Niche Complementarity and Resistance to Grazing Promote the Invasion Success of Sargassum horneri in North America

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study System
2.2. Competition
2.3. Complementarity
2.4. Herbivory
2.5. Software Used for Statistical Analysis
3. Results
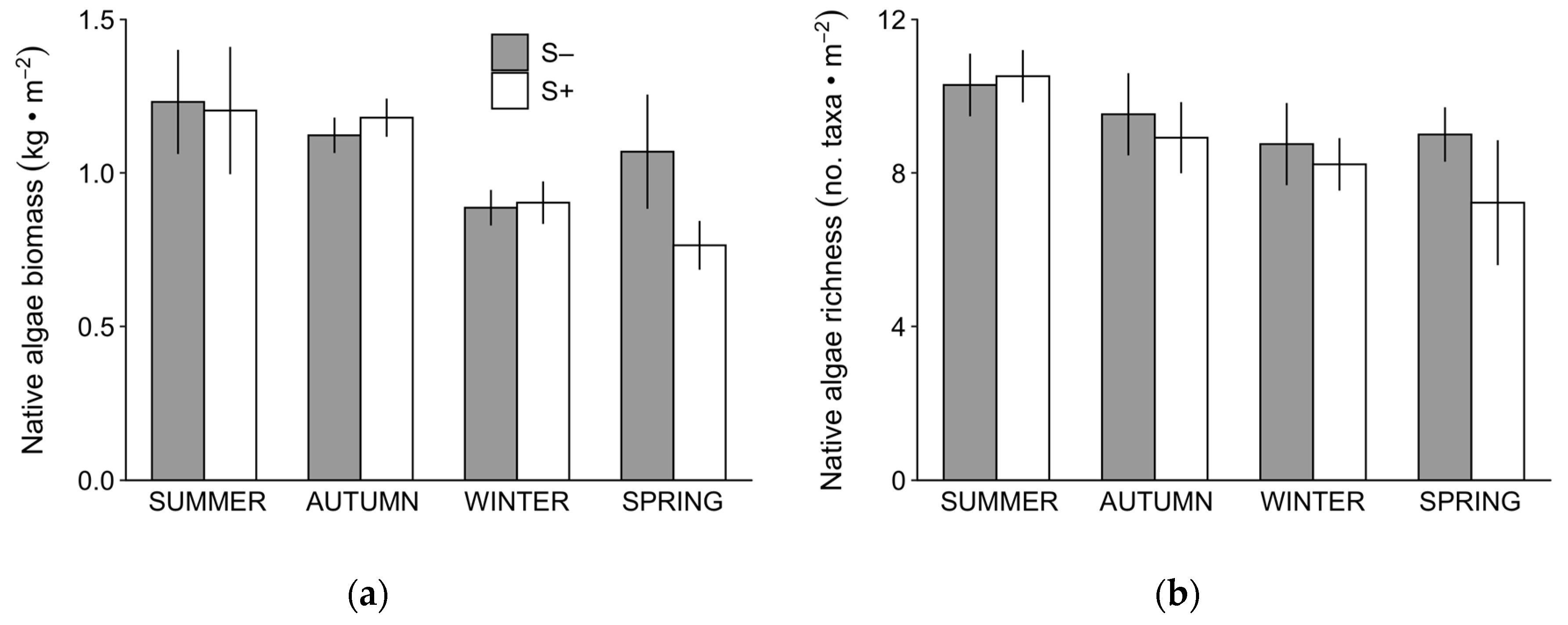
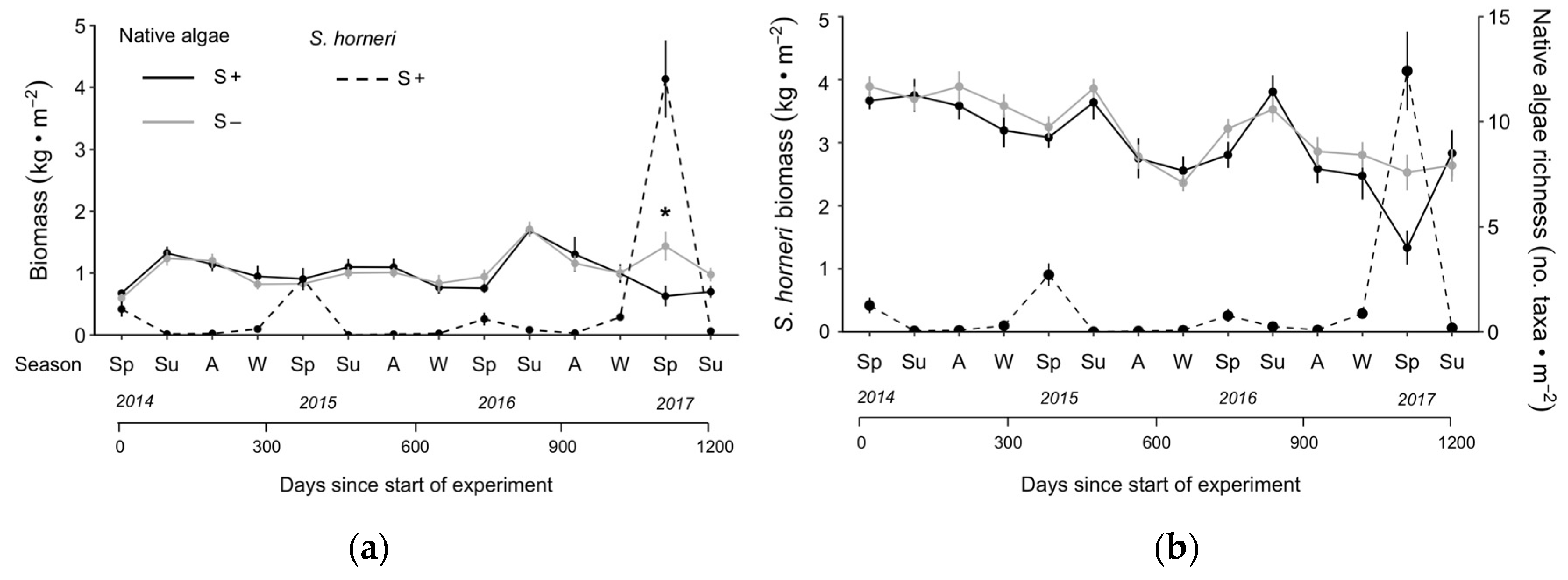
3.1. Competition
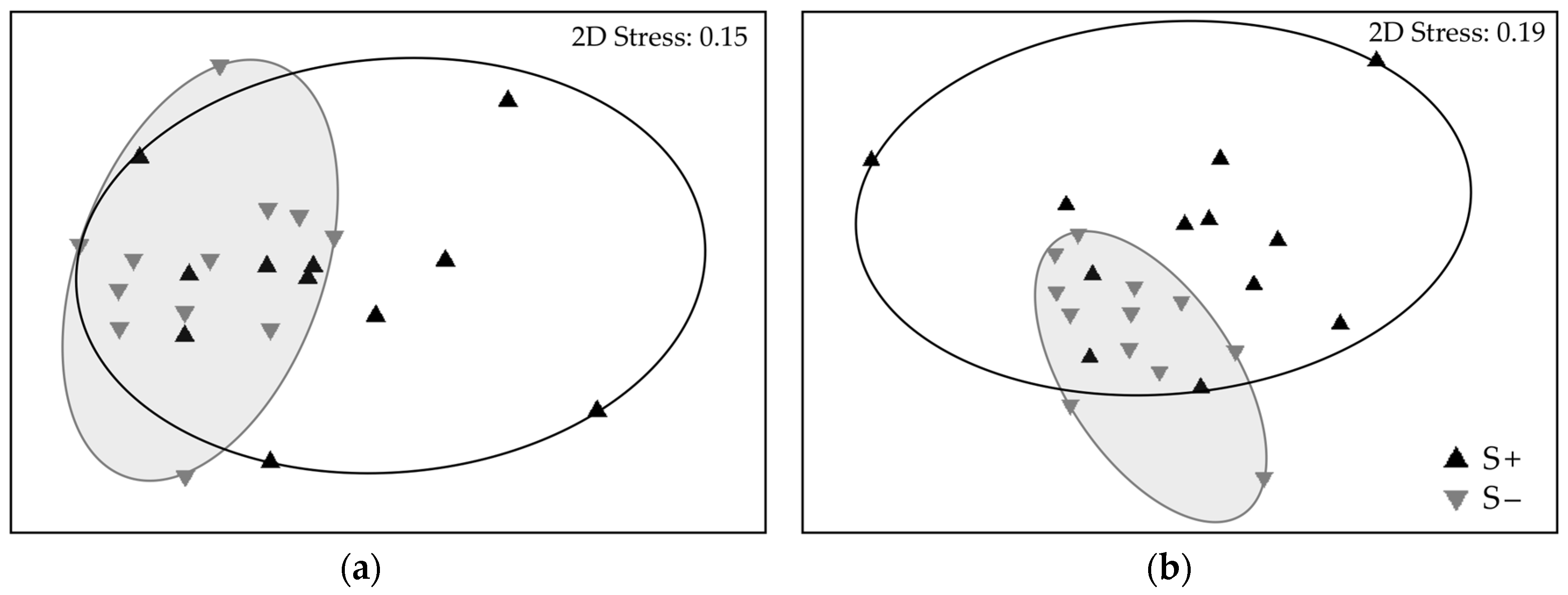
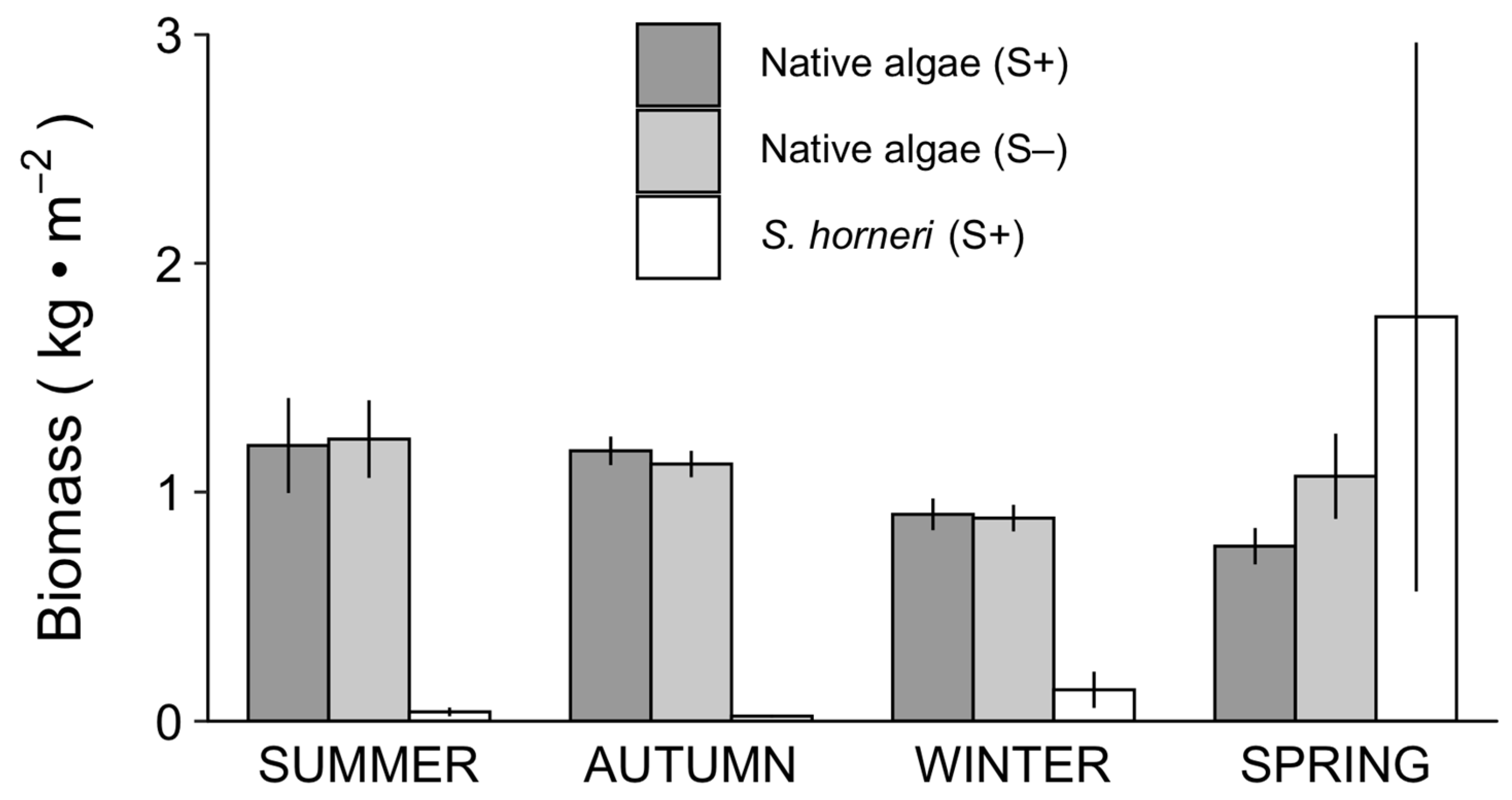
3.2. Complementarity
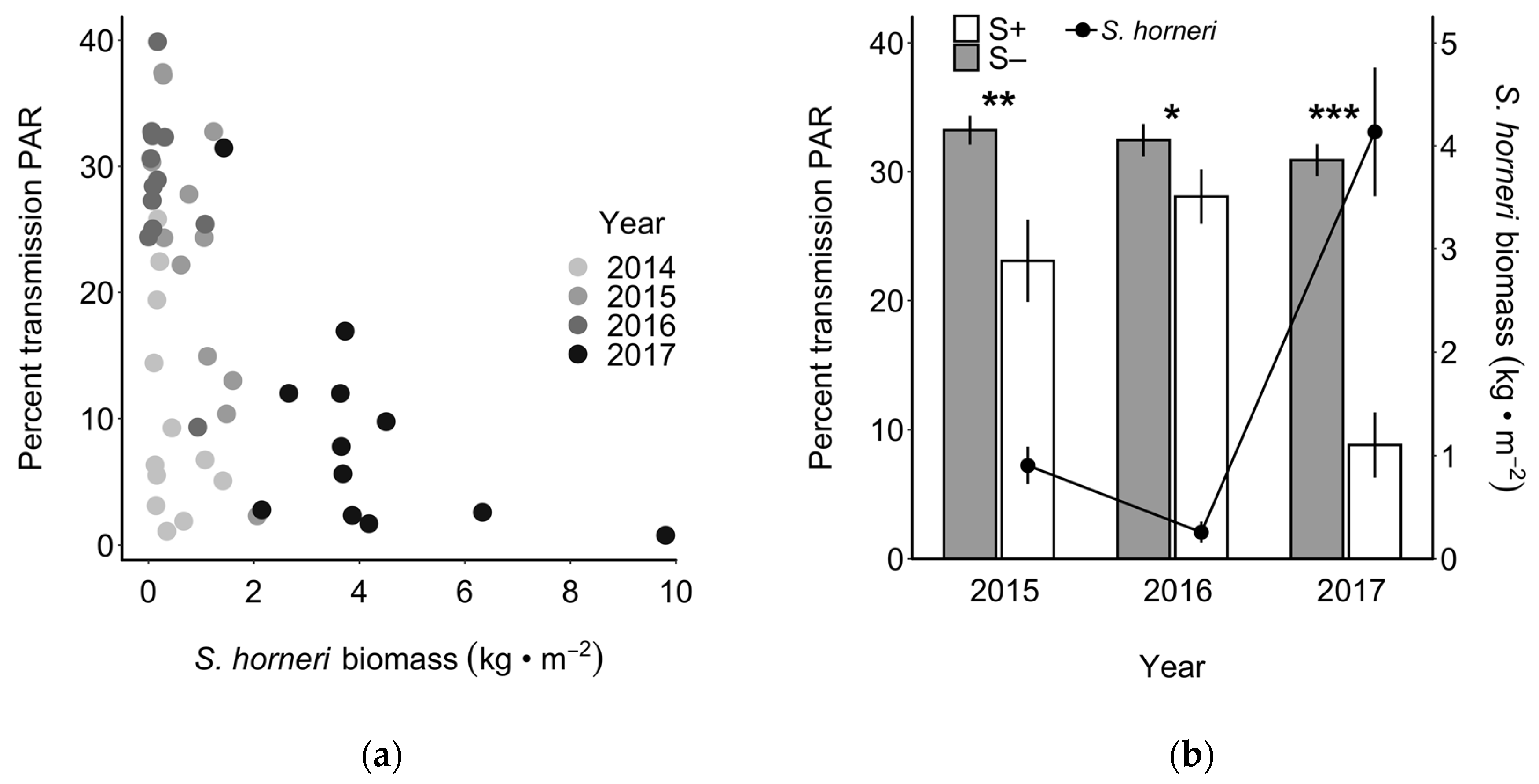
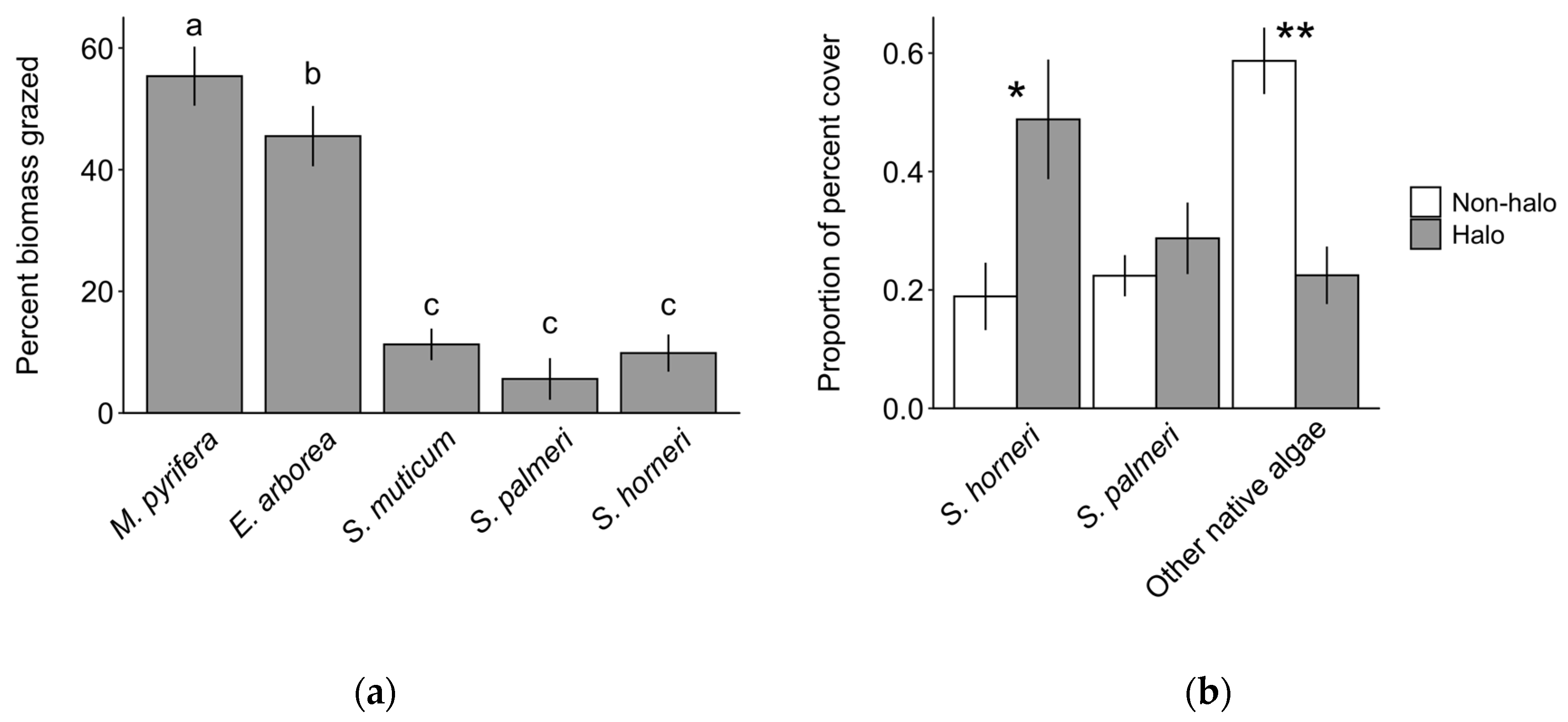
3.3. Herbivory
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cohen, A.N.; Carlton, J.T. Accelerating invasion rate in a highly invaded estuary. Science 1998, 279, 555–558. [Google Scholar] [CrossRef] [Green Version]
- Stachowicz, J.J.; Terwin, J.R.; Whitlatch, R.B.; Osman, R.W. Linking climate change and biological invasions: Ocean warming facilitates nonindigenous species invasions. Proc. Natl. Acad. Sci. USA 2002, 99, 15497–15500. [Google Scholar] [CrossRef] [Green Version]
- Molnar, J.L.; Gamboa, R.L.; Revenga, C.; Spalding, M.D. Assessing the global threat of invasive species to marine biodiversity. Front. Ecol. Environ. 2008, 6, 485–492. [Google Scholar] [CrossRef]
- Sorte, C.J.; Williams, S.L.; Zerebecki, R.A. Ocean warming increases threat of invasive species in a marine fouling community. Ecology 2010, 91, 2198–2204. [Google Scholar] [CrossRef]
- Seebens, H.; Gastner, M.T.; Blasius, B. The risk of marine bioinvasion caused by global shipping. Ecol. Lett. 2013, 16, 782–790. [Google Scholar] [CrossRef] [PubMed]
- Papacostas, K.J.; Rielly-Carroll, E.W.; Georgian, S.E.; Long, D.J.; Princiotta, S.D.; Quattrini, A.M.; Reuter, K.E.; Freestone, A.L. Biological mechanisms of marine invasions. Mar. Ecol. Prog. Ser. 2017, 565, 251–268. [Google Scholar] [CrossRef] [Green Version]
- Caselle, J.C.; Davis, K.; Marks, L.M. Marine management affects the invasion success of a non-native species in a temperate reef system in California, USA. Ecol. Lett. 2018, 21, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Elton, C.S. The Ecology of Invasions by Animals and Plants; Methuen: London, UK, 1958; 181p. [Google Scholar]
- Levine, J.M.; Adler, P.B.; Yelenik, S.G. A meta-analysis of biotic resistance to exotic plant invasions. Ecol. Lett. 2004, 7, 975–989. [Google Scholar] [CrossRef] [Green Version]
- Maron, J.L.; Vilà, M. When do herbivores affect plant invasion? Evidence for the natural enemies and biotic resistance hypotheses. Oikos 2001, 95, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Parker, J.D.; Hay, M.E. Biotic resistance to plant invasions? Native herbivores prefer non-native plants. Ecol. Lett. 2005, 8, 959–967. [Google Scholar] [CrossRef]
- Valéry, L.; Fritz, H.; Lefeuvre, J.C.; Simberloff, D. In search of a real definition of the biological invasion phenomenon itself. Biol. Invasions 2008, 10, 1345–1351. [Google Scholar] [CrossRef]
- Fleming, J.P.; Dibble, E.D. Ecological mechanisms of invasion success in aquatic macrophytes. Hydrobiologia 2015, 746, 23–37. [Google Scholar] [CrossRef]
- MacArthur, R.; Levins, R. The limiting similarity, convergence, and divergence of coexisting species. Am. Nat. 1967, 101, 377–385. [Google Scholar] [CrossRef]
- Spencer, D.F.; Rejmanek, M. Propagule type influences competition between two submersed aquatic macrophytes. Oecologia 1989, 81, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Petruzzella, A.; Manschot, J.; van Leeuwen, C.H.A.; Grutters, B.M.C.; Bakker, E.S. Mechanisms of invasion resistance of aquatic plant communities. Front. Plant Sci. 2018, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Madsen, J.D. Predicting invasion success of Eurasian watermilfoil. J. Aquat. Plant Manag. 1998, 36, 28–32. [Google Scholar] [CrossRef]
- Crawley, M.J.; Harvey, P.H.; Purvis, A. Comparative ecology of the native and alien floras of the British Isles. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1996, 351, 1251–1259. [Google Scholar] [CrossRef]
- Lloret, F.; Medail, F.; Brundu, G.; Camarda, I.; Moragues, E.; Rita, J.; Lambdon, P.; Hulme, P. Species attributes and invasion success by alien plants on Mediterranean islands. J. Ecol. 2005, 93, 512–520. [Google Scholar] [CrossRef]
- Mack, R.N. Phylogenetic constraint, absent life forms, and preadapted alien plants: A prescription for biological invasions. Int. J. Plant Sci. 2003, 164, 185–196. [Google Scholar] [CrossRef]
- Muthukrishnan, R.; Hansel-Welch, N.; Larkin, D.J.; Nilsson, C. Environmental filtering and competitive exclusion drive biodiversity-invasibility relationships in shallow lake plant communities. J. Ecol. 2018, 106, 2058–2070. [Google Scholar] [CrossRef]
- Grosholz, E. Avoidance by grazers facilitates spread of an invasive hybrid plant. Ecol. Lett. 2010, 13, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Pulzatto, M.M.; Lolis, L.A.; Louback-Franco, N.; Mormul, R.P. Herbivory on freshwater macrophytes from the perspective of biological invasions: A systematic review. Aquat. Ecol. 2018, 52, 297–309. [Google Scholar] [CrossRef]
- Miller, K.A.; Engle, J.M.; Uwai, S.; Kawai, H. First report of the Asian seaweed Sargassum filicinum Harvey (Fucales) in California, USA. Biol. Invasions 2007, 9, 609–613. [Google Scholar] [CrossRef] [Green Version]
- Marks, L.M.; Salinas-Ruiz, P.; Reed, D.C.; Holbrook, S.J.; Culver, C.S.; Engle, J.M.; Kushner, D.J.; Caselle, J.E.; Freiwald, J.; Williams, J.P.; et al. Range expansion of a non-native, invasive macroalga Sargassum horneri (Turner) C. Agardh, 1820 in the eastern Pacific. BioInvasions Rec. 2015, 4, 243–248. [Google Scholar] [CrossRef]
- Baker, H.G. Characteristics and modes of origin of weeds. In The Genetics of Colonizing Species; Baker, H.G., Stebbins, G.L., Eds.; Academic Press: New York, NY, USA, 1965; Volume X, pp. 147–168. [Google Scholar]
- Marks, L.M.; Reed, D.C.; Holbrook, S.J. Life history traits of the invasive seaweed Sargassum horneri at Santa Catalina Island, California. Aquat. Invasions 2018, 13, 339–350. [Google Scholar] [CrossRef]
- DeWreede, R.E. Sargassum muticum (Fucales, Phaeophyta): Regrowth and interaction with Rhodomelalarix (Ceramiales, Rhodophyta). Phycologia 1983, 22, 153–160. [Google Scholar] [CrossRef]
- Casas, G.; Scrosati, R.; Piriz, L. The invasive kelp Undaria pinnatifida (Phaeophyceae, Laminariales) reduces native seaweed diversity in Nuevo Gulf (Patagonia, Argentina). Biol. Invasions 2004, 6, 411–416. [Google Scholar] [CrossRef]
- Britton-Simmons, K.H. Direct and indirect effects of the introduced alga Sargassum muticum on benthic, subtidal communities of Washington State, USA. Mar. Ecol. Prog. Ser. 2004, 27, 61–78. [Google Scholar] [CrossRef] [Green Version]
- Mann, K.H. Destruction of kelp beds by sea urchins: A cyclical phenomenon or irreversible degradation? Helgolander Wissenshaftlichen Meeresuntersuchungen 1977, 30, 455–732. [Google Scholar] [CrossRef] [Green Version]
- Shears, N.T.; Babcock, R.C. Marine reserves demonstrate top-down control of community structure on temperate reefs. Oecologia 2002, 132, 131–142. [Google Scholar] [CrossRef]
- Lafferty, K.D. Fishing for lobsters indirectly increases epidemics in sea urchins. Ecol. Appl. 2004, 14, 1566–1573. [Google Scholar] [CrossRef]
- Geiselman, J.A.; McConnell, O.J. Polyphenols in brown algae Fucus vesiculosus and Ascophyllum nodusum: Chemical defenses against the marine herbivorous snail, Littorina littorea. J. Chem. Ecol. 1981, 8, 1115–1133. [Google Scholar] [CrossRef] [PubMed]
- Alstyne, K.L. Herbivore grazing increases polyphenolic defenses in the intertidal brown algae Fucus distichus. Ecology 1988, 69, 655–663. [Google Scholar] [CrossRef]
- Estes, J.A.; Steinberg, P.D. Predation, herbivory, and kelp evolution. Paleobiology 1988, 4, 19–36. [Google Scholar] [CrossRef]
- Van Alstyne, K.L.; McCarthy, J.J., III; Hustead, C.L.; Duggins, D.O. Geographic variation in polyphenolic levels of Northeastern Pacific kelps and rockweeds. Mar. Biol. 1999, 133, 371–379. [Google Scholar] [CrossRef]
- Vance, R.R.; Schmitt, R.J. The effect of the predator-avoidance behavior of the sea urchin, Centrostephanus coronatus, on the breadth of its diet. Oecologia 1979, 44, 21–25. [Google Scholar] [CrossRef]
- Harrer, S.L.; Reed, D.C.; Miller, R.J.; Holbrook, S.J. Patterns and controls of the dynamics of net primary production by understory macroalgal assemblages in giant kelp forests. J. Phycol. 2013, 49, 248–257. [Google Scholar] [CrossRef]
- Nelson, J.; Reed, D.; Harrer, S.; Miller, R. SBC LTER: Reef: Coefficients for estimating biomass from body size or percent cover for kelp forest species. Environ. Data Initiat. 2020, in press. [Google Scholar]
- Santa Barbara Coastal Long Term Ecological Research; SBC LTER (University of California Santa Barbara): Santa Barbara, CA, USA, 2019; Unpublished data.
- Thomsen, M.S.; Wernberg, T.; South, P.M.; Schiel, D.R. To include or not to include (the invader in community analyses)? that is the question. Biol. Invasions 2016, 18, 1515–1521. [Google Scholar] [CrossRef]
- Gelman, A.; Hill, J. Data Analysis Using Regression and Multilevel/Hierarchical Models, 2nd ed.; Cambridge University Press: London, UK, 2006; p. 648. [Google Scholar]
- RStudio Team (2019). RStudio: Integrated Development for R. RStudio, Inc.: Boston, MA, USA. Available online: http://www.rstudio.com/ (accessed on 14 December 2019).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, K.R.; Gorley, R.N. PRIMER v7: User Manual/Tutorial, 1st ed.; PRIMER-E: Plymouth, UK, 2015; p. 300. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2018; p. 214. [Google Scholar]
- Levine, J.M.; Vilà, M.; D’Antonio, C.M.; Dukes, J.S.; Grigulis, K.; Lavorel, S. Mechanisms underlying the impacts of exotic plant invasions. Proc. R. Soc. Lond. B 2003, 270, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Vilà, M.; Weiner, J. Are invasive plant species better competitors than native plant species? – evidence from pair-wise experiments. Oikos 2004, 105, 229–238. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Wernberg, T.; Tuya, F.; Silliman, B.R. Evidence for impacts of nonindigenous macroalgae: A meta-analysis of experimental field studies. J. Phycol. 2009, 45, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Theuekauf, S.J.; Pukett, B.J.; Theuekauf, K.W.; Theuerkauf, E.J.; Eggleston, D.B. Density-dependent role of an invasive marsh grass, Phragmites australis, on ecosystem service provision. PLoS ONE 2017, 12, e0173007. [Google Scholar] [CrossRef] [Green Version]
- White, L.F.; Shurin, J.B. Density dependent effects of an exotic marine macroalga on native community diversity. J. Exp. Mar. Biol. Ecol. 2011, 405, 111–119. [Google Scholar] [CrossRef]
- Olson, E.R.; Doherty, J.M. Macrophyte diversity-abundance relationship with respect to invasive and native dominants. Aquat. Bot. 2014, 119, 111–119. [Google Scholar] [CrossRef]
- Shea, K.; Chesson, P. Community ecology theory as a framework for biological invasions. Trends Ecol. Evol. 2002, 17, 170–176. [Google Scholar] [CrossRef]
- Benes, K.M.; Carpenter, R.C. Kelp canopy facilitates understory algal assemblage via competitive release during early stages of secondary succession. Ecology 2015, 96, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Reed, D.C.; Rassweiler, A.; Arkema, K. Density derived estimates of standing crop and net primary production in the giant kelp Macrocystis pyrifera. Mar. Biol. 2009, 156, 2077–2083. [Google Scholar] [CrossRef] [Green Version]
- Kastendiek, J. Competitor-mediated coexistence: Interactions among three species of benthic macroalgae. J. Exp. Mar. Biol. Ecol. 1982, 62, 201–210. [Google Scholar] [CrossRef]
- Anderson, T.W. Role of macroalgal structure in the distribution and abundance of a temperate reef fish. Mar. Ecol. Prog. Ser. 1994, 113, 279–290. [Google Scholar] [CrossRef]
- Lafferty, K.D.; Behrens, M.D.; Davis, G.E.; Haaker, P.L.; Kushner, D.J.; Richards, D.V.; Taniguchi, I.K.; Tegner, M.J. Habitat of endangered white abalone, Haliotis sorenseni. Biol. Conservation 2004, 116, 191–194. [Google Scholar] [CrossRef]
- Morrow, K.; Carpenter, R. Shallow kelp canopies mediate macroalgal composition: effects on the distribution and abundance of Corynactis californica (Corallimorpharia). Mar. Ecol. Prog. Ser. 2008, 361, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T. Japanese species of Sargassum subgenus Bactrophycus (Phaeophyta, Fucales). J. Fac. of Sci., Hokkaido Univ. Ser. V (Botany) 1983, 13, 99–246. [Google Scholar]
- Tkaba, M.; Mizokami, A. Seasonal fluctuation of Sargassum communities and vertical distribution of Sargassaceae at Kuroshima Island in the western Akinada. Bull. Hiroshima. Fish. Exp. Stn. 1982, 12, 33–44. [Google Scholar]
- Aguilar-Rosas, L.E.; Aguilar-Rosas, R.; Kawai, H.; Uwai, S.; Valenzuela-Espinoza, E. New record of Sargassum filicinum Harvey (Fucales, Phaeophyceae) in the Pacific coast of Mexico. Algae 2007, 22, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Trejo, G.I.; Ibarra-Obando, S.E.; Aguilar-Rosas, L.E.; Poumian-Tapia, M.; Solana-Arellano, E. Presence of Sargassum horneri at Todos Santos Bay, Baja California, Mexico: its effects on the local macroalgae community. Am. J. Plant Sci. 2015, 6, 2693–2707. [Google Scholar] [CrossRef] [Green Version]
- Riosmena-Rodríguez, R.; Boo, G.H.; López-Vivas, J.M.; Hernández-Velasco, A.; Sáenz-Arroyo, A.; Boo, S.M. The invasive seaweed Sargassum filicinum (Fucales, Phaeophyceae) is on the move along the Mexican Pacific coastline. Bot. Mar. 2012, 55, 547–551. [Google Scholar] [CrossRef]
- Epstein, G.; Hawkins, S.; Smale, D. Identifying niche and fitness dissimilarities in invaded marine macroalgal canopies within the context of contemporary coexistence theory. Sci. Rep. 2019, 9, 8816. [Google Scholar] [CrossRef] [Green Version]
- Sullaway, G.; Edwards, M. Impacts of the non-native alga, Sargassum horneri, on benthic community production in a California Kelp Forest. Mar. Ecol. Prog. Ser. 2020, in press. [Google Scholar] [CrossRef]
- MacDougall, A.S.; Turkington, R. Are invasive species the drivers or passengers of change in degraded ecosystems? Ecology 2005, 86, 42–55. [Google Scholar] [CrossRef] [Green Version]
- Bulleri, F.; Tamburello, L.; Beneditti-Cecchi, L. Loss of consumers alters the effects of resident assemblages on the local spread of an introduced macroalga. Oikos 2009, 118, 269–279. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M. Traits associated with invasiveness in alien plants: where do we stand. In Biol. Invasions; Nentwig, W., Ed.; Springer-Verlag: Berlin, Germany, 2007; pp. 97–125. [Google Scholar] [CrossRef]
