Competitive Interactions of Flowering Rush (Butomus umbellatus L.) Cytotypes in Submersed and Emergent Experimental Aquatic Plant Communities


Abstract
:1. Introduction
2. Materials and Methods
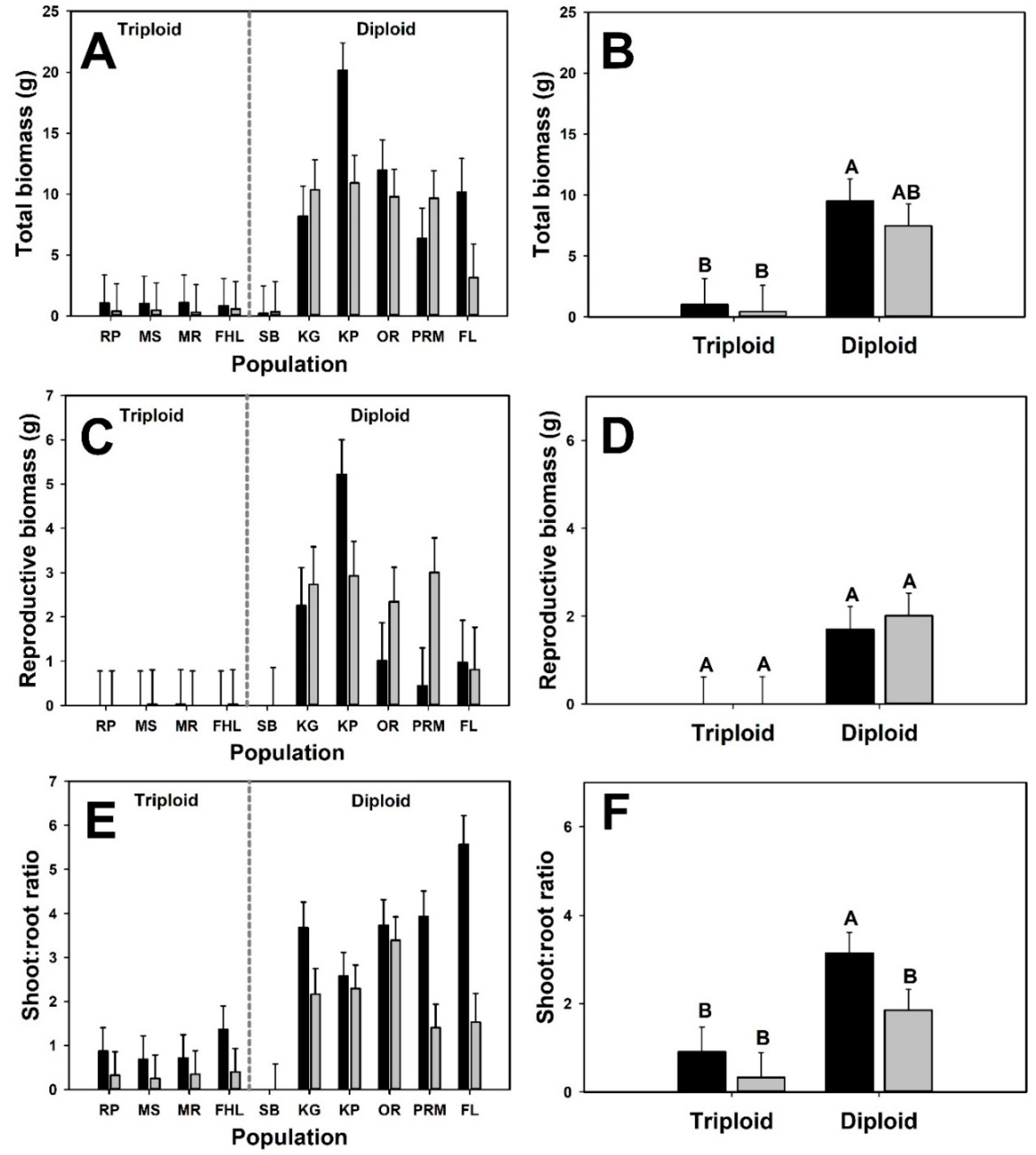
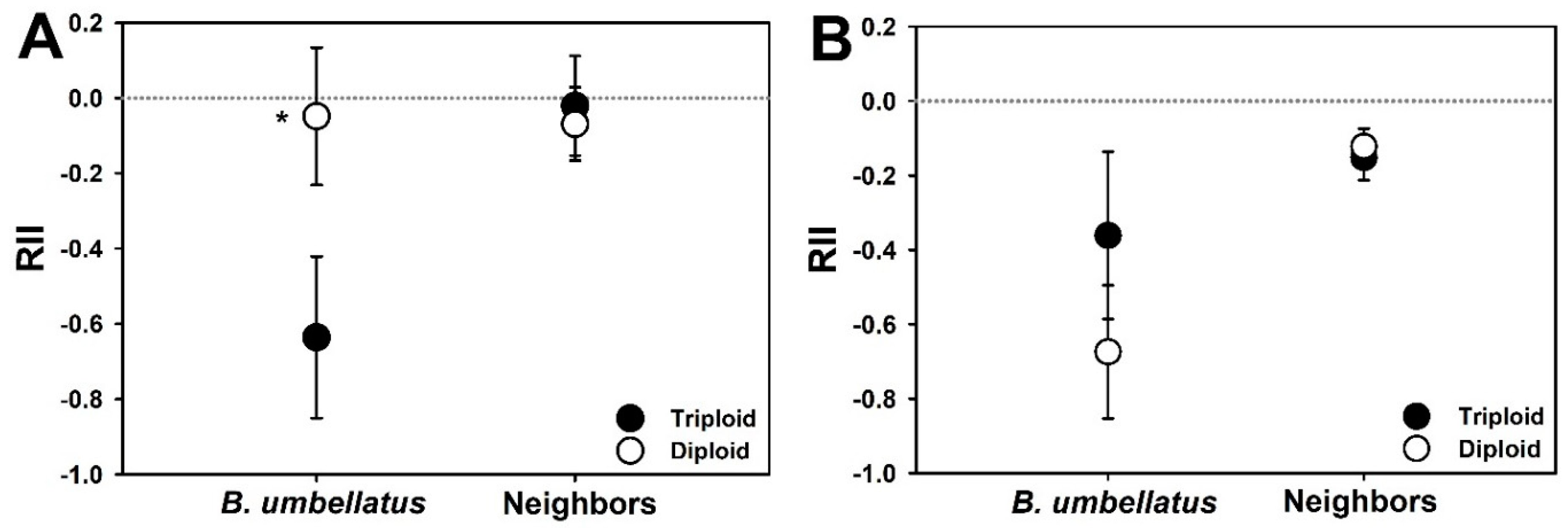
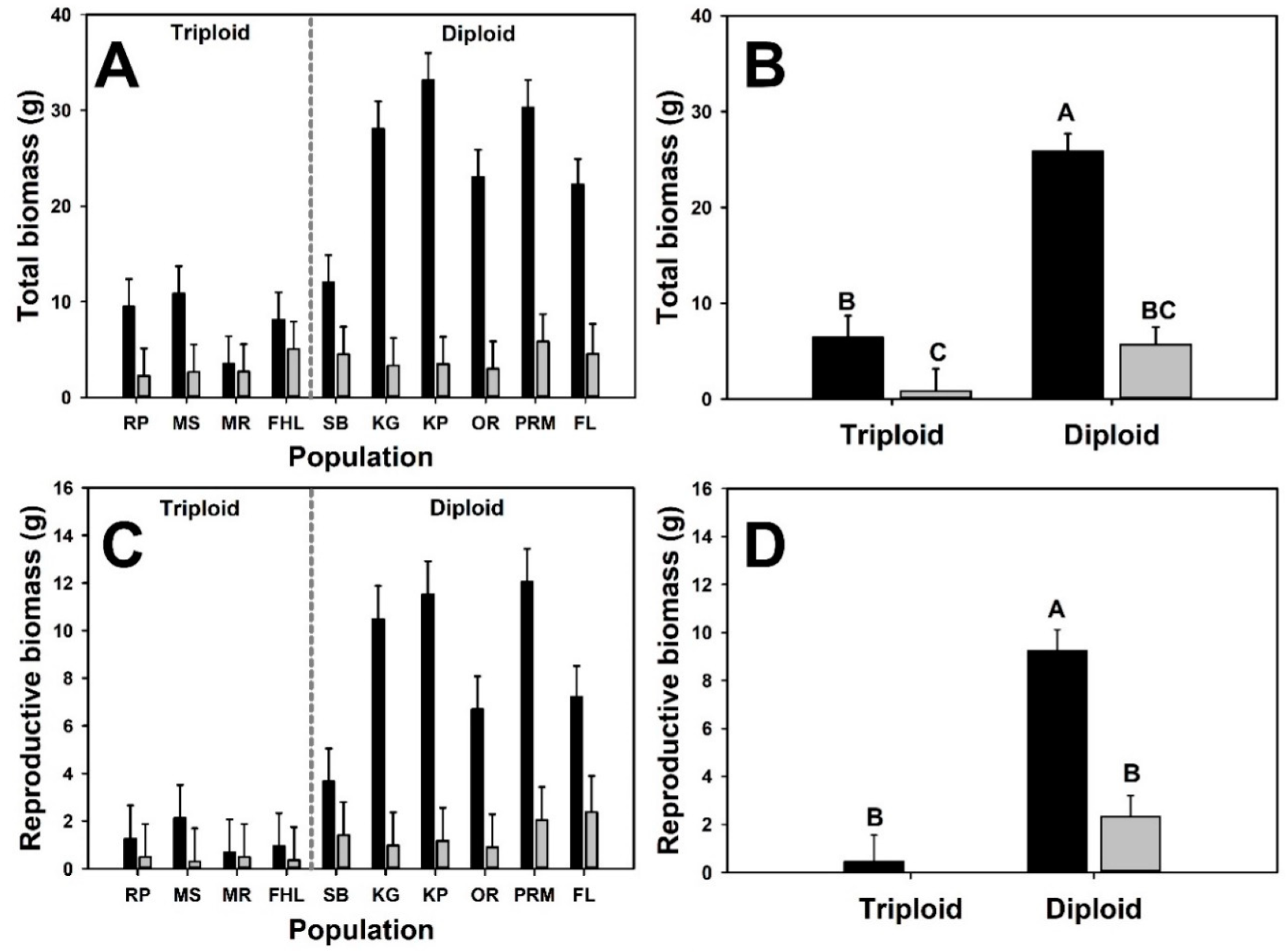
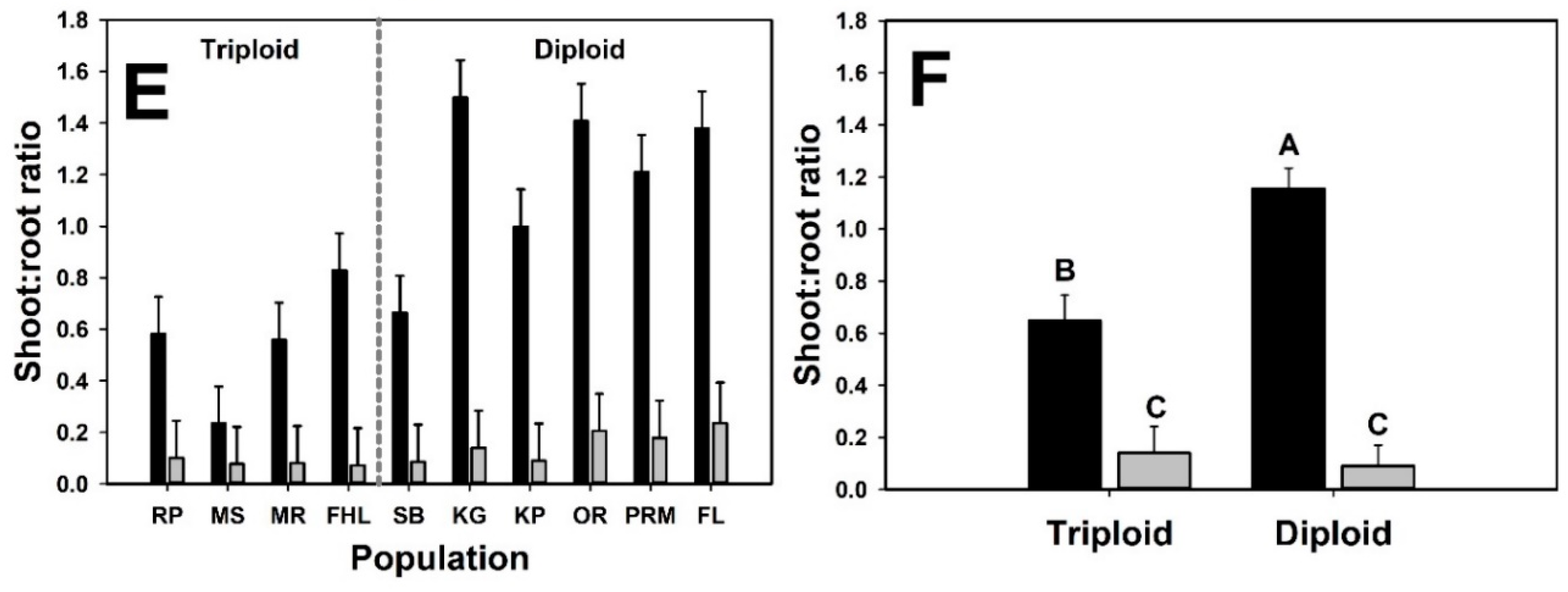
3. Results
4. Discussion
Funding
Acknowledgments
Conflicts of Interest
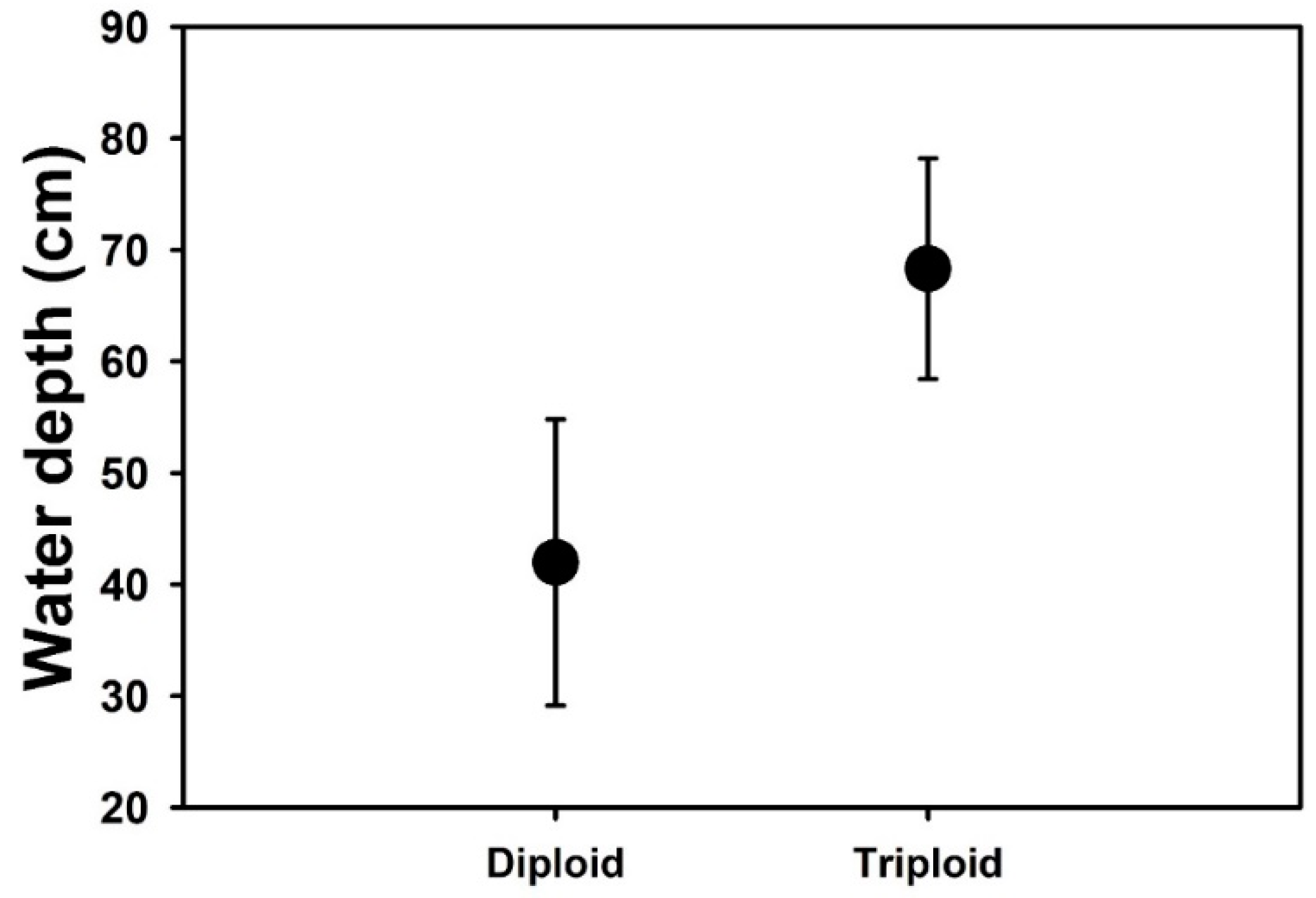
Appendix A. Field Observations on Water Depth of B. umbellatus Populations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Year | Cytotype | Water Depth (cm) |
|---|---|---|---|
| Aberdeen Golf Course Canal, ID, USA | 2014 | Triploid | 100 |
| Columbia River @ Kennewick, WA, USA | 2014 | Triploid | 200 |
| Flathead Lake, MT, USA | 2014 | Triploid | 100 |
| Lake Pend Oreille, ID, USA | 2014 | Triploid | 125 |
| Lake Spokane, WA, USA | 2014 | Triploid | 160 |
| Pend Oreille River, WA, USA | 2014 | Triploid | 100 |
| Rose Pond, ID, USA | 2014 | Triploid | 100 |
| Yakima River, Horn Rapids Park, WA, USA | 2014 | Triploid | 100 |
| Forest Lake, MN, USA | 2015 | Diploid | 115 |
| Kildeer Pond 33, OH, USA | 2015 | Diploid | 100 |
| Olentangy River, OH, USA | 2015 | Diploid | 0 |
| Point Rosa Marsh, MI, USA | 2015 | Diploid | 0 |
| Sterling State Park, MI, USA | 2015 | Diploid | 0 |
| Bertram Lake, WI, USA | 2015 | Triploid | 0 |
| Lake Kawaguesaga, WI, USA | 2015 | Triploid | 100 |
| Lanes Lake, MI, USA | 2015 | Triploid | 0 |
| MS River near Galena, IL, USA | 2015 | Triploid | 0 |
| Oconto Falls, WI, USA | 2015 | Triploid | 100 |
| Village Park, Fremont, WI, USA | 2015 | Triploid | 0 |
| Cayuga Lake, NY, USA | 2016 | Diploid | 10 |
| East Bay Wildlife Management Area, NY, USA | 2016 | Diploid | 0 |
| Forest Lake, MN, USA | 2016 | Diploid | 100 |
| Kildeer Pond 33, OH, USA | 2016 | Diploid | 0 |
| Oswegatchie River, VT, USA | 2016 | Diploid | 10 |
| Point Rosa Marsh, MI, USA | 2016 | Diploid | 10 |
| Shelburne Bay, VT, USA | 2016 | Diploid | 0 |
| Springbrook Pond, IL, USA | 2016 | Diploid | 0 |
| Sterling State Park, MI, USA | 2016 | Diploid | 0 |
| Three Mile Bay, NY, USA | 2016 | Diploid | 25 |
| Unity Island, NY, USA | 2016 | Diploid | 30 |
| Aberdeen Golf Course Canal, ID, USA | 2016 | Triploid | 40 |
| Columbia River @ Kennewick, WA, USA | 2016 | Triploid | 125 |
| Flathead Lake, MT, USA | 2016 | Triploid | 70 |
| Lake Pend Oreille, ID, USA | 2016 | Triploid | 80 |
| Missisquoi River, VT, USA | 2016 | Triploid | 0 |
| Pend Oreille River, WA, USA | 2016 | Triploid | 80 |
