Green Fluorescence Patterns in Closely Related Symbiotic Species of Zanclea (Hydrozoa, Capitata)

 ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Analyses and Fluorescence Essay
2.2. Molecular Characterization
3. Results
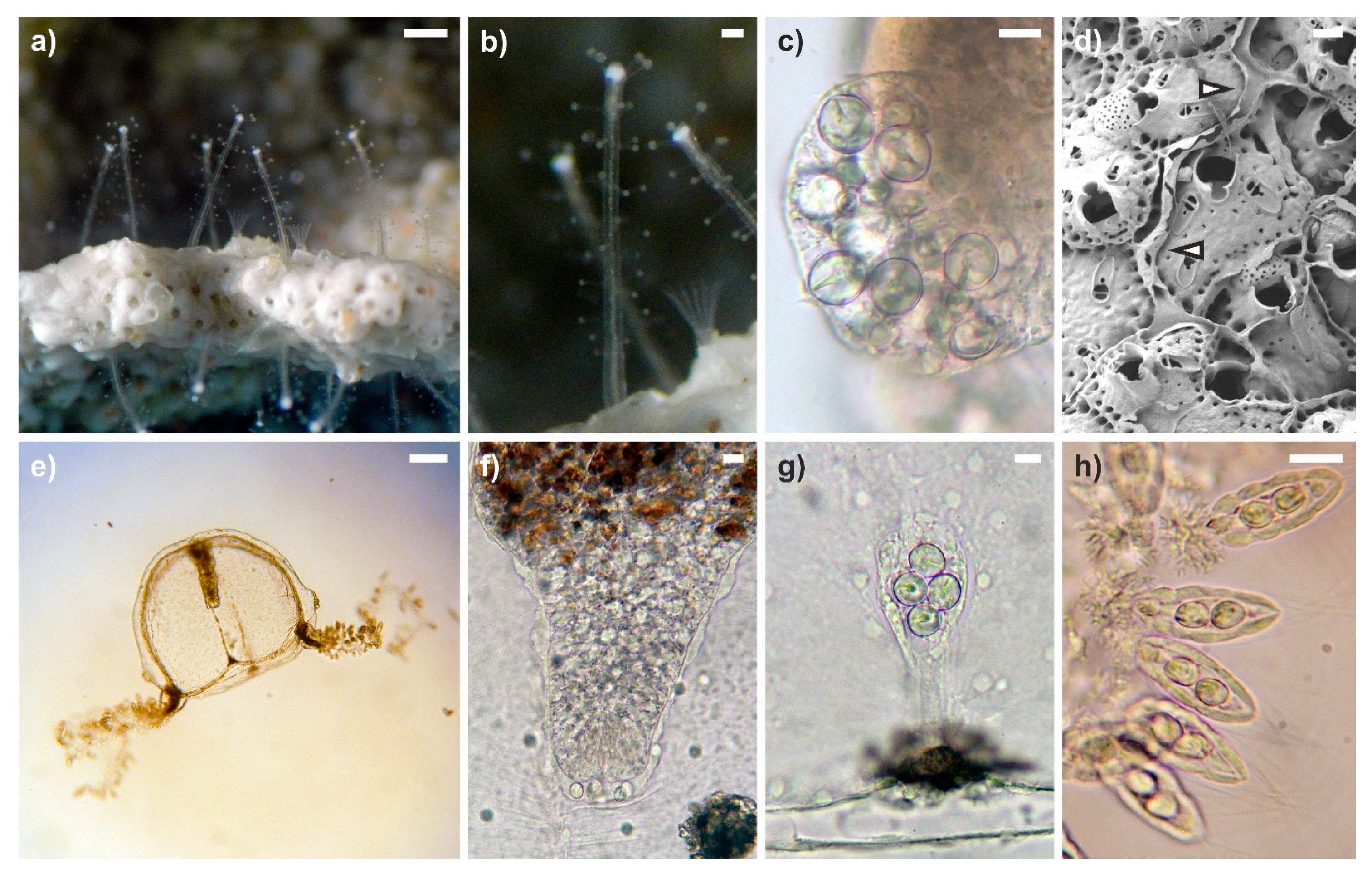
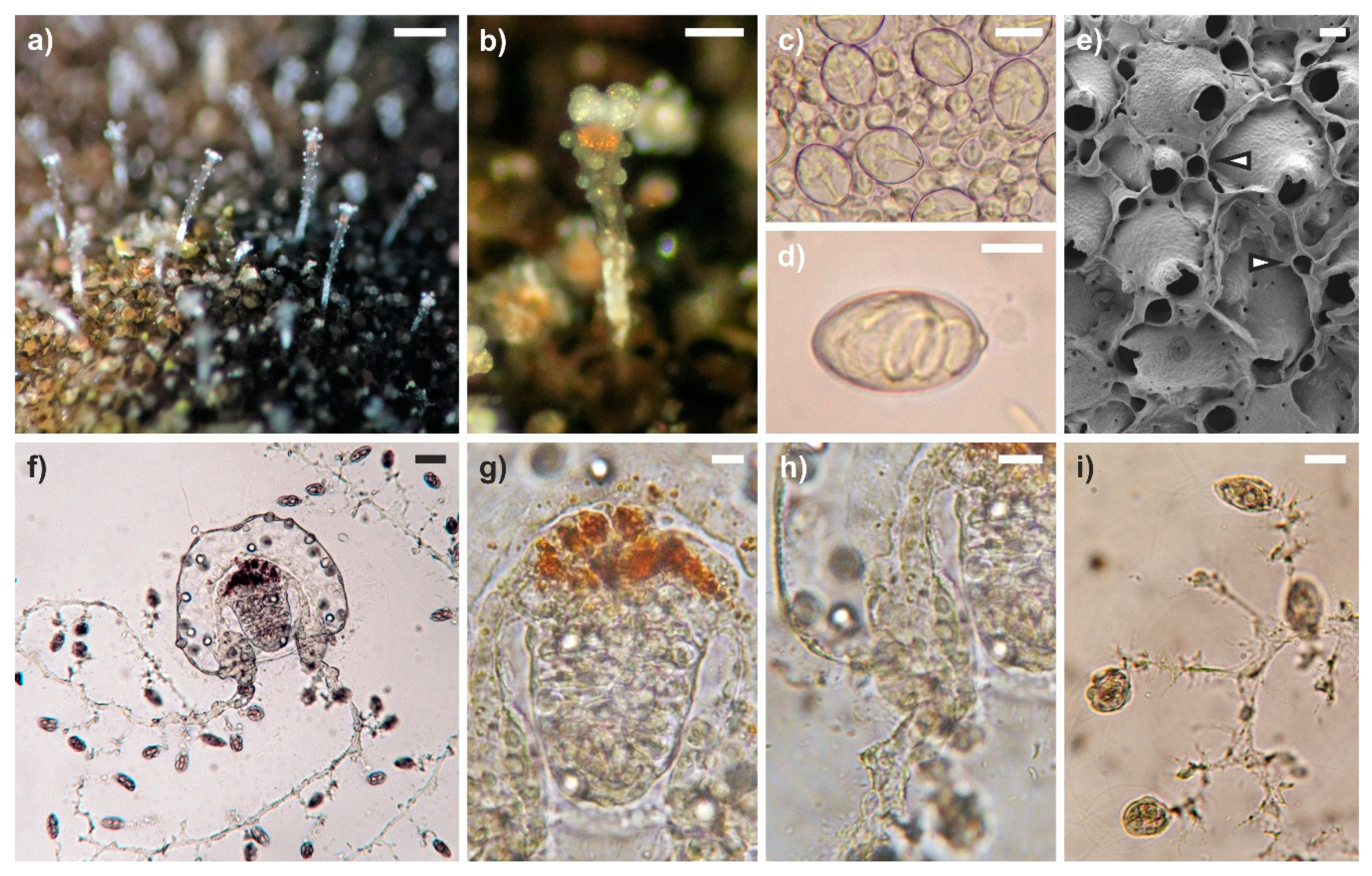
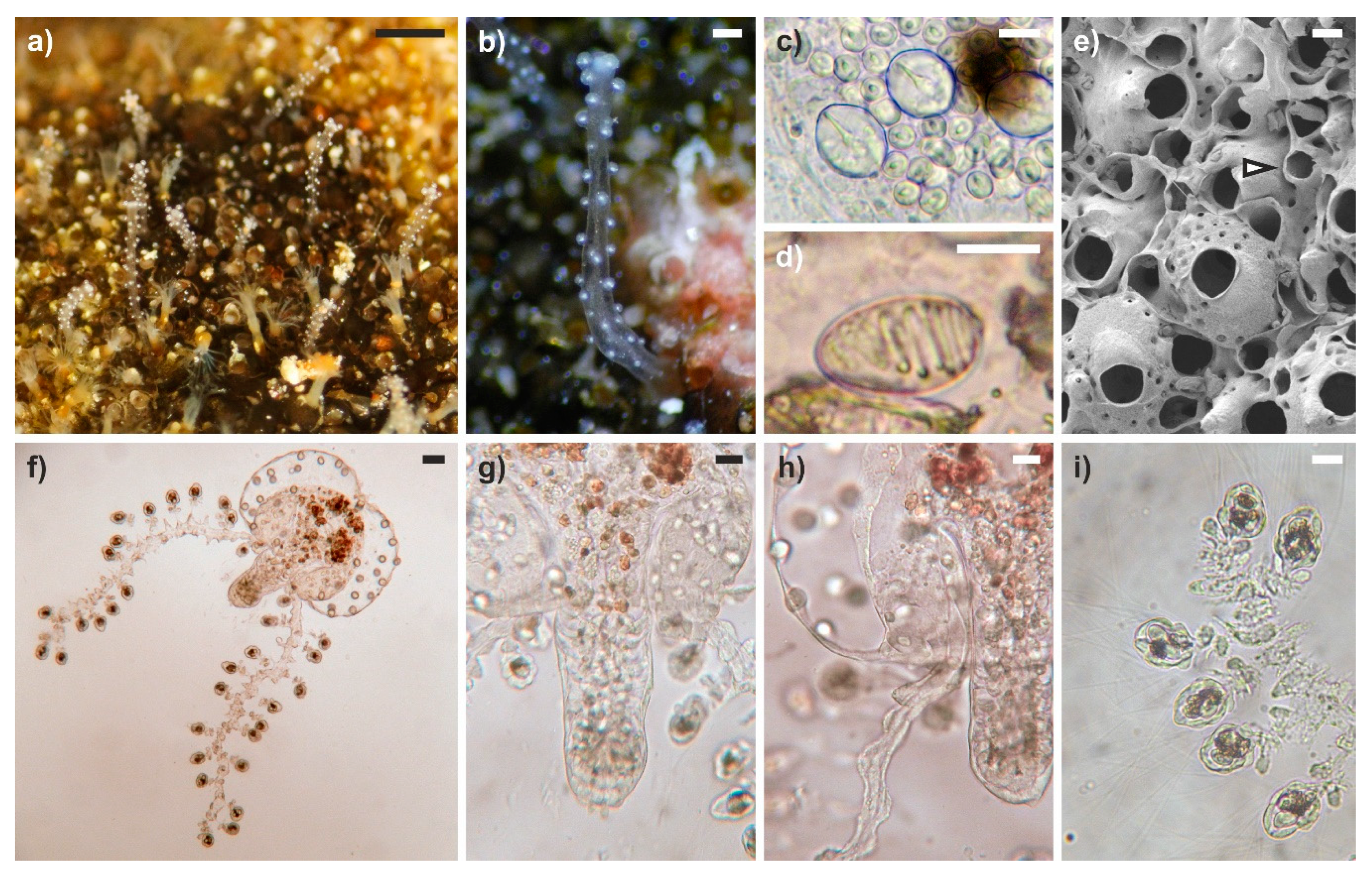
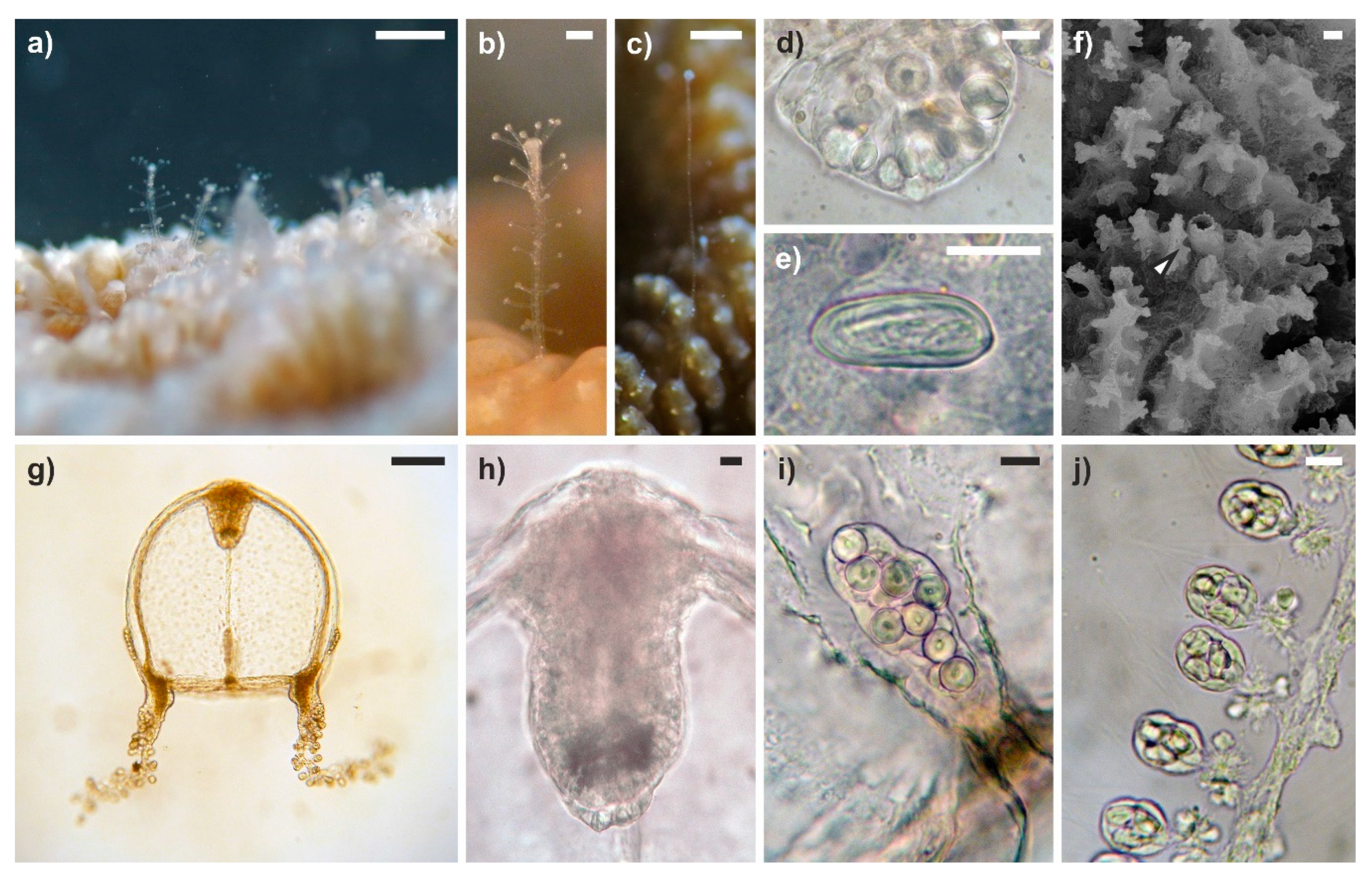
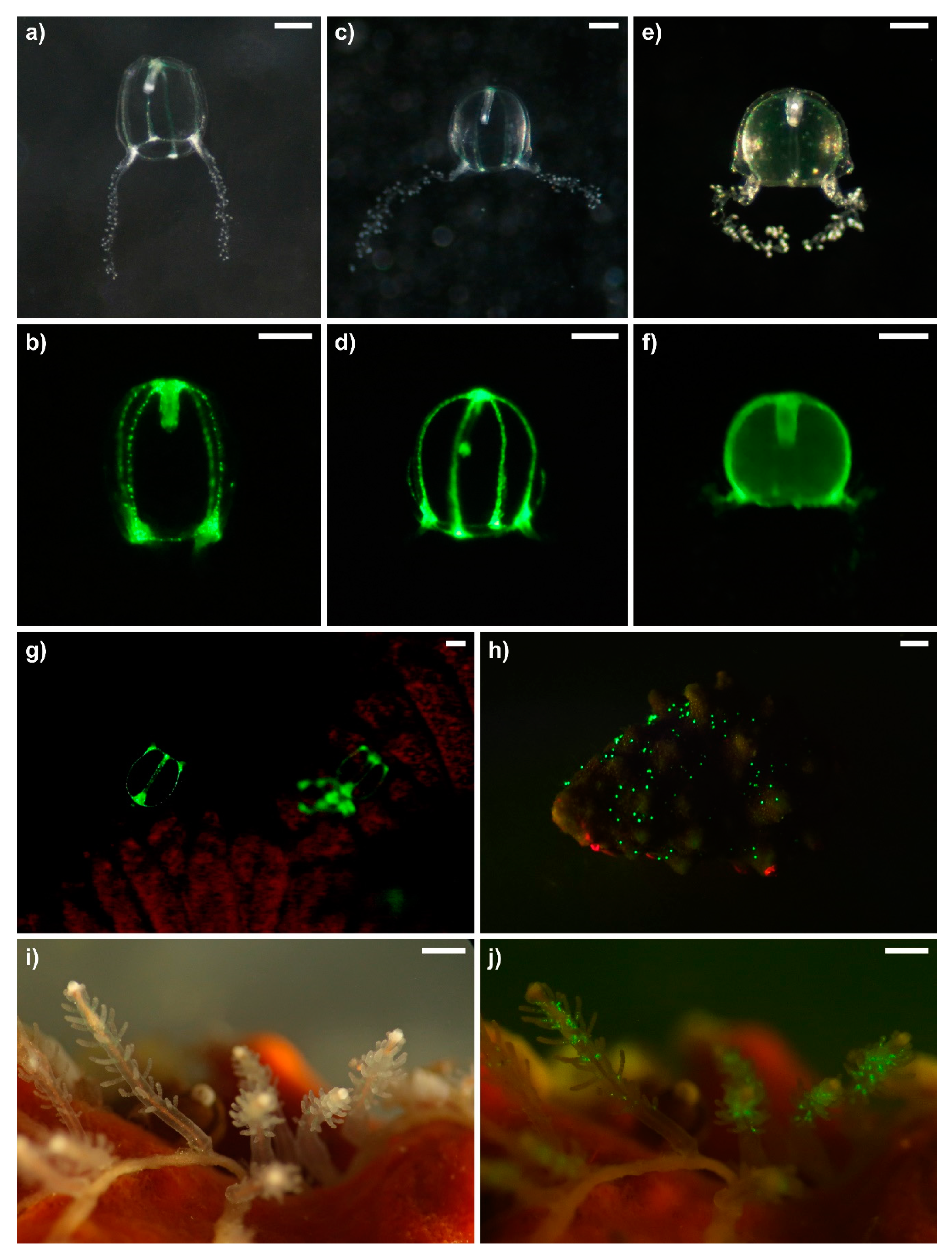
3.1. General Morphology of Polyps and Medusae
3.2. Modifications of the Hosts
3.3. Green Fluorescence Essay
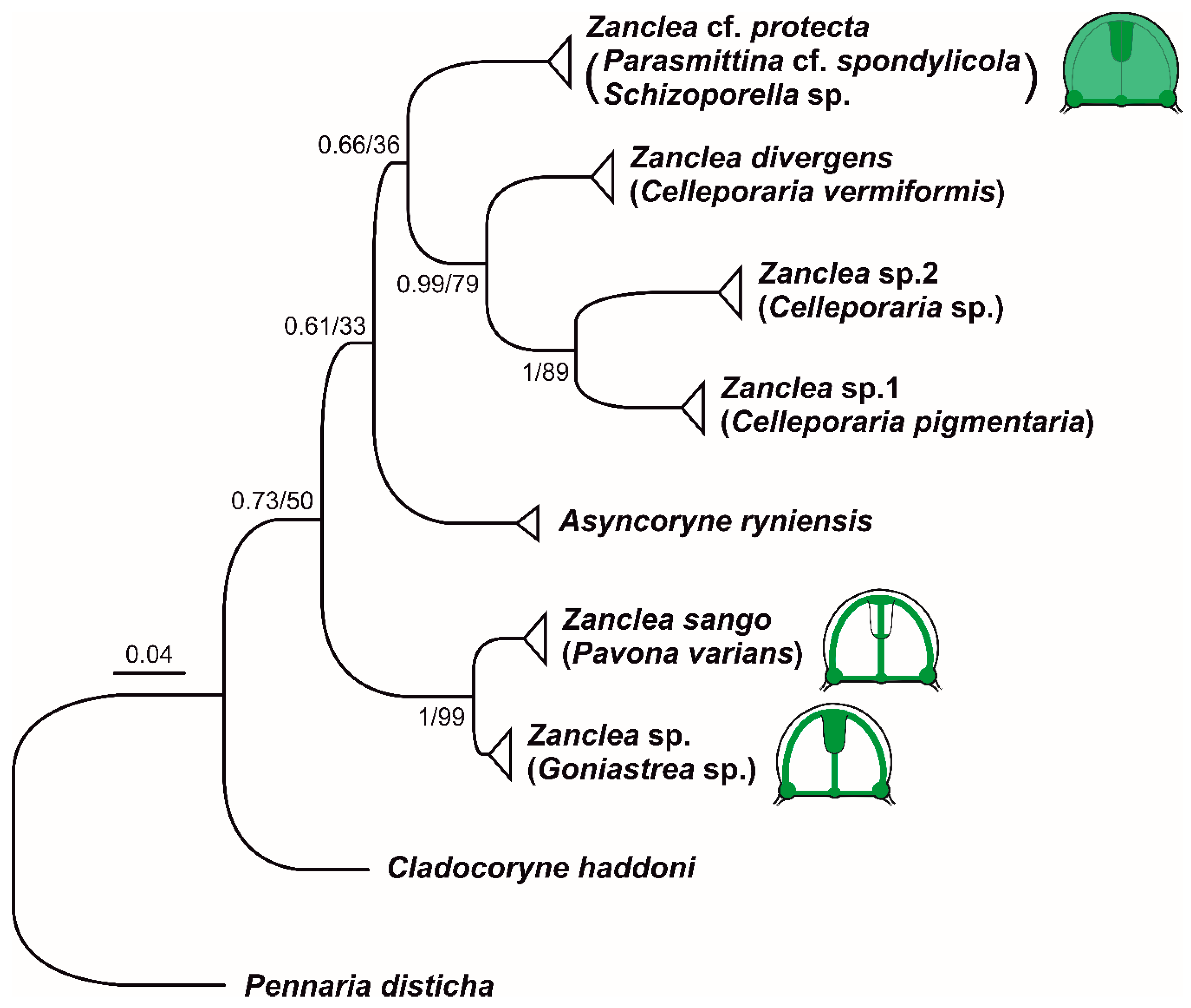
3.4. 16S rRNA Phylogeny
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chudakov, D.M.; Matz, M.V.; Lukyanov, S.; Lukyanov, K.A. Fluorescent proteins and their applications in imaging living cells and tissues. Physiol. Rev. 2010, 90, 1103–1163. [Google Scholar] [CrossRef]
- Shimomura, O.; Johnson, F.H.; Saiga, Y. Extraction, purification and properties of aequorin, a bioluminescent protein from the luminous hydromedusan, Aequorea. J. Cell. Comp. Physiol. 1962, 59, 223–239. [Google Scholar] [CrossRef]
- Miyawaki, A. Green fluorescent protein-like proteins in reef Anthozoa animals. Cell Struct. Funct. 2002, 27, 343–347. [Google Scholar] [CrossRef] [Green Version]
- Shagin, D.A.; Barsova, E.V.; Yanushevich, Y.G.; Fradkov, A.F.; Lukyanov, K.A.; Labas, Y.A.; Semenova, T.N.; Ugalde, J.A.; Meyers, A.; Nunez, J.M.; et al. GFP-like proteins as ubiquitous metazoan superfamily: evolution of functional features and structural complexity. Mol. Biol. Evol. 2004, 21, 841–850. [Google Scholar] [CrossRef]
- Salih, A.; Larkum, A.; Cox, G.; Kühl, M.; Hoegh-Guldberg, O. Fluorescent pigments in corals are photoprotective. Nature 2000, 408, 850. [Google Scholar] [CrossRef]