- Koricheva, J. Meta-analysis of sources of variation in fitness costs of plant antiherbivore defenses. Ecology 2002, 83, 176–190. [Google Scholar] [CrossRef]
- Strauss, S.; Rudgers, J.A.; Lau, J.A.; Irwin, R.E. Direct and ecological costs of resistance to herbivory. Trends Ecol. Evol. 2002, 17, 278–285. [Google Scholar] [CrossRef]
- Marks, L.; Reed, D.; Holbrook, S. SBC LTER: REEF: Data to support "Niche complementarity and resistance to grazing promote the invasion success of Sargassum horneri in North America". Environ. Data Initiat 2020. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Model | df | AIC | χ2 | Chi df | p (<χ 2) |
|---|---|---|---|---|---|---|
| a. Biomass of native algae1 | ||||||
| Individual parameters | Null | 4 | 2140.4 | |||
| Days | 5 | 2139.9 | 2.52 | 1 | 0.112 | |
| Removal | 5 | 2141.9 | 0.51 | 1 | 0.473 | |
| Season | 7 | 2106.3 | 40.22 | 3 | < 0.001 | |
| Interactions | Days + Removal | 6 | 2141.4 | |||
| Days × Removal | 7 | 2132.2 | 11.15 | 1 | < 0.001 | |
| Season + Removal | 8 | 2107.7 | ||||
| Season × Removal | 11 | 2104.7 | 9.02 | 3 | 0.029 | |
| b. Richness of native algae | ||||||
| Individual parameters | Null | 4 | 1489.7 | |||
| Days | 5 | 1403.8 | 87.83 | 1 | < 0.001 | |
| Removal | 5 | 1491.6 | 0.04 | 1 | 0.842 | |
| Season | 7 | 1447.1 | 48.54 | 3 | < 0.001 | |
| Interactions | Days + Removal | 6 | 1405.8 | |||
| Days × Removal | 7 | 1407.1 | 0.72 | 1 | 0.397 | |
| Season + Removal | 8 | 1449.1 | ||||
| Season × Removal | 11 | 1446.6 | 8.54 | 3 | 0.036 |
| Taxonomic | Taxon | Spring | Summer | ||||
|---|---|---|---|---|---|---|---|
| Group | S+ | S− | % | S+ | S− | % | |
| Subcanopy | Stephanocystis neglecta | 25.9 ± 15.3 | 27.2 ± 11.3 | 7.6 | 27.5 ± 10.3 | 19.0 ± 8.1 | 5.9 |
| algae | Sargassum palmeri | 415.1 ± 154.1 | 911.3 ± 232.5 | 29.0 | 172.3 ± 45.8 | 262.6 ± 67.6 | 14.3 |
| Understory algae | Articulated coralline spp. | 0 ± 0 | 1.6 ± 1.6 | . | 0.4 ± 0.4 | 1.6 ± 1.6 | . |
| Asparagopsis taxiformis | 0 ± 0 | 0 ± 0 | . | 0.3 ± 0.3 | 1.1 ± 1.1 | . | |
| Brown blade spp. | 0 ± 0 | 6.0 ± 4.3 | . | 0 ± 0 | 0 ± 0 | . | |
| Cladophora graminea | 0 ± 0 | 0 ± 0 | . | 0.1 ± 0.1 | 0 ± 0 | . | |
| Colpomenia sinuosa | 0 ± 0 | 10.6 ± 8.4 | . | 148.3 ± 72.3 | 12.7 ± 5.4 | 12.3 | |
| Chondria californica | 0 ± 0 | 0.3 ± 0.3 | 1.1 ± 0.5 | 0.5 ± 0.4 | |||
| Corallina chilensis | 20.2 ± 10.6 | 30.5 ± 13.0 | 7.2 | 13.6 ± 7.4 | 12.3 ± 7.0 | . | |
| Dictyopteris undulata | 3.9 ± 2.2 | 23.5 ± 6.1 | 6.6 | 14.8 ± 6.7 | 45.9 ± 12.4 | 7.2 | |
| Dictyota spp. | 1.1 ± 0.7 | 20.4 ± 7.7 | . | 24.0 ± 11.2 | 24.0 ± 11.6 | . | |
| Filamentous brown spp. | 0 ± 0 | 0 ± 0 | . | 0.1 ± 0.1 | 0 ± 0 | . | |
| Filamentous green spp. | 0 ± 0 | 0.2 ± 0.2 | . | 0.2 ± 0.2 | 0.5 ± 0.3 | . | |
| Filamentous red spp. | 1.1 ± 1.1 | 1.2 ± 1.0 | . | 0 ± 0 | 2.4 ± 1.9 | . | |
| Green foliose spp. | 0 ± 0 | 0.5 ± 0.5 | . | 0 ± 0 | 0 ± 0 | . | |
| Halicystis ovalis | 0 ± 0 | 0 ± 0 | . | 0.2 ± 0.2 | 0.7 ± 0.4 | . | |
| Haliptylon gracile | 20.6 ± 11.5 | 20.4 ± 7.9 | . | 37.8 ± 21.3 | 28.7 ± 13.4 | 6.6 | |
| Hydroclathrus clathratus | 0 ± 0 | 0 ± 0 | . | 8.5 ± 4.2 | 5.3 ± 4.3 | . | |
| Laurencia pacifica | 0.6 ± 0.6 | 3.6 ± 1.1 | . | 7.7 ± 3.0 | 11.3 ± 5.2 | . | |
| Lithothrix aspergillum | 19.8 ± 10.0 | 21.0 ± 11.5 | 6.5 | 33.7 ± 20.9 | 25.1 ± 15.0 | 6.3 | |
| Plocamium cartilagineum | 2.7 ± 2.3 | 2.2 ± 1.5 | . | 17.3 ± 10.5 | 2.4 ± 1.4 | . | |
| Pterocladia capillacea | 4.6 ± 4.6 | 5.3 ± 4.3 | . | 8.5 ± 4.2 | 6.4 ± 4.3 | . | |
| Rhodymenia californica | 0 ± 0 | 1.9 ± 1.3 | . | 1 ± 0.5 | 0.6 ± 0.4 | . | |
| Scytosiphon lomentaria | 0 ± 0 | 0 ± 0 | . | 1 ± 0.7 | 0 ± 0 | . | |
| Zonaria farlowii | 171.9 ± 34.9 | 347.6 ± 72.2 | 16.2 | 182.7 ± 47.1 | 513.2 ± 84.7 | 17.7 | |
| Cumulative % contribution to dissimilarity | - | - | 73.1 | - | - | 70.3 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marks, L.M.; Reed, D.C.; Holbrook, S.J. Niche Complementarity and Resistance to Grazing Promote the Invasion Success of Sargassum horneri in North America. Diversity 2020, 12, 54. https://doi.org/10.3390/d12020054
Marks LM, Reed DC, Holbrook SJ. Niche Complementarity and Resistance to Grazing Promote the Invasion Success of Sargassum horneri in North America. Diversity. 2020; 12(2):54. https://doi.org/10.3390/d12020054
Chicago/Turabian StyleMarks, Lindsay M., Daniel C. Reed, and Sally J. Holbrook. 2020. "Niche Complementarity and Resistance to Grazing Promote the Invasion Success of Sargassum horneri in North America" Diversity 12, no. 2: 54. https://doi.org/10.3390/d12020054
APA StyleMarks, L. M., Reed, D. C., & Holbrook, S. J. (2020). Niche Complementarity and Resistance to Grazing Promote the Invasion Success of Sargassum horneri in North America. Diversity, 12(2), 54. https://doi.org/10.3390/d12020054