| Rose Pond, ID, USA | 2016 | Triploid | 70 |
| Sabattus Creek, ME, USA | 2016 | Triploid | 20 |
| Yakima River @ Prosser, WA, USA | 2016 | Triploid | 15 |

References
- Gopal, B. Water Hyacinth; Elsevier Science Publishers: Amsterdam, The Netherlands, 1987. [Google Scholar]
- Wainger, L.A.; Harms, N.E.; Magen, C.; Liang, N.; Nesslage, G.M.; McMurray, A.M.; Cofrancesco, A.F. Evidence-based economic analysis demonstrates that ecosystem service benefits of water hyacinth management greatly exceed research and control costs. PeerJ 2018, 6, e4824. [Google Scholar] [CrossRef] [PubMed]
- Villamagna, A.M.; Murphy, B.R. Ecological and socio-economic impacts of invasive water hyacinth (Eichhornia crassipes): A review. Freshw. Biol. 2010, 55, 282–298. [Google Scholar] [CrossRef]
- Pejchar, L.; Mooney, H.A. Invasive species, ecosystem services and human well-being. Trends Ecol. Evol. 2009, 24, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.R. Effects of invasive, non-indigenous plant species on ecosystem processes: Lessons from Florida. Ecol. Appl. 1998, 8, 975–989. [Google Scholar] [CrossRef]
- Evans, J.M. Ecosystem implications of invasive aquatic plants and aquatic plant control in florida springs. In Summary and Synthesis of Available Literature on the Effects of Nutrients on Springs Organisms and Systems; Florida Department of Environmental Protection: Tallahassee, FL, USA, 2008; pp. 249–270. [Google Scholar]
- Gallardo, B.; Clavero, M.; Sánchez, M.I.; Vilà, M. Global ecological impacts of invasive species in aquatic ecosystems. Glob. Chang. Biol. 2016, 22, 151–163. [Google Scholar] [CrossRef]
- Zhang, C.; Boyle, K.J. The effect of an aquatic invasive species (Eurasian watermilfoil) on lakefront property values. Ecol. Econ. 2010, 70, 394–404. [Google Scholar] [CrossRef]
- Keller, R.P.; Masoodi, A.; Shackleton, R.T. The impact of invasive aquatic plants on ecosystem services and human well-being in Wular Lake, India. Reg. Environ. Chang. 2018, 18, 847–857. [Google Scholar] [CrossRef]
- Wu, H.; Ding, J.Q. Global Change Sharpens the Double-Edged Sword Effect of Aquatic Alien Plants in China and Beyond. Front. Plant Sci. 2019, 10, 787. [Google Scholar] [CrossRef] [Green Version]
- Dukes, J.S.; Mooney, H.A. Disruption of ecosystem processes in western North America by invasive species. Rev. Chil. Hist. Nat. 2004, 77, 411–437. [Google Scholar] [CrossRef]
- Charles, H.; Dukes, J.S. Impacts of Invasive Species on Ecosystem Services. In Biological Invasions; Nentwig, W., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 217–237. [Google Scholar]
- Fournier, A.; Penone, C.; Pennino, M.G.; Courchamp, F. Predicting future invaders and future invasions. Proc. Natl. Acad. Sci. USA 2019, 116, 7905–7910. [Google Scholar] [CrossRef] [Green Version]
- Divíšek, J.; Chytrý, M.; Beckage, B.; Gotelli, N.J.; Lososová, Z.; Pyšek, P.; Richardson, D.M.; Molofsky, J. Similarity of introduced plant species to native ones facilitates naturalization, but differences enhance invasion success. Nat. Commun. 2018, 9, 4631. [Google Scholar] [CrossRef]
- Kolar, C.S.; Lodge, D.M. Progress in invasion biology: Predicting invaders. Trends Ecol. Evol. 2001, 16, 199–204. [Google Scholar] [CrossRef]
- Mack, R.N. Predicting the identity and fate of plant invaders: Emergent and emerging approaches. Biol. Conserv. 1996, 78, 107–121. [Google Scholar] [CrossRef]
- Keller, R.P.; Lodge, D.M.; Finnoff, D.C. Risk assessment for invasive species produces net bioeconomic benefits. Proc. Natl. Acad. Sci. USA 2007, 104, 203–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodge, D.M.; Williams, S.; MacIsaac, H.J.; Hayes, K.R.; Leung, B.; Reichard, S.; Mack, R.N.; Moyle, P.B.; Smith, M.; Andow, D.A.; et al. Biological invasions: Recommendations for U.S. policy and management. Ecol. Appl. 2006, 16, 2035–2054. [Google Scholar] [CrossRef] [Green Version]
- Simberloff, D.; Parker, I.M.; Windle, P.N. Introduced species policy, management, and future research needs. Front. Ecol. Environ. 2005, 3, 12–20. [Google Scholar] [CrossRef]
- Mack, R.N.; Simberloff, D.; Mark Lonsdale, W.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Humphries, S.E. An Integrated Approach to the Ecology and Management of Plant Invasions. Conserv. Biol. 1995, 9, 761–770. [Google Scholar] [CrossRef] [Green Version]
- Zanden, M.J.V.; Olden, J.D. A management framework for preventing the secondary spread of aquatic invasive species. Can. J. Fish. Aquat. Sci. 2008, 65, 1512–1522. [Google Scholar] [CrossRef]
- Zanden, M.J.V.; Hansen, G.J.; Higgins, S.N.; Kornis, M.S. A pound of prevention, plus a pound of cure: Early detection and eradication of invasive species in the Laurentian Great Lakes. J. Great Lakes Res. 2010, 36, 199–205. [Google Scholar] [CrossRef]
- Byers, J.E.; Noonburg, E.G. Scale dependent effects of biotic resistance to biological invasion. Ecology 2003, 84, 1428–1433. [Google Scholar] [CrossRef]
- Levine, J.M.; Adler, P.B.; Yelenik, S.G. A meta-analysis of biotic resistance to exotic plant invasions. Ecol. Lett. 2004, 7, 975–989. [Google Scholar] [CrossRef] [Green Version]