- Morin, J.G.; Hastings, J.W. Energy transfer in a bioluminescent system. J. Cell. Physiol. 1971, 77, 313–318. [Google Scholar] [CrossRef]
- Sparks, J.S.; Schelly, R.C.; Smith, W.L.; Davis, M.P.; Tchernov, D.; Pieribone, V.A.; Gruber, D.F. The covert world of fish biofluorescence: A phylogenetically widespread and phenotypically variable phenomenon. PLoS ONE 2014, 9, e83259. [Google Scholar] [CrossRef] [Green Version]
- Haddock, S.H.; Dunn, C.W. Fluorescent proteins function as a prey attractant: experimental evidence from the hydromedusa Olindias formosus and other marine organisms. Biol. Open 2015, 4, 1094–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Brauwer, M.; Hobbs, J.P.A. Stars and stripes: biofluorescent lures in the striated frogfish indicate role in aggressive mimicry. Coral Reefs 2016, 35, 1171. [Google Scholar] [CrossRef] [Green Version]
- Prudkovsky, A.A.; Ivanenko, V.N.; Nikitin, M.A.; Lukyanov, K.A.; Belousova, A.; Reimer, J.D.; Berumen, M.L. Green fluorescence of Cytaeis hydroids living in association with Nassarius gastropods in the Red Sea. PLoS ONE 2016, 11, e0146861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubota, S.; Pagliara, P.; Gravili, C. Fluorescence distribution pattern allows to distinguish two species of Eugymnanthea (Leptomedusae: Eirenidae). J. Mar. Biol. Assoc. U. K. 2008, 88, 1743–1746. [Google Scholar] [CrossRef] [Green Version]
- Kubota, S. Various distribution patterns of green fluorescence in small hydromedusae. Kuroshio Biosphere 2010, 6, 11–14. [Google Scholar]
- Kubota, S.; Gravili, C. Rare distribution of green fluorescent protein (GFP) in hydroids from Porto Cesareo, Lecce, Italy, with reference to biological meaning of this rarity. Biogeogr. 2011, 13, 9–11. [Google Scholar]
- Ryusaku, D.; Mayu, O.; Hiroshi, N. Green fluorescent protein (GFP)-like substance in the hydrozoan jellyfish Cytaeis uchidae: examination of timing and localization of its expression and utilization for biological education. Bull. Miyagi Univ. Educ. 2012, 47, 95–100. [Google Scholar]
- Maggioni, D.; Montano, S.; Seveso, D.; Galli, P. Molecular evidence for cryptic species in Pteroclava krempfi (Hydrozoa, Cladocorynidae) living in association with alcyonaceans. Syst. Biodivers. 2016, 14, 484–493. [Google Scholar] [CrossRef]
- Montano, S.; Maggioni, D.; Galli, P.; Hoeksema, B.W. A cryptic species in the Pteroclava krempfi species complex (Hydrozoa, Cladocorynidae) revealed in the Caribbean. Mar. Biodivers. 2017, 47, 83–89. [Google Scholar] [CrossRef]
- Miglietta, M.P.; Maggioni, D.; Matsumoto, Y. Phylogenetics and species delimitation of two hydrozoa (phylum Cnidaria): Turritopsis (McCrady, 1857) and Pennaria (Goldfuss, 1820). Mar. Biodivers. 2019, 49, 1085–1100. [Google Scholar] [CrossRef]
- Montano, S.; Maggioni, D.; Arrigoni, R.; Seveso, D.; Puce, S.; Galli, P. The hidden diversity of Zanclea associated with scleractinians revealed by molecular data. PLoS ONE 2015, 10, e0133084. [Google Scholar] [CrossRef] [Green Version]
- Maggioni, D.; Montano, S.; Arrigoni, R.; Galli, P.; Puce, S.; Pica, D.; Berumen, M.L. Genetic diversity of the Acropora-associated hydrozoans: new insight from the Red Sea. Mar. Biodivers. 2017, 47, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Boero, F.; Bouillon, J.; Gravili, C. A survey of Zanclea, Halocoryne and Zanclella (Cnidaria, Hydrozoa, Anthomedusae, Zancleidae) with description of new species. Ital. J. Zool. 2000, 67, 93–124. [Google Scholar] [CrossRef]
- Manca, F.; Puce, S.; Caragnano, A.; Maggioni, D.; Pica, D.; Seveso, D.; Galli, P.; Montano, S. Symbiont footprints highlight the diversity of scleractinia - associated Zanclea hydrozoans (Cnidaria, Hydrozoa). Zool. Scr. 2019, 48, 399–410. [Google Scholar] [CrossRef]
- Maggioni, D.; Arrigoni, R.; Galli, P.; Berumen, M.L.; Seveso, D.; Montano, S. Polyphyly of the genus Zanclea and family Zancleidae (Hydrozoa, Capitata) revealed by the integrative analysis of two bryozoan-associated species. Contrib. Zool. 2018, 87, 87–104. [Google Scholar] [CrossRef] [Green Version]
- Zietara, M.S.; Arndt, A.; Geets, A.; Hellemans, B.; Volckaert, F.A. The nuclear rDNA region of Gyrodactylus arcuatus and G. branchicus (Monogenea: Gyrodactylidae). J. Parasitol. 2000, 86, 1368–1373. [Google Scholar] [CrossRef]
- Maggioni, D.; Puce, S.; Galli, P.; Seveso, D.; Montano, S. Description of Turritopsoides marhei sp. nov. (Hydrozoa, Anthoathecata) from the Maldives and its phylogenetic position. Mar. Biol. Res. 2017, 13, 983–992. [Google Scholar] [CrossRef]
- Cunningham, C.W.; Buss, L.W. Molecular evidence for multiple episodes of paedomorphosis in the family Hydractiniidae. Biochem. Syst. Ecol. 1993, 21, 57–69. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: more models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Maggioni, D.; (Universoty of Milano-Bicocca). Personal Observation. 2020. [Google Scholar]
- Maggioni, D.; Galli, P.; Berumen, M.L.; Arrigoni, R.; Seveso, D.; Montano, S. Astrocoryne cabela, gen. nov. et sp. nov. (Hydrozoa: Sphaerocorynidae), a new sponge-associated hydrozoan. Invertebr. Syst. 2017, 31, 734–746. [Google Scholar] [CrossRef]
- Nawrocki, A.M.; Schuchert, P.; Cartwright, P. Phylogenetics and evolution of Capitata (Cnidaria: Hydrozoa), and the systematics of Corynidae. Zool. Scr. 2010, 39, 290–304. [Google Scholar] [CrossRef]
- Montano, S.; Arrigoni, R.; Pica, D.; Maggioni, D.; Puce, S. New insights into the symbiosis between Zanclea (Cnidaria, Hydrozoa) and scleractinians. Zool. Scr. 2015, 44, 92–105. [Google Scholar] [CrossRef]
- Hastings, A.B. LIV. On the association of a gymnoblastic Hydroid (Zanclea protecta, sp. n.) with various cheilostomatous polyzoa from the Tropical E. Pacific. Ann. Mag. Nat. Hist. 1930, 5, 552–560. [Google Scholar] [CrossRef]
- Montano, S.; Fattorini, S.; Parravicini, V.; Berumen, M.L.; Galli, P.; Maggioni, D.; Arrigoni, R.; Seveso, D.; Strona, G. Corals hosting symbiotic hydrozoans are less susceptible to predation and disease. Proc. R. Soc. B 2017, 284, 20172405. [Google Scholar] [CrossRef] [Green Version]
- Osman, R.W.; Haugsness, J.A. Mutualism among sessile invertebrates: a mediator of competition and predation. Science 1981, 211, 846–848. [Google Scholar] [CrossRef] [Green Version]
- Fontana, S.; Keshavmurthy, S.; Hsieh, H.J.; Denis, V.; Kuo, C.Y.; Hsu, C.M.; Leung, J.K.L.; Tsai, W.S.; Wallace, C.C.; Chen, C.A. Molecular evidence shows low species diversity of coral-associated hydroids in Acropora corals. PLoS ONE 2012, 7, e50130. [Google Scholar] [CrossRef] [Green Version]
- Puce, S.; Cerrano, C.; Boyer, M.; Ferretti, C.; Bavestrello, G. Zanclea (Cnidaria: Hydrozoa) species from Bunaken Marine Park (Sulawesi Sea, Indonesia). J. Mar. Biol. Assoc. U. K. 2002, 82, 943–954. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Zanclea divergens | Zanclea cf. protecta | Zanclea sp. 1 | Zanclea sp. 2 | Zanclea sango | Zanclea sp. (Clade I) | Asyncoryne ryniensis |
|---|---|---|---|---|---|---|---|
| Host/Substrate | Celleporaria vermiformis | Parasmittina cf. spondylicola, Schizoporella sp. | Celleporaria pigmentaria | Celleporaria sp. | Pavona varians | Goniastrea sp. | Rock, sponge |
| Hydrorhiza | Below the bryozoan skeleton, coming out in irregular notches | On the bryozoan, overgrown by the host skeleton | Below the bryozoan skeleton, coming out in irregular notches | Below the bryozoan skeleton, coming out for some distance | Below the coral skeleton and tissues | Below the coral skeleton and tissues | On the substrate |
| Perisarc | No | No | No | No | Yes | Yes | Yes |
| Gastrozooid | Cylindrical, up to 3.5 mm long | Cylindrical, up to 1.5 mm long | Claviform, up to 1.5 mm long | Cylindrical, up to 3 mm long | Cylindrical to claviform, up to 1 mm long | Cylindrical to claviform, up to 1 mm long | Cylindrical, up to 6 mm long |
| Gastrozooid tentacles | Capitate: 4–5 oral, 16–39 aboral | Capitate: 4–5 oral, 15–44 aboral | Capitate: 4–5 oral, 18–20 aboral | Capitate: 4–5 oral, 23–27 aboral | Capitate: 4–6 oral, 12–22 aboral | Capitate: 4–5 oral, 23–32 aboral | Capitate:3–4 oral; moniliform: 28–36 aboral |
| Dactylozooid | No | No | No | No | Up to 3 mm, globular apex, no tentacles | Up to 3 mm, globular apex, no tentacles | No |
| Medusa buds localization | Hydrorhiza | Hydrorhiza and polyps | Hydrorhiza | Hydrorhiza | Hydrorhiza and polyps | Hydrorhiza and polyps | Polyps |
| Color | Whitish-transparent, white hypostome | Transparent, white hypostome | Transparent, white band, whitish to orange hypostome | Transparent | Transparent, white hypostome | Transparent, white hypostome | Transparent, orange gastroderm, white hypostome |
| Stenoteles | Capitula | Capitula, hydrorhiza | Capitula, hydrorhiza | Capitula, hydrorhiza | Capitula, hydrorhiza, dactylozooid | Capitula, hydrorhiza, dactylozooid | Capitula, moniliform tentacles, body wall, hydrorhiza |
| Stenoteles size (μm) | 11–15 × 9–12, 5–8 × 4–6 | 11–15 × 8–13, 5–8 × 4–7 | 14–17 × 12–13, 6–7 × 4–6 | 18–20 × 12–18, 15–16 × 12–15, 6–7 × 4–6 | 10–14 × 9–14, 6–8 × 5–6 | 10–14 × 8–13, 6–9 × 4–6 | 30–32 × 26–29, 10–11 × 7–8, 8–9 × 6–7 |
| Euryteles | Macrobasic holotrichous, in hypostome, hydrorhiza | No | Macrobasic holotrichous, in hydrorhiza | Macrobasic holotrichous, in hydrorhiza | Macrobasic apotrichous, in hypostome, hydrorhiza, dactylozooid | Macrobasic apotrichous, in hypostome, hydrorhiza, dactylozooid | Macrobasic holotrichous, in body wall, hydrorhiza |
| Euryteles size (μm) | 29–33 × 16–18 | No | 27–29 × 14–16 | 18–21 × 11–15 | 17–21 × 6–10 | 16–20 × 7–9 | 20–21 × 15–16 |
| Species | Zanclea divergens | Zanclea cf. protecta | Zanclea sp. 1 | Zanclea sp. 2 | Zanclea sango | Zanclea sp. (Clade I) | Asyncoryne ryniensis |
|---|---|---|---|---|---|---|---|
| Umbrella | Bell-shaped, diameter: 0.5 mm | Globular to bell-shaped, diameter: 0.5 mm | Globular, diameter: 0.15 mm | Globular, diameter: 0.2 mm | Bell-shaped, diameter: 0.7 mm | Bell-shaped, diameter: 0.5 mm | Bell-shaped, diameter: 0.5 mm |
| Canals | 4 radials, one circular | 4 radials, one circular | No | No | 4 radials, one circular | 4 radials, one circular | 4 radials, one circular |
| Bulbs | 4, 2 tentacular larger | 4, 2 tentacular larger | 2 | 2 | 4, 2 tentacular larger | 4, 2 tentacular larger | 4, 2 tentacular larger |
| Nematocyst pouches | 4, 2 above tentacular bulbs larger | 4, same size | No | No | 4, same size | 4, same size | 4, same size |
| Manubrium | Cylindrical, 1/3 of the subumbrellar cavity | Cylindrical, 1/3 of the subumbrellar cavity | Reaching the velar opening, with 4 arms | Protruding from the bell cavity, with 4 arms | Cylindrical, 1/3 of the subumbrellar cavity | Cylindrical, 1/3 of the subumbrellar cavity | Cylindrical, 1/3 of the subumbrellar cavity |
| Tentacles | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| Cnidophores | 21–31, slightly elongated | 25–37, elongated | 10–15, rounded to elongated | 15–17, rounded | 35–40, rounded | 32–43, rounded | 17–24 |
| Color | Transparent, transparent to white manubrium | Transparent, whitish manubrium | Transparent, orange to white manubrium | Transparent, orange to whitish manubrium | Transparent, whitish manubrium | Transparent, whitish manubrium | Transparent, whitish to orange manubrium |
| Exumbrellar nematocysts (size in µm) | Isorhizae: 5–7 × 5–6 | Basitrichous isorhizae: 5–6 × 4–5 | Macrobasic holotrichous mastigophores: 7–8 × 6–7 | Macrobasic holotrichous mastigophores: 7–9 × 6–8 | No | No | Macrobasic holotrichous euryteles: 6–8 × 6–7 |
| Pouches nematocysts (size in µm) | Stenoteles: 12–16 × 11–13 | Stenoteles: 11–13 × 9–10 | No | No | Macrobasic apotrