The Effects of Habitat Heterogeneity at Distinct Spatial Scales on Hard-Bottom-Associated Communities


Abstract
:1. Introduction
2. Material and Methods
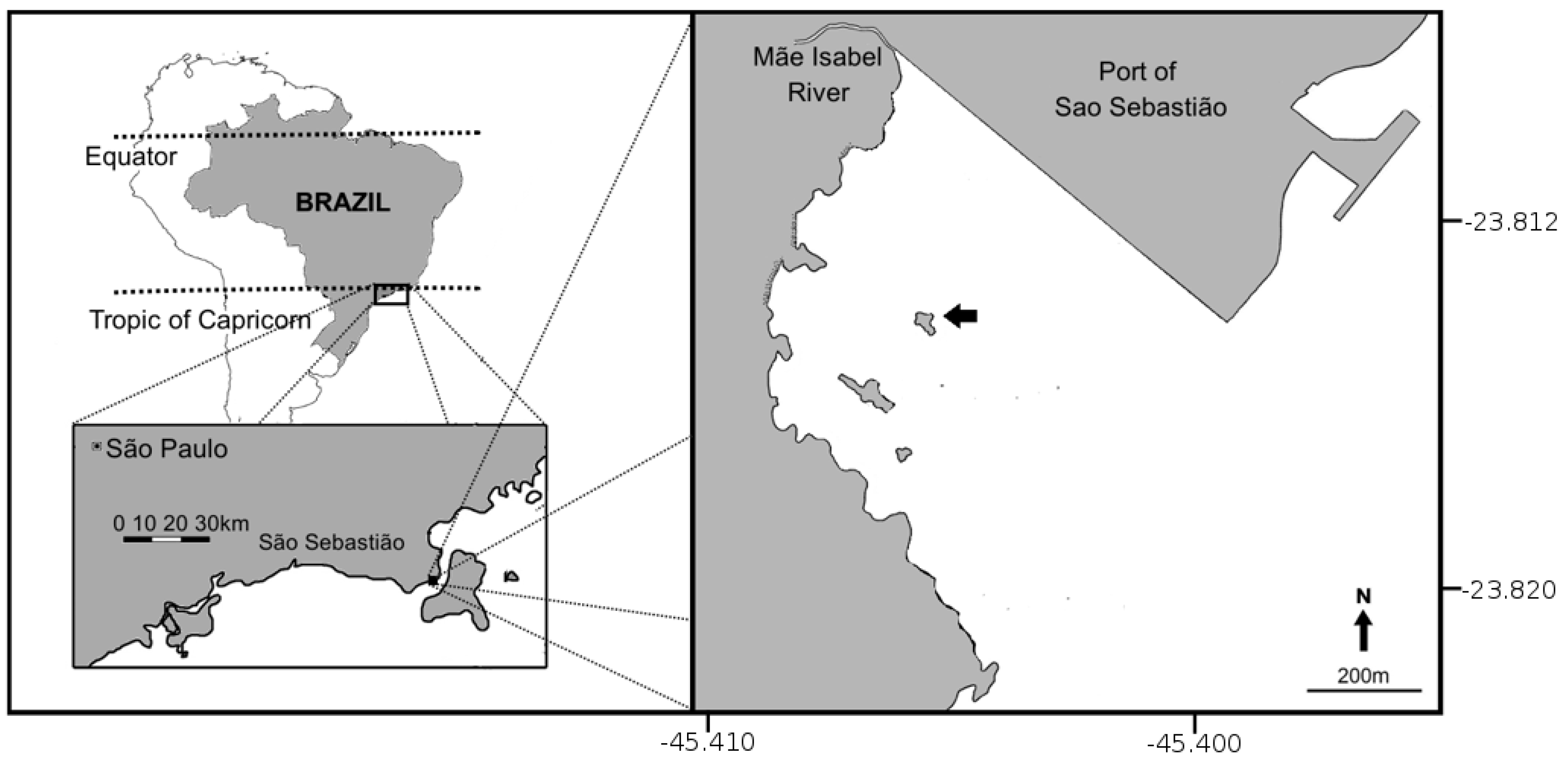
2.1. Study Area
2.2. Experimental Design
2.3. Sampling and Sample Processing
2.4. Statistical Analyses
3. Results
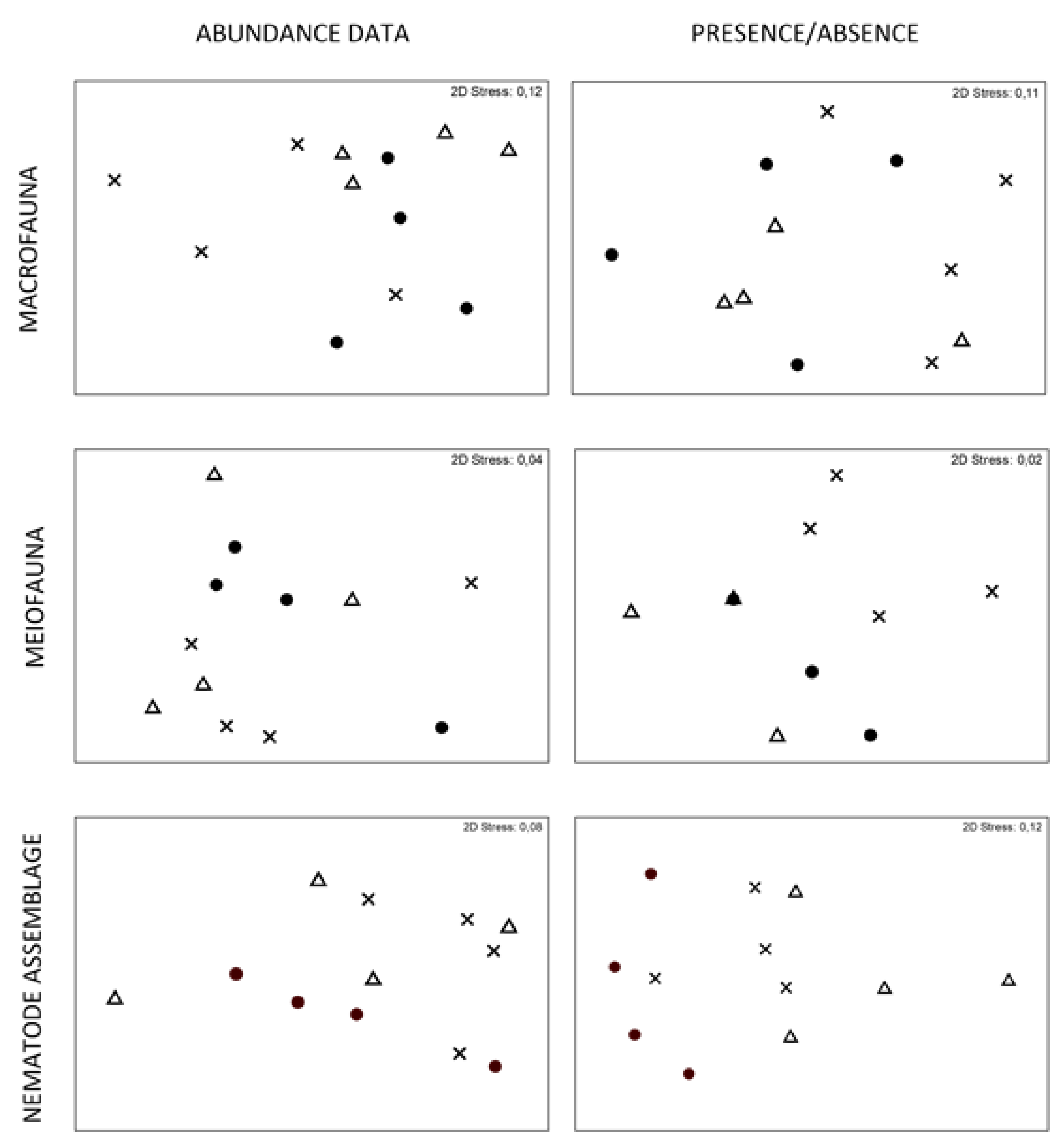
3.1. Different Biological Substrates Harbor Different Associated Communities
3.1.1. Meiofauna Community and Nematode Assemblage
3.1.2. Macrofauna Community
3.2. The Same Biological Substrate Growing in Different Habitats/Environments Harbor Different Associated Communities
3.2.1. Meiofauna Community and Nematodes Assemblage
3.2.2. Macrofauna Community
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hewitt, J.E.; Thrush, S.F.; Dayton, P.K.; Bonsdorff, E. The effect of spatial and temporal heterogeneity on the design and analysis of empirical studies of scale-dependent systems. Am. Nat. 2007, 169, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Moens, T.; Braeckman, U.; Derycke, S.; Fonseca, G.; Gallucci, F.; Gingold, R.; Guilini, K.; Ingels, J.; Leduc, D.; Vanaverbeke, J.; et al. Ecology of free-living marine nematodes. In Nematoda; DE GRUYTER: Berlin, Germany, 2013. [Google Scholar]
- Downes, B.J.; Lake, P.S.; Schreiber, E.S.G.; Glaister, A. Habitat Structure and Regulation of Local Species Diversity in a Stony, Upland Stream. Ecol. Monogr. 1998, 68, 237. [Google Scholar] [CrossRef]
- Chesson, P.; Rosenzweig, M. Behavior, Heterogeneity, and the Dynamics of Interacting Species. Ecology 1991, 72, 1187–1195. [Google Scholar] [CrossRef]
- Christie, H.; Norderhaug, K.; Fredriksen, S. Macrophytes as habitat for fauna. Mar. Ecol. Prog. Ser. 2009, 396, 221–233. [Google Scholar] [CrossRef] [Green Version]
- Kelaher, B. Influence of physical characteristics of coralline turf on associated macrofaunal assemblages. Mar. Ecol. Prog. Ser. 2002, 232, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Moreno, T.R.; da Rocha, R.M. Associated Fauna with Eudistoma Carolinense (Tunicata, Ascidiacea) Compared with other Biological Substrates with Different Architectures. J. Coast. Res. 2006, 39, 1695–1699. [Google Scholar]
- Chemello, R.; Milazzo, M. Effect of algal architecture on associated fauna: Some evidence from phytal molluscs. Mar. Biol. 2002, 140, 981–990. [Google Scholar]
- Coull, B.C.; Wells, J.B.J. Refuges from Fish Predation: Experiments with Phytal Meiofauna from the New Zealand Rocky Intertidal. Ecology 1983, 64, 1599–1609. [Google Scholar] [CrossRef]
- Duffy, J.E.; Hay, M.E. Strong Impacts of Grazing Amphipods on the Organization of a Benthic Community. Ecol. Monogr. 2000, 70, 237. [Google Scholar] [CrossRef]
- Gibbons, M.J. Impact of predation by juvenile Clinus superciliosus on phytal meiofauna: Are fish important as predators? Mar. Ecol. Prog. Ser. 1988, 45, 13–22. [Google Scholar] [CrossRef]
- Hicks, G.R.F. Structure of phytal harpacticoid copepod assemblages and the influence of habitat complexity and turbidity. J. Exp. Mar. Biol. Ecol. 1980, 44, 157–192. [Google Scholar] [CrossRef]
- Martin-Smith, K.M. Abundance of mobile epifauna: The role of habitat complexity and predation by fishes. J. Exp. Mar. Biol. Ecol. 1993, 174, 243–260. [Google Scholar] [CrossRef]
- Thiébaut, E.; Cabioch, L.; Dauvin, J.-C.; Retiere, C.; Gentil, F. Spatio-Temporal Persistence of the Abra Alba-Pectinaria Koreni Muddy-Fine Sand Community of the Eastern Bay of Seine. J. Mar. Biol. Assoc. UK 1997, 77, 1165–1185. [Google Scholar] [CrossRef]
- Van Hoey, G.; Vincx, M.; Degraer, S. Temporal variability in the Abra alba community determined by global and local events. J. Sea Res. 2007, 58, 144–155. [Google Scholar] [CrossRef]
- Schratzberger, M.; Maxwell, T.A.D.; Warr, K.; Ellis, J.R.; Rogers, S.I. Spatial variability of infaunal nematode and polychaete assemblages in two muddy subtidal habitats. Mar. Biol. 2008, 153, 621–642. [Google Scholar] [CrossRef]
- Lebreton, B.; Richard, P.; Galois, R.; Radenac, G.; Brahmia, A.; Colli, G.; Grouazel, M.; André, C.; Guillou, G.; Blanchard, G.F. Food sources used by sediment meiofauna in an intertidal Zostera noltii seagrass bed: A seasonal stable isotope study. Mar. Biol. 2012, 159, 1537–1550. [Google Scholar] [CrossRef] [Green Version]
- Moens, T.; Verbeeck, L.; de Maeyer, A.; Swings, J.; Vincx, M. Selective attraction of marine bacterivorous nematodes to their bacterial food. Mar. Ecol. Prog. Ser. 1999, 176, 165–178. [Google Scholar] [CrossRef]
- Gallucci, F.; Steyaert, M.; Moens, T. Can field distributions of marine predacious nematodes be explained by sediment constraints on their foraging success? Mar. Ecol. Prog. Ser. 2005, 304, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Vafeiadou, A.-M.; Materatski, P.; Adão, H.; de Troch, M.; Moens, T. Resource utilization and trophic position of nematodes and harpacticoid copepods in and adjacent to Zostera noltii beds. Biogeosciences 2014, 11, 4001–4014. [Google Scholar] [CrossRef] [Green Version]
- Species Size Distributions in Marine Benthic Communities on JSTOR. Available online: https://www.jstor.org/stable/4217200?seq=1#metadata_info_tab_contents (accessed on 29 November 2019).
- Eckman, J.E. Hydrodynamic processes affecting benthic recruitment1. Limnol. Oceanogr. 1983, 28, 241–257. [Google Scholar] [CrossRef]
- Amaral, A.C.Z.; Ciotti, A.M.; Fonseca, G. Biodiversity and functioning of a subtropical coastal ecosystem: Subsidies for integrated management. Ocean Coast. Manag. 2018, 164, 1–3. [Google Scholar] [CrossRef]
- Pardal-Souza, A.L.; Dias, G.M.; Jenkins, S.R.; Ciotti, Á.M.; Christofoletti, R.A. Shading impacts by coastal infrastructure on biological communities from subtropical rocky shores. J. Appl. Ecol. 2017, 54, 826–835. [Google Scholar] [CrossRef] [Green Version]
- Vieira, E.A.; Filgueiras, H.R.; Bueno, M.; Leite, F.P.P.; Dias, G.M. Algas co-ocorrentes e morfologicamente distintas suportam uma diversa fauna associada na zona entremarés na baía do araçá, Brasil. Biota Neotrop. 2018, 18. [Google Scholar] [CrossRef] [Green Version]
- Danovaro, R.; Fraschetti, S. Meiofaunal vertical zonation on hard-bottoms: Comparison with soft-bottom meiofauna. Mar. Ecol. Prog. Ser. 2002, 230, 159–169. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Austral Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Taniguchi, H.; Tokeshi, M. Effects of habitat complexity on benthic assemblages in a variable environment. Freshw. Biol. 2004, 49, 1164–1178. [Google Scholar] [CrossRef]
- Gratwicke, B.; Speight, M.R. The relationship between fish species richness, abundance and habitat complexity in a range of shallow tropical marine habitats. J. Fish Biol. 2005, 66, 650–667. [Google Scholar] [CrossRef]
- Sueiro, M.C.; Bortolus, A.; Schwindt, E. Habitat complexity and community composition: Relationships between different ecosystem engineers and the associated macroinvertebrate assemblages. Helgol. Mar. Res. 2011, 65, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Willis, S.C.; Winemiller, K.O.; Lopez-Fernandez, H. Habitat structural complexity and morphological diversity of fish assemblages in a Neotropical floodplain river. Oecologia 2005, 142, 284–295. [Google Scholar] [CrossRef]
- St. Pierre, J.I.; Kovalenko, K.E. Effect of habitat complexity attributes on species richness. Ecosphere 2014, 5, art22. [Google Scholar] [CrossRef]
- Hauser, A.; Attrill, M.J.; Cotton, P.A. Effects of habitat complexity on the diversity and abundance of macrofauna colonising artificial kelp holdfasts. Mar. Ecol. Prog. Ser. 2006, 325, 93–100. [Google Scholar] [CrossRef]
- Öster, M.; Cousins, S.A.O.; Eriksson, O. Size and heterogeneity rather than landscape context determine plant species richness in semi-natural grasslands. J. Veg. Sci. 2007, 18, 859–868. [Google Scholar] [CrossRef]
- Passarelli, C.; Olivier, F.; Paterson, D.; Hubas, C. Impacts of biogenic structures on benthic assemblages: Microbes, meiofauna, macrofauna and related ecosystem functions. Mar. Ecol. Prog. Ser. 2012, 465, 85–97. [Google Scholar] [CrossRef] [Green Version]
- Reise, K. High abundance of small zoobenthos around biogenic structures in tidal sediments of the Wadden Sea. Helgoländer Meeresunters. 1981, 34, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Hasemann, C.; Soltwedel, T. Small-Scale Heterogeneity in Deep-Sea Nematode Communities around Biogenic Structures. PLoS ONE 2011, 6, e29152. [Google Scholar] [CrossRef] [Green Version]
- Bell, S.S.; Watzin, M.C.; Coull, B.C. Biogenic structure and its effect on the spatial heterogeneity of meiofauna in a salt marsh. J. Exp. Mar. Biol. Ecol. 1978, 35, 99–107. [Google Scholar] [CrossRef]
- Veiga, P.; Sousa-Pinto, I.; Rubal, M. Meiofaunal assemblages associated with native and non-indigenous macroalgae. Cont. Shelf Res. 2016, 123, 1–8. [Google Scholar] [CrossRef]
- Trotter, D.B.; Webster, J.M. Feeding preferences and seasonality of free-living marine nematodes inhabiting the kelp Macrocystis integrifolia. Mar. Ecol. Prog. Ser. 1984, 14, 151–157. [Google Scholar] [CrossRef]
- Frame, K.; Hunt, G.; Roy, K. Intertidal meiofaunal biodiversity with respect to different algal habitats: A test using phytal ostracodes from Southern California. Hydrobiologia 2007, 586, 331–342. [Google Scholar] [CrossRef] [Green Version]
- Hicks, G.R.F. Observations on substrate preference of marine phytal harpacticoids (copepoda). Hydrobiologia 1977, 56, 7–9. [Google Scholar] [CrossRef]
- Gibbons, M.J. The impact of sediment accumulations, relative habitat complexity and elevation on rocky shore meiofauna. J. Exp. Mar. Biol. Ecol. 1988, 122, 225–241. [Google Scholar] [CrossRef]
- Russo, A.R. Role of habitat complexity in mediating predation by the gray damselfish Abudefduf sordidus on epiphytal amphipods. Mar. Ecol. Prog. Ser. 1987, 36, 101–105. [Google Scholar] [CrossRef]
- Watt, C.A.; Scrosati, R.A. Bioengineer effects on understory species richness, diversity, and composition change along an environmental stress gradient: Experimental and mensurative evidence. Estuar. Coast. Shelf Sci. 2013, 123, 10–18. [Google Scholar] [CrossRef]
- Umanzor, S.; Ladah, L.; Calderon-Aguilera, L.E.; Zertuche-González, J.A. Intertidal macroalgae influence macroinvertebrate distribution across stress scenarios. Mar. Ecol. Prog. Ser. 2017, 584, 67–77. [Google Scholar] [CrossRef]
- De Ronde, C.E.J.; Stoffers, P.; Garbe-Schönberg, D.; Christenson, B.W.; Jones, B.; Manconi, R.; Browne, P.R.L.; Hissmann, K.; Botz, R.; Davy, B.W.; et al. Discovery of active hydrothermal venting in Lake Taupo, New Zealand. J. Volcanol. Geotherm. Res. 2002, 115, 257–275. [Google Scholar] [CrossRef]
- Rota, E.; Manconi, R. Taxonomy and Ecology of Sponge-Associate Marionina spp. (Clitellata: Enchytraeidae) from the Horomatangi Geothermal System of Lake Taupo, New Zealand. Int. Rev. Hydrobiol. 2004, 89, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Ávila, E.; Ortega-Bastida, A.L. Influence of habitat and host morphology on macrofaunal assemblages associated with the sponge Halichondria melanadocia in an estuarine system of the southern Gulf of Mexico. Mar. Ecol. 2015, 36, 1345–1353. [Google Scholar] [CrossRef]
- Bueno, M.; Dias, G.M.; Leite, F.P.P. The importance of shore height and host identity for amphipod assemblages. Mar. Biol. Res. 2017, 13, 870–877. [Google Scholar] [CrossRef]
- Thiel, M.; Ullrich, N. Hard rock versus soft bottom: The fauna associated with intertidal mussel beds on hard bottoms along the coast of Chile, and considerations on the functional role of mussel beds. Helgol. Mar. Res. 2002, 56, 21–30. [Google Scholar] [CrossRef]
- Gagarin, V.G.; Thanh, N.V. Four new species of monhysterids (Nematoda: Monhysterida) from mangroves of the Mekong river estuaries of Vietnam. TAP CHI SINH HOC 2014, 30, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Brustolin, M.C.; Nagelkerken, I.; Fonseca, G. Large-scale distribution patterns of mangrove nematodes: A global meta-analysis. Ecol. Evol. 2018, 8, 4734–4742. [Google Scholar] [CrossRef] [PubMed]
- Malerba, M.E.; White, C.R.; Marshall, D.J. The outsized trophic footprint of marine urbanization. Front. Ecol. Environ. 2019, 17, 400–406. [Google Scholar] [CrossRef]
- Schratzberger, M.; Ingels, J. Meiofauna matters: The roles of meiofauna in benthic ecosystems. J. Exp. Mar. Biol. Ecol. 2018, 502, 12–25. [Google Scholar] [CrossRef]
- Belley, R.; Snelgrove, P.V.R. Relative contributions of biodiversity and environment to benthic ecosystem functioning. Front. Mar. Sci. 2016, 3, 242. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| df | F | p | |
|---|---|---|---|
| Meiofauna density | 2 | 0.079 | 0.925 |
| Nematode density | 2 | 0.513 | 0.610 |
| Number of meiofaunal taxa | 2 | 0.600 | 0.569 |
| Number of nematode species | 2 | 2.212 | 0.149 |
| Nematode diversity (H’) | 2 | 2.101 | 0.162 |
| Macrofauna density | 2 | 3.055 | 0.097 |
| Number of macrofaunal taxa | 2 | 0.694 | 0.524 |
| Macrofauna diversity (H’) | 2 | 2.486 | 0.138 |
| Abundance Data (Log (x + 1)) | Presence/Absence Data | |||||
|---|---|---|---|---|---|---|
| Source | df | Pseudo-F | p (perm) | df | Pseudo-F | p (perm) |
| Macrofauna | ||||||
| Treatment | 2 | 1.174 | 0.323 | 2 | 0.969 | 0.441 |
| Residual | 9 | 9 | ||||
| Meiofauna | ||||||
| Treatment | 2 | 1.642 | 0.115 | 2 | 5.021 | 0.003 |
| Residual | 9 | 9 | ||||
| Nematodes | ||||||
| Treatment | 2 | 1.282 | 0.212 | 2 | 2.6989 | 0.003 |
| Residual | 9 | 9 | ||||
| df | F | p | |
|---|---|---|---|
| Meiofauna density | 1 | 0.751 | 0.419 |
| Nematode density | 1 | 0.400 | 0.550 |
| Number of meiofaunal taxa | 1 | 0.297 | 0.606 |
| Meiofauna diversity (H’) | 1 | 0.128 | 0.732 |
| Number of nematode species | 1 | 3.261 | 0.121 |
| Nematode diversity (H’) | 1 | 11.97 | 0.013 |
| Macrofauna density | 1 | 0.235 | 0.645 |
| Number of macrofaunal taxa | 1 | 25.00 | 0.002 |
| Macrofauna diversity (H’) | 1 | 53.50 | <0.001 |
| Source | df | Pseudo-F | p (perm) |
|---|---|---|---|
| Macrofauna | |||
| Treatment | 1 | 5.2506 | 0.026 |
| Residual | 6 | ||
| Meiofauna | |||
| Treatment | 1 | 2.3293 | 0.125 |
| Residual | 6 | ||
| Nematodes | |||
| Treatment | 1 | 3.7215 | 0.032 |
| Residual | 6 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallucci, F.; A. Christofoletti, R.; Fonseca, G.; M. Dias, G. The Effects of Habitat Heterogeneity at Distinct Spatial Scales on Hard-Bottom-Associated Communities. Diversity 2020, 12, 39. https://doi.org/10.3390/d12010039
Gallucci F, A. Christofoletti R, Fonseca G, M. Dias G. The Effects of Habitat Heterogeneity at Distinct Spatial Scales on Hard-Bottom-Associated Communities. Diversity. 2020; 12(1):39. https://doi.org/10.3390/d12010039
Chicago/Turabian StyleGallucci, Fabiane, Ronaldo A. Christofoletti, Gustavo Fonseca, and Gustavo M. Dias. 2020. "The Effects of Habitat Heterogeneity at Distinct Spatial Scales on Hard-Bottom-Associated Communities" Diversity 12, no. 1: 39. https://doi.org/10.3390/d12010039
APA StyleGallucci, F., A. Christofoletti, R., Fonseca, G., & M. Dias, G. (2020). The Effects of Habitat Heterogeneity at Distinct Spatial Scales on Hard-Bottom-Associated Communities. Diversity, 12(1), 39. https://doi.org/10.3390/d12010039