- Parker, J.D.; Hay, M.E. Biotic resistance to plant invasions? Native herbivores prefer non-native plants. Ecol. Lett. 2005, 8, 959–967. [Google Scholar] [CrossRef]
- Byun, C.; Lee, E.J. Ecological application of biotic resistance to control the invasion of an invasive plant, Ageratina altissima. Ecol. Evol. 2017, 7, 2181–2192. [Google Scholar] [CrossRef]
- Byun, C.; de Blois, S.; Brisson, J. Plant functional group identity and diversity determine biotic resistance to invasion by an exotic grass. J. Ecol. 2013, 101, 128–139. [Google Scholar] [CrossRef] [Green Version]
- Maron, J.L.; Vilà, M. When do herbivores affect plant invasion? Evidence for the natural enemies and biotic resistance hypotheses. Oikos 2001, 95, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Richardson, D.M.; Pyšek, P. Naturalization of introduced plants: Ecological drivers of biogeographical patterns. New Phytol. 2012, 196, 383–396. [Google Scholar] [CrossRef]
- Barbosa, P.; Hines, J.; Kaplan, I.; Martinson, H.; Szczepaniec, A.; Szendrei, Z. Associational Resistance and Associational Susceptibility: Having Right or Wrong Neighbors. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Andersen, C.P.; Louda, S.M. Abundance of and Floral Herbivory on Exotic Bull Thistle Versus Native Tall Thistle in Western Tallgrass Prairie; University of Nebraska at Kearney: Kearney, NE, USA, 2006. [Google Scholar]
- Yannelli, F.; Koch, C.; Jeschke, J.; Kollmann, J. Limiting similarity and Darwin’s naturalization hypothesis: Understanding the drivers of biotic resistance against invasive plant species. Oecologia 2017, 183, 775–784. [Google Scholar] [CrossRef]
- Kennedy, T.A.; Naeem, S.; Howe, K.M.; Knops, J.M.H.; Tilman, D.; Reich, P. Biodiversity as a barrier to ecological invasion. Nature 2002, 417, 636–638. [Google Scholar] [CrossRef]
- Thiele, J.; Isermann, M.; Otte, A.; Kollmann, J. Competitive displacement or biotic resistance? Disentangling relationships between community diversity and invasion success of tall herbs and shrubs. J. Veg. Sci. 2010, 21, 213–220. [Google Scholar] [CrossRef]
- Britton-Simmons, K.H. Functional group diversity, resource preemption and the genesis of invasion resistance in a community of marine algae. Oikos 2006, 113, 395–401. [Google Scholar] [CrossRef]
- Funk, J.L.; Cleland, E.E.; Suding, K.N.; Zavaleta, E.S. Restoration through reassembly: Plant traits and invasion resistance. Trends Ecol. Evol. 2008, 23, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Middleton, E.L.; Bever, J.D.; Schultz, P.A. The Effect of Restoration Methods on the Quality of the Restoration and Resistance to Invasion by Exotics. Restor. Ecol. 2010, 18, 181–187. [Google Scholar] [CrossRef]
