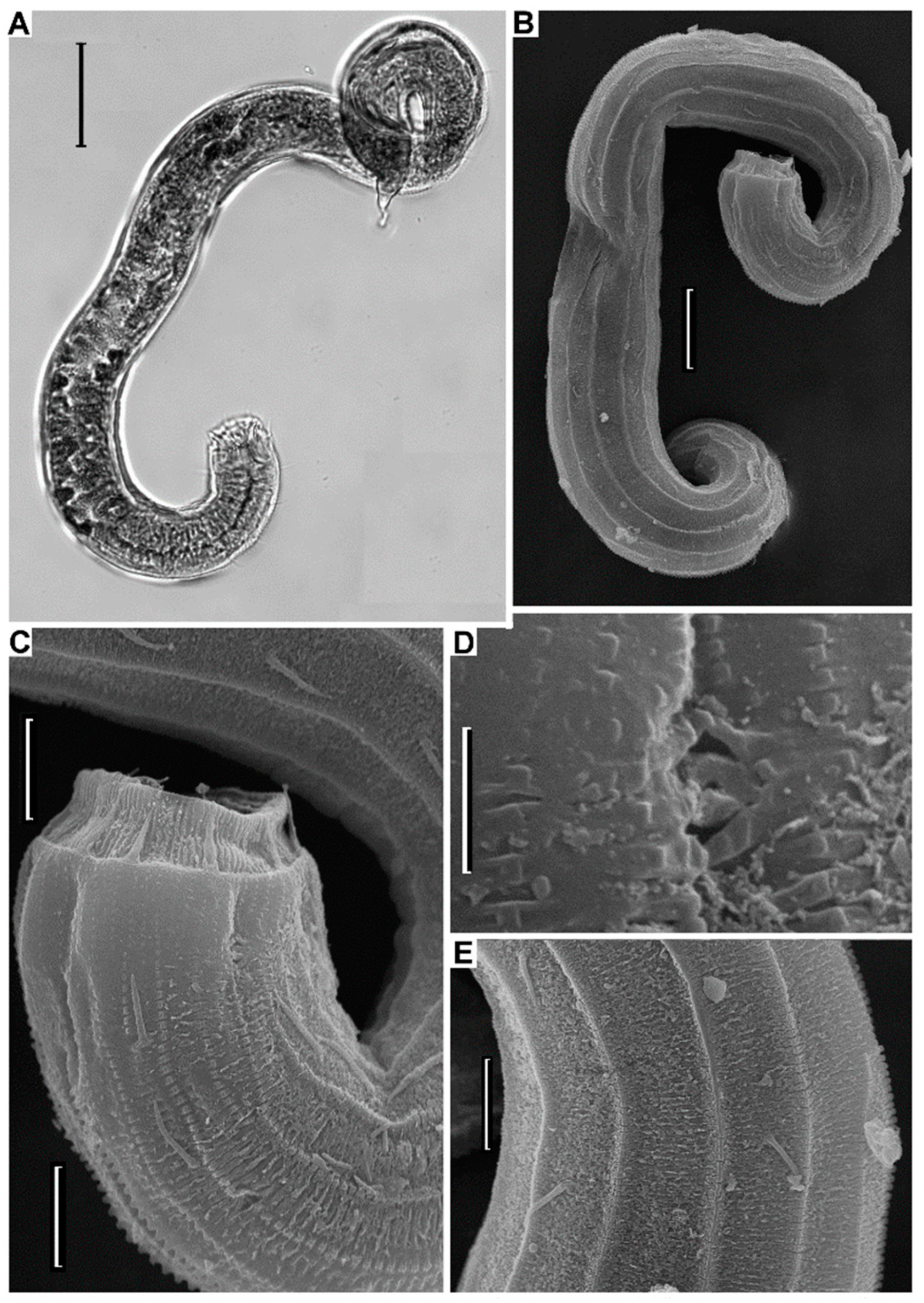
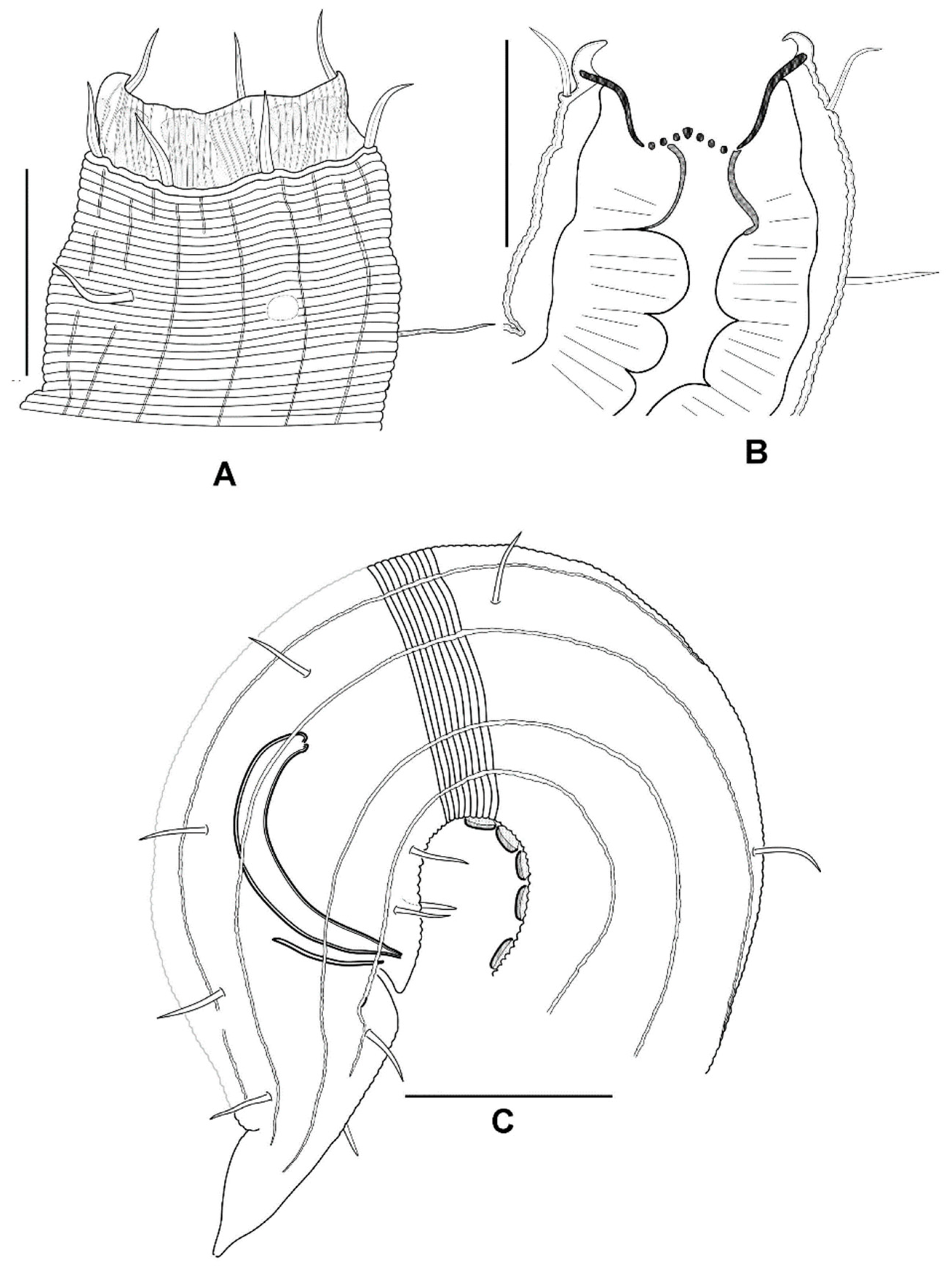
3.3.3. Description
3.3.4. Diagnosis
3.3.5. Differential Diagnosis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| a | body length divided by body diameter at midbody |
| b | body length divided by pharynx length |
| c | body length divided by tail length |
| c’ | tail length divided by anal body diameter |
| CV | coefficient of variation (SD divided by mean, in %) |
| SD | standard deviation |
| V | distance from anterior end to vulva in % of entire body length |
References
- Rho, H.S.; Kim, W. Paradraconema jejuense, a new species of genus Paradraconema (Nematoda: Draconematidae) from Korea. Korean J. Syst. Zool. 2004, 21, 81–91. [Google Scholar]
- Rho, H.S.; Kim, W. A new free-living marine nematode species of the genus Dracogalerus Allen and Noffsinger (Nematoda: Draconematidae) from a shallow subtidal zone of Jeju Island, Korea. Integr. Biosci. 2005, 9, 113–122. [Google Scholar] [CrossRef]
- Rho, H.S.; Decraemer, W.; Sørensen, M.V.; Min, W.G.; Jung, J.; Kim, W. Megadraconema cornutum, a new genus and species from Korea, with a discussion of its classification and relationships within the family Draconematidae (Nematoda, Desmodorida) based on morphological and molecular characters. Zool. Sci. 2011, 28, 68–84. [Google Scholar] [CrossRef]
- Lim, H.W.; Chang, C.Y. First record of Desmoscolex Nematoda (Desmoscolecida: Desmoscolecidae) from Korea. Integr. Biosci. 2006, 10, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Jeong, R.; Tchesunov, A.V.; Lee, W. Bibliographic revision of Mesacanthion Filipjev, 1927 (Nematoda: Thoracostomopsidae) with description of a new species from Jeju Island, South Korea. PeerJ 2019, 7, e8023. [Google Scholar] [CrossRef] [PubMed]
- Jeong, R.; Tchesunov, A.V.; Lee, W. Two species of Thoracostomopsidae (Nematoda: Enoplida) from Jeju Island, South Korea. PeerJ, submitted, under review.
- Okhlopkov, J.R. Feeding of free-living nematodes of the families Selachinematidae and Richtersiidae in the White Sea. Proc. Pertsov White Sea Biol. Stn. 2003, 9, 127–139. (In Russian) [Google Scholar]
- Moens, T.; Braeckman, U.; Derycke, S.; Fonseca, F.; Galucci, F.; Gingold, R.; Guilini, K.; Ingels, J.; Leduc, D.; Vanaverbeke, J.; et al. Ecology of free-living marine nematodes. In Handbook of Zoology; Schmidt-Rhaesa, A., Ed.; De Gruyter: Berlin, Germany, 2014; Volume 2, pp. 109–152. [Google Scholar]
- Warwick, R.M. Nematode associations in the Exe estuary. J. Mar. Biol. Assoc. UK 1971, 51, 439–454. [Google Scholar] [CrossRef] [Green Version]
- Gallucci, F.; Steyaert, M.; Moens, T. Can field distributions of marine predacious nematodes be explained by sediment constraints on their foraging success? Mar. Ecol. Prog. Ser. 2005, 304, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Burgess, R. An improved protocol for separating meiofauna from sediments using colloidal silica soils. Mar. Ecol. Prog. Ser. 2001, 214, 161–165. [Google Scholar] [CrossRef]
- Seinhorst, J. A rapid method for the transfer of nematodes from fixative to anhydrous glycerin. Nematologica 1959, 4, 67–69. [Google Scholar] [CrossRef] [Green Version]
- Leduc, D. Two new genera and five new species of Selachinematidae (Nematoda, Chromadorida) from the continental slope of New Zealand. Eur. J. Taxon. 2013, 63, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Tchesunov, A.V. Order Chromadorida Chitwood, 1933. In Handbook of Zoology; Schmidt-Rhaesa, A., Ed.; Gastrotricha, Cycloneuralia and Gnathifera, Volume 2: Nematoda; De Gruyter: Berlin, Germany, 2014; pp. 373–398. [Google Scholar]
- Okhlopkov, J.R. Free-living nematodes of the families Selachinematidae and Richtersiidae in the White Sea (Nematoda, Chromadoria). Zoosyst. Ross. 2002, 11, 41–55. [Google Scholar]
- Gerlach, S.A. Zur Kenntnis der freilebenden marinen Nematoden von San Salvador. Z. für Wiss. Zool. 1955, 158, 249–303. [Google Scholar]
- Gerlach, S.A. Die Nematodenbesiedlung des Sandstrandes und des Küstengrundwassers an der italienischen Küste. I. Systematischer Teil. Arch. Zool. Ital. 1953, 37, 517–640. [Google Scholar]
- Gagarin, V.G.; Klerman, A.K. New species of predatory chromadorids (Nematoda, Chromadorida) from the Mediterranean Sea. Zool. Zhurnal 2007, 86, 778–786, (In Russian with English Abstract). [Google Scholar]
- Gerlach, S.A. Die Nematodenfauna der Uferzonen und des Küstengrundwassers am finnischen Meeresbusen. Acta Zool. Fenn. 1953, 73, 1–32. [Google Scholar]
- Gerlach, S.A. Revision der Choniolaimidae und Selachinematidae (Freilebende Meeres-Nematoden); Kosswig-Festschrift; Mitteilungen aus dem Hamburgischen Zoologischen Museum und Institut: Hamburg, Germany, 1964; pp. 23–50. [Google Scholar]
- Cobb, N.A. One hundred new nemas (type species of 100 new genera). Contrib. Sci. Nematol. 1920, 9, 217–343. [Google Scholar]
- Shi, B.; Xu, K. Two new rapacious nematodes from intertidal sediments, Gammanema magnum sp. nov. and Synonchium caudatubatum sp. nov. (Nematoda, Selachinematidae). Eur. J. Taxon. 2018, 405, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Vitiello, P. Nématodes libres marins des vases profondes du Golfe du Lion. II. Chromadorida. Tethys 1970, 2, 449–500. [Google Scholar]
- Ditlevsen, H. Marine freeliving nematodes from Danish waters. Vidensk. Meddelelser Dansk Naturhistorisk Foren. I Kjobenhavn 1918, 70, 147–214. [Google Scholar]
- Murphy, D.G. Chilean marine nematodes. Veröff. Inst. für Meeresforsch. Bremerhav. 1965, 9, 173–203. [Google Scholar]
- Ssaweljev, S. Zur Kenntnis der freilebenden Nematoden des Kolafjords und des Relictensee Mogilnoje. Trav. Soc. Imp. Nat. St. Pétersbg. 1912, 43, 108–126. [Google Scholar]
- Murphy, D.G. Free-living marine nematodes, I. Southerniella youngi, Dagda phinneyi, and Gammanema smithi, new species. Proc. Helminthol. Soc. Wash. 1964, 31, 190–198. [Google Scholar]
- Tchesunov, A.V.; Okhlopkov, Y.R. On some selachinematid nematodes (Chromadorida: Selachinematidae) deposited in the collection of the Smithsonian National Museum of Natural History. Nematology 2006, 8, 21–44. [Google Scholar]
- Platt, H.M. Pictorial taxonomic keys: Their construction and use for the identification of free-living marine nematodes. Cah. Biol. Mar. 1984, 25, 83–91. [Google Scholar]
- Leduc, D.; Zhao, Z. Latronema whataitai sp. n. (Nematoda: Selachinematidae) from intertidal sediments of New Zealand, with notes on relationships within the family based on preliminary 18S and D2-D3 phylogenetic analyses. Nematology 2015, 17, 941–952. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Characters | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Body Length | a | Inner Labial Setae Length | Outer Labial Setae Length | Cephalic Setae Length | Amphid Width | Supplements Shape | Number of Supplements | Spicule Sength | Some Particular Bold Features | |
| agglutinans | 544‒696 | 10‒11 | 2 | 2 | 10‒11 | 10 | tubular | ≥6 | 34‒36 | Cuticle with minute spines, somatic setae leaf-shaped |
| anthostoma | 1002‒1950 | 22–29 | 5‒6 | 5‒8 | 6‒8 | 14‒26 | absent | absent | 62‒87 | Prominent circumoral projections each consisted of peduncle and triangular cap |
| conicauda | 1985‒2484 | 41‒48 | 16 | 17 | 30‒33 | 15 | conical papillae | 22 | 33‒40 | Prominent circumoral hair-like projections |
| curvata | 3286 | 76 | not indicated | 17 | 13 | 8‒8.5 | cup-shaped | 16 | 28 | - |
| fennicum | 2165‒2600 | 28‒49 | 17‒24 | 8‒11 | 34‒50 | 28‒31 | sucker-like | 13‒21 | 62‒64 | - |
| kosswigi | 1200 | 29 | 8 | 10‒12 | 20 | 30 | weak setose | 4 | 39 | Outer labial setae jointed |
| magnum | 2900–4085 | 48–52 | 15–16 | 22–25 | 31–35 | 42–45 | absent | absent | left 51‒56, right 62‒69 | Spicules slightly unequal |
| okhlopkovi | 961‒1150 | 25‒31 | 4‒5 | 3‒7 | 8‒19 | 11‒18 | sucker-shaped | 10‒13 | 32‒39 | Prominent circumoral Projections peg-like |
| polyodonta | 3200‒4200 | 21‒27 | not indicated | papillae | 16 | 42‒45 | cup-shaped | 43‒52 | 135 | Very massive appearance; Anterior half of body opaque because of densely pigmented intestine |
| smithi | 2210 | 41 | not indicated | 5 | 20 | 14 | cup-shaped | 20 | 78 | - |
| tchesunovi | 1603‒1889 | 15‒16 | not indicated | 5 | 8‒8 | 15‒18 | cup-shaped | 23‒24 | 85‒87 | Tail rounded and conical, terminal cone not developed |
| uniformis | 2700‒2950 | 45‒61 | 1‒2 | 2‒2 | 9‒10 | 10‒11 | cup-shaped | 19‒22 | 82‒87 | Cephalic cuticle thickened and internally sclerotized around anterior chamber of pharyngostome |
| Species | Characters | ||||||
|---|---|---|---|---|---|---|---|
| Body Length | a | Inner Labial Setae | Outer Labial Setae | Cephalic Setae | Amphid Width | Some Particular Bold Features | |
| agglutinans | 763‒918 | 11‒15 | 2 | 2 | 11‒19 | 5 | Cuticle with minute spines, somatic setae leaf-shaped |
| anthostoma | 1487‒1782 | 17‒24 | 5‒6.5 | 5‒7.5 | 15‒19 | 9.5‒10.5 | Prominent circumoral projections; each consisted of peduncle and triangular cap |
| cancellatum | 2465 | 24 | no data | 40 | 15 | 8 | Complex ornamentation of cuticle consists of dots and longitudinally arranged short rods |
| conicauda | 2492‒3724 | 30‒35 | 16 | 12 | 30‒33 | 8‒9 | Prominent circumoral hair-like projections |
| curvata | 2392‒2639 | 64‒71 | no data | 15‒17 | 12‒13 | 8‒8.5 | - |
| fennicum | 1672‒2782 | 13‒25 | 9‒13 | 10‒13 | 16‒37 | 9.5‒11 | - |
| magnum | 2540‒3145 | 28‒33 | 10‒14 | 20‒21 | 40‒45 | 9‒10 | - |
| mediterraneum | 660‒745 | 18‒20 | papillae | papillae | 7.5 | 5‒6 | - |
| polyodonta | 3730‒4050 | 23‒31 | papillae | papillae | 16 | 20 | Very massive appearance; anterior half of body opaque because of densely pigmented intestine |
| tchesunovi | 1562‒1682 | 11‒12 | papillae | 5 | 7‒8 | 16‒18 | Tail rounded and conical, terminal cone not developed |
| uniformis | 2260‒2790 | 40‒57 | 1‒2 | 2‒2 | 9‒12 | 9‒10 | Cephalic cuticle thickened and internally sclerotized around anterior chamber of pharyngostome |
| Character | Holotype | Total Number | Min.–Max. | Mean | SD | CV% |
|---|---|---|---|---|---|---|
| Body length | 961 | 8 | 961‒1150 | 1082 | 61.8 | 5.71 |
| pharynx length | 178 | 8 | 164‒207 | 185 | 15.1 | 8.15 |
| tail length | 48 | 7 | 42‒57.5 | 50.4 | 5.6 | 11.1 |
| body diameter at level of cs | 37 | 7 | 36.9‒43.7 | 39.2 | 2.40 | 6.12 |
| body diameter at level of nerve ring | 35 | 8 | 33.1‒40.0 | 36.8 | 2.32 | 6.30 |
| body diameter at level of cardia | 35 | 8 | 34.1‒41.0 | 36.5 | 2.45 | 6.72 |
| body diameter at level of midbody | 37 | 8 | 37.0‒42.0 | 38.8 | 2.54 | 6.65 |
| body diameter at level of cloaca | 36 | 8 | 36.0‒42.5 | 38.2 | 2.54 | 6.65 |
| a | 26 | 8 | 25.6‒31.1 | 27.8 | 2.17 | 7.80 |
| b | 5.4 | 8 | 5.40–6.56 | 5.88 | 0.41 | 6.97 |
| c | 20 | 7 | 18.5‒24.6 | 21.6 | 2.32 | 10.7 |
| c’ | 1.33 | 7 | 1.01‒1.53 | 1.31 | 0.16 | 12.2 |
| inner labial setae length | 3.5‒4 | 6 | 4.0‒5.0 | 4.27 | 0.39 | 9.13 |
| lateral outer labial setae length | 3.7 | 6 | 3.0‒7.2 | 5.22 | 1.63 | 31.2 |
| latero-median outer labial setae length | 5 | 5 | 5.0‒7.0 | 5.88 | 0.76 | 12.9 |
| cephalic setae length | 12 | 6 | 8.5‒19.0 | 14.1 | 4.01 | 28.5 |
| anterior cervical setae length | 7.6 | 7 | 7.6‒15.6 | 10.8 | 2.56 | 23.8 |
| amphid width | 16 | 7 | 11.0‒18.0 | 14.8 | 2.01 | 9.13 |
| amphid width/corresponding body diameter | 43 | 8 | 32.5‒47.7 | 37.7 | 5.22 | 13.9 |
| distance amphid—cephalic apex | 8.6 | 8 | 7.3‒11.3 | 8.95 | 1.37 | 15.3 |
| anterior pharyngostome rhabdion length | 15 | 8 | 13.0‒16.9 | 14.9 | 1.49 | 10.0 |
| posterior pharyngostome rhabdion length | 14 | 8 | 11.2‒16.0 | 13.5 | 1.40 | 10.3 |
| spicule length (arc) | 34 | 6 | 32.0‒39.0 | 35.2 | 2.86 | 8.12 |
| spicule ‘length (chord) | 28 | 6 | 29.0‒36.0 | 32.9 | 2.4 | 7.29 |
| number of supplements | 12 | 6 | 10‒13 | 11.5 | 1.05 | 9.13 |
| tail terminal cone length | 17 | 4 | 16.8–23.5 | 19.6 | 2.93 | 15.0 |
| Character | Holotype Male | Paratype Female 1 | Paratype Female 2 |
|---|---|---|---|
| Body length | 591 | 666 | 718 |
| a | 15.2 | 13.9 | 19.0 |
| b | 5.09 | 4.38 | 3.78 |
| c | 14.4 | tail not measurable | tail not measurable |
| c’ | 1.60 | tail not measurable | tail not measurable |
| V, % | - | 57.4 | 70.0 |
| body diameter at level of cephalic setae | 30.0 | 38.0 | 33.1 |
| body diameter at level of nerve ring | 30.5 | ? | 30.6 |
| body diameter at level of cardia | 35.0 | 41.5 | 33.8 |
| body diameter at level of midbody | 39.0 | 48.0 | 37.0 |
| body diameter at level of cloaca/anus | 25.6 | 30.0 | ? |
| inner labial setae length | 8.7 | 7.7 | 6.0 |
| latero-median outer labial setae length | 11.6 | not measurable | not measurable |
| lateral outer labial setae length | 10.0 | 5.5 | 6.3 |
| cephalic setae length | 8.8 | not measurable | not measurable |
| anterior cervical setae length | 10.0 | 12.0 | not measurable |
| anterior pharyngostome rhabdion length | 12.0 | 12.0 | 12.4 |
| posterior pharyngostome rhabdion length | 8.0 | 7.5 | 10.7 |
| spicules length (arc) | 38.0 | - | - |
| spicules length (chord) | 33.0 | - | - |
| gubernaculum length | 12.0 | - | - |
| tail terminal cone length | 16 | not measurable | not measurable |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tchesunov, A.; Jeong, R.; Lee, W. Two New Marine Free-Living Nematodes from Jeju Island Together with a Review of the Genus Gammanema Cobb 1920 (Nematoda, Chromadorida, Selachinematidae). Diversity 2020, 12, 19. https://doi.org/10.3390/d12010019
Tchesunov A, Jeong R, Lee W. Two New Marine Free-Living Nematodes from Jeju Island Together with a Review of the Genus Gammanema Cobb 1920 (Nematoda, Chromadorida, Selachinematidae). Diversity. 2020; 12(1):19. https://doi.org/10.3390/d12010019
Chicago/Turabian StyleTchesunov, Alexei, Raehyuk Jeong, and Wonchoel Lee. 2020. "Two New Marine Free-Living Nematodes from Jeju Island Together with a Review of the Genus Gammanema Cobb 1920 (Nematoda, Chromadorida, Selachinematidae)" Diversity 12, no. 1: 19. https://doi.org/10.3390/d12010019
APA StyleTchesunov, A., Jeong, R., & Lee, W. (2020). Two New Marine Free-Living Nematodes from Jeju Island Together with a Review of the Genus Gammanema Cobb 1920 (Nematoda, Chromadorida, Selachinematidae). Diversity, 12(1), 19. https://doi.org/10.3390/d12010019