- Iii, B.V.I.; Potter, K.M.; Guo, Q.; Jo, I.; Oswalt, C.M.; Fei, S. Environmental harshness drives spatial heterogeneity in biotic resistance. NeoBiota 2018, 40, 87–105. [Google Scholar]
- Iannone, B.V.; Oswalt, C.M.; Liebhold, A.M.; Guo, Q.; Potter, K.M.; Nunez-Mir, G.C.; Oswalt, S.N.; Pijanowski, B.C.; Fei, S. Region-specific patterns and drivers of macroscale forest plant invasions. Divers. Distrib. 2015, 21, 1181–1192. [Google Scholar] [CrossRef] [Green Version]
- Araújo, M.B.; Rozenfeld, A. The geographic scaling of biotic interactions. Ecography 2014, 37, 406–415. [Google Scholar] [CrossRef] [Green Version]
- Souza, L.; Bunn, W.A.; Simberloff, D.; Lawton, R.M.; Sanders, N.J. Biotic and abiotic influences on native and exotic richness relationship across spatial scales: Favourable environments for native species are highly invasible. Funct. Ecol. 2011, 25, 1106–1112. [Google Scholar] [CrossRef]
- Richards, C.L.; Pennings, S.C.; Donovan, L.A. Habitat range and phenotypic variation in salt marsh plants. Plant Ecol. 2005, 176, 263–273. [Google Scholar] [CrossRef]
- Davidson, A.M.; Jennions, M.D.; Nicotra, A.B. Do invasive species show higher phenotypic plasticity than native species and, if so, is it adaptive? A meta-analysis. Ecol. Lett. 2011, 14, 419–431. [Google Scholar] [CrossRef]
- Turner, K.G.; Fréville, H.; Rieseberg, L.H. Adaptive plasticity and niche expansion in an invasive thistle. Ecol. Evol. 2015, 5, 3183–3197. [Google Scholar] [CrossRef] [PubMed]
- Vretare, V.; Weisner, S.E.; Strand, J.A.; Granéli, W. Phenotypic plasticity in Phragmites australis as a functional response to water depth. Aquat. Bot. 2001, 69, 127–145. [Google Scholar] [CrossRef]
- Pendergrass, A.G.; Knutti, R.; Lehner, F.; Deser, C.; Sanderson, B.M. Precipitation variability increases in a warmer climate. Sci. Rep. 2017, 7, 17966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, S.Q.; Wei, F.; Zeng, L.Y.; Li, X.K.; Tang, S.C.; Zhong, Y.; Geng, Y.P. Multiple introductions are responsible for the disjunct distributions of invasive Parthenium hysterophorus in China: Evidence from nuclear and chloroplast DNA. Weed Res. 2009, 49, 373–380. [Google Scholar] [CrossRef]
- Kelager, A.; Pedersen, J.S.; Bruun, H.H. Multiple introductions and no loss of genetic diversity: Invasion history of Japanese Rose, Rosa rugosa, in Europe. Biol. Invasions 2013, 15, 1125–1141. [Google Scholar] [CrossRef]
- Zhu, X.C.; Gopurenko, D.; Serrano, M.; Spencer, M.A.; Pieterse, P.J.; Skoneczny, D.; Lepschi, B.J.; Reigosa, M.J.; Gurr, G.M.; Callaway, R.M.; et al. Genetic evidence for plural introduction pathways of the invasive weed Paterson’s curse (Echium plantagineum L.) to southern Australia. PLoS ONE 2019, 14, e0222696. [Google Scholar] [CrossRef] [Green Version]
- Ward, S.M.; Gaskin, J.F.; Wilson, L.M. Ecological Genetics of Plant Invasion: What Do We Know? Invasive Plant Sci. Manag. 2008, 1, 98–109. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.A.; Overholt, W.A.; Cuda, J.P.; Hughes, C.R. Chloroplast and microsatellite DNA diversities reveal the introduction history of Brazilian peppertree (Schinus terebinthifolius) in Florida. Mol. Ecol. 2005, 14, 3643–3656. [Google Scholar] [CrossRef]
- Larue, E.A.; Zuellig, M.P.; Netherland, M.D.; Heilman, M.A.; Thum, R.A. Hybrid watermilfoil lineages are more invasive and less sensitive to a commonly used herbicide than their exotic parent (Eurasian watermilfoil). Evol. Appl. 2013, 6, 462–471. [Google Scholar] [CrossRef]
- Thum, R.A. Genetic variation and aquatic plant management: Key concepts and practical implications. J. Aquat. Plant Manag. 2018, 56, 101–106. [Google Scholar]
- Gaskin, J.F.; Bon, M.C.; Cock, M.J.; Cristofaro, M.; De Biase, A.; De Clerck-Floate, R.; Ellison, C.A.; Hinz, H.L.; Hufbauer, R.A.; Julien, M.H.; et al. Applying molecular-based approaches to classical biological control of weeds. Biol. Control 2011, 58, 1–21. [Google Scholar] [CrossRef]
- Williams, W.I.; Friedman, J.M.; Gaskin, J.F.; Norton, A.P. Hybridization of an invasive shrub affects tolerance and resistance to defoliation by a biological control agent. Evol. Appl. 2014, 7, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Hroudová, Z.; Zákravský, P. Ecology of two cytotypes of Butomus umbellatus II. Reproduction, growth and biomass production. Folia Geobot. Phytotaxon. 1993, 28, 413–424. [Google Scholar] [CrossRef]
- Anderson, L.C.; Zeis, C.D.; Alam, S.F. Phytogeography and Possible Origins of Butomus in North America. Bull. Torrey Bot. Club 1974, 101, 292–296. [Google Scholar] [CrossRef]
- Harms, N.; Shearer, J.; Cronin, J.T.; Gaskin, J.F. Geographic and genetic variation in susceptibility of Butomus umbellatus to foliar fungal pathogens. Biol. Invasions 2019, 1–14. [Google Scholar] [CrossRef]
- Cao, L.; Berent, L.; Fusaro, A. Butomus umbellatus L. Available online: https://nas.er.usgs.gov/queries/greatLakes/FactSheet.aspx?SpeciesID=1100&Potential=N&Type=0&HUCNumber= (accessed on 21 November 2019).
- Parkinson, H.; Mangold, J.; Dupuis, V.; Rice, P. Biology, Ecology and Mangament of Flowering Rush (Butomus umbellatus); Montana State University: Bozeman, MT, USA, 2010; pp. 1–12. [Google Scholar]
- Lesica, P.; Lavin, M.; Stickney, P.F. Manual of Montana Vascular Plants; BRIT Press: Ft. Worth, TX, USA, 2012. [Google Scholar]
- Jacobs, J.; Mangold, J.; Parkinson, H.; Dupuis, V.; Rice, P. Ecology and management of flowering rush (Butomus umbellatus L.); Invasive Species Technical Note No. MT-33; United States Department of Agriculture: Washington, DC, USA; Natural Resources Conservation Service: Washington, DC, USA, 2011.
- Roach, D.A.; Wulff, R.D. Maternal Effects in Plants. Annu. Rev. Ecol. Syst. 1987, 18, 209–235. [Google Scholar] [CrossRef]
- Leger, E.A.; Espeland, E.K. PERSPECTIVE: Coevolution between native and invasive plant competitors: Implications for invasive species management. Evol. Appl. 2010, 3, 169–178. [Google Scholar] [CrossRef]
- Smart, R.M.; Barko, J.W. Laboratory culture of submersed freshwater macrophytes on natural sediments. Aquat. Bot. 1985, 21, 251–263. [Google Scholar] [CrossRef]
- Bose, R.C.; Clatworthy, W.H.; Shrikhande, S.S. Tables of Partially Balanced Designs with Two Associate Classes; North Carolina State University: Raleigh, NC, USA, 1954. [Google Scholar]
- Armas, C.; Ordiales, R.; Pugnaire, F.I. Measuring Plant Interactions: A New Comparative Index. Ecology 2004, 85, 2682–2686. [Google Scholar] [CrossRef] [Green Version]
- Cavieres, L.A.; Sanhueza, A.K.; Torres-Mellado, G.; Casanova-Katny, A. Competition between native Antarctic vascular plants and invasive Poa annua changes with temperature and soil nitrogen availability. Biol. Invasions 2018, 20, 1597–1610. [Google Scholar] [CrossRef] [Green Version]
- Madsen, J.D.; Sartain, B.; Turnage, G.; Marko, M. Management of flowering rush in the Detroit Lakes, Minnesota. J. Aquat. Plant Manag. 2016, 54, 61–67. [Google Scholar]
- Turnage, G.; Madsen, J.D.; Wersal, R.M.; Byrd, J.D. Simulated mechanical control of flowering rush (Butomus umbellatus) under mesocosm conditions. Invasive Plant Sci. Manag. 2019, 12, 120–123. [Google Scholar] [CrossRef]
- Bhattarai, G.P.; Meyerson, L.A.; Anderson, J.; Cummings, D.; Allen, W.J.; Cronin, J.T. Biogeography of a plant invasion: Genetic variation and plasticity in latitudinal clines for traits related to herbivory. Ecol. Monogr. 2017, 87, 57–75. [Google Scholar] [CrossRef]
- Bhattarai, G.P.; Meyerson, L.A.; Cronin, J.T. Geographic variation in apparent competition between native and invasive Phragmites australis. Ecology 2017, 98, 349–358. [Google Scholar] [CrossRef] [Green Version]
- Gillard, M.; Grewell, B.J.; Futrell, C.J.; Deleu, C.; Thiébaut, G. Germination and Seedling Growth of Water Primroses: A Cross Experiment between Two Invaded Ranges with Contrasting Climates. Front. Plant Sci. 2017, 8, 1677. [Google Scholar] [CrossRef] [PubMed]
- Grewell, B.J.; Thomason, M.J.S.; Futrell, C.J.; Iannucci, M.; Drenovsky, R.E. Trait responses of invasive aquatic macrophyte congeners: Colonizing diploid outperforms polyploid. AoB Plants 2016, 8. [Google Scholar] [CrossRef] [Green Version]
- Castillo, J.M.; Grewell, B.J.; Pickart, A.; Bortolus, A.; Peña, C.; Figueroa, E.; Sytsma, M. Phenotypic plasticity of invasive Spartina densiflora (Poaceae) along a broad latitudinal gradient on the Pacific Coast of North America. Am. J. Bot. 2014, 101, 448–458. [Google Scholar] [CrossRef] [Green Version]
- Grewell, B.J.; Castillo, J.M.; Thomason, M.J.S.; Drenovsky, R.E. Phenotypic plasticity and population differentiation in response to salinity in the invasive cordgrass Spartina densiflora. Biol. Invasions 2016, 18, 2175–2187. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.J.; Zhou, F.; Pan, X.Y.; van Kleunen, M.; Liu, M.; Li, B. Evolution of increased intraspecific competitive ability following introduction: The importance of relatedness among genotypes. J. Ecol. 2019, 107, 387–395. [Google Scholar] [CrossRef]
- Liu, L.; Dong, B.C.; Alpert, P.; Yu, F.H. Effects of soil substrate heterogeneity and moisture on interspecific competition between Alternanthera philoxeroidesand four native species. J. Plant Ecol. 2016, 10, 528–537. [Google Scholar]
- Liu, M.; Zhou, F.; Pan, X.Y.; Zhang, Z.J.; Traw, M.B.; Li, B. Specificity of herbivore-induced responses in an invasive species, Alternanthera philoxeroides (alligator weed). Ecol. Evol. 2018, 8, 59–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aerts, R.; Boot, R.G.A.; Van Der Aart, P.J.M. The relation between above- and belowground biomass allocation patterns and competitive ability. Oecologia 1991, 87, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Cahill, J.J.F.; Casper, B.B. Investigating the relationship between neighbor root biomass and belowground competition: Field evidence for symmetric competition belowground. Oikos 2000, 90, 311–320. [Google Scholar] [CrossRef] [Green Version]
- Rudak, A.; Wódkiewicz, M.; Znój, A.; Chwedorzewska, K.J.; Galera, H. Plastic biomass allocation as a trait increasing the invasiveness of annual bluegrass (Poa annua L.) in Antarctica. Polar Biol. 2018, 42, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef]
- Ren, G.Q.; Li, Q.; Li, Y.; Li, J.; Adomako, M.O.; Dai, Z.C.; Li, G.L.; Wan, L.Y.; Zhang, B.; Zou, C.B.; et al. The enhancement of root biomass increases the competitiveness of an invasive plant against a co-occurring native plant under elevated nitrogen deposition. Flora 2019, 261, 151486. [Google Scholar] [CrossRef]
- Harms, N.E.; Cronin, J.T.; Gaskin, J.F. Increased ploidy in the invasive range of Butomus umbellatus L. is not associated with higher phenotypic plasticity to N and P. in review. 2020. [Google Scholar]
- Sterck, F.J.; Clark, D.B.; Clark, D.A.; Bongers, F. Light fluctuations, crown traits, and response delays for tree saplings in a Costa Rican lowland rain forest. J. Trop. Ecol. 1999, 15, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Bargeron, C.T.; Moorhead, D.J. EDDMapS—Early detection and distribution mapping system for the southeast exotic pest plant council. Wildland Weeds 2007, 10, 4–8. [Google Scholar]




| Population | Latitude | Longitude | Cytotype | Genotype * |
|---|---|---|---|---|
| Kildeer Pond, OH, USA | 40.710 | −83.369 | Diploid | G4 |
| Springbrook Pond, IL, USA | 41.730 | −88.205 | Diploid | G3 |
| Point Rosa Marsh, MI, USA | 42.576 | −82.805 | Diploid | G4 |
| Oswegatchie River, NY, USA | 44.690 | −75.495 | Diploid | G4 |
| Kenduskeag River, ME, USA | 44.818 | −68.793 | Diploid | G4 |
| Forest Lake, MN, USA | 45.272 | −92.937 | Diploid | G5 |
| Mississippi River, IL, USA | 42.343 | −90.409 | Triploid | G1 |
| Rose Pond, ID, USA | 43.247 | −112.315 | Triploid | G1 |
| Mississquoi River, VT, USA | 44.951 | −73.162 | Triploid | G1 |
| Flathead Lake, MT, USA | 47.697 | −114.071 | Triploid | G1 |
| Factor | Num df | Den df | F | p | |
|---|---|---|---|---|---|
| Submersed | Total biomass | ||||
| Cytotype | 1 | 13.4 | 8.74 | 0.011 | |
| Neighbor presence | 1 | 98 | 1.49 | 0.225 | |
| Cytotype × Neighbor presence | 1 | 98.8 | 0.77 | 0.383 | |
| Initial mass | 1 | 103 | 1.21 | 0.275 | |
| Reproductive biomass | |||||
| Cytotype | 1 | 15.7 | 5.39 | 0.034 | |
| Neighbor presence | 1 | 98.2 | 0.18 | 0.670 | |
| Cytotype × Neighbor presence | 1 | 99.4 | 0.09 | 0.771 | |
| Initial mass | 1 | 90.6 | 0.38 | 0.539 | |
| Shoot:root Ratio | |||||
| Cytotype | 1 | 11.8 | 6.7 | 0.024 | |
| Neighbor presence | 1 | 97.3 | 12.81 | <0.001 | |
| Cytotype × Neighbor presence | 1 | 98.1 | 2.2 | 0.141 | |
| Initial mass | 1 | 105 | 0.59 | 0.445 | |
| Emergent | Total biomass | ||||
| Cytotype | 1 | 10.4 | 20.93 | <0.001 | |
| Neighbor presence | 1 | 107 | 94.28 | <0.001 | |
| Cytotype × Neighbor presence | 1 | 107 | 34.64 | <0.001 | |
| Initial mass | 1 | 109 | 7.44 | 0.007 | |
| Reproductive biomass | |||||
| Cytotype | 1 | 11.5 | 23.81 | <0.001 | |
| Neighbor presence | 1 | 107 | 39.67 | <0.001 | |
| Cytotype × Neighbor presence | 1 | 107 | 23.6 | <0.001 | |
| Initial mass | 1 | 96.1 | 3.93 | 0.05 | |
| Shoot:root Ratio | |||||
| Cytotype | 1 | 11.9 | 4.86 | 0.048 | |
| Neighbor presence | 1 | 106 | 124.8 | <0.001 | |
| Cytotype × Neighbor presence | 1 | 106 | 18.73 | <0.001 | |
| Initial mass | 1 | 94.1 | 4.78 | 0.031 |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harms, N.E. Competitive Interactions of Flowering Rush (Butomus umbellatus L.) Cytotypes in Submersed and Emergent Experimental Aquatic Plant Communities. Diversity 2020, 12, 40. https://doi.org/10.3390/d12010040
Harms NE. Competitive Interactions of Flowering Rush (Butomus umbellatus L.) Cytotypes in Submersed and Emergent Experimental Aquatic Plant Communities. Diversity. 2020; 12(1):40. https://doi.org/10.3390/d12010040
Chicago/Turabian StyleHarms, Nathan E. 2020. "Competitive Interactions of Flowering Rush (Butomus umbellatus L.) Cytotypes in Submersed and Emergent Experimental Aquatic Plant Communities" Diversity 12, no. 1: 40. https://doi.org/10.3390/d12010040
APA StyleHarms, N. E. (2020). Competitive Interactions of Flowering Rush (Butomus umbellatus L.) Cytotypes in Submersed and Emergent Experimental Aquatic Plant Communities. Diversity, 12(1), 40. https://doi.org/10.3390/d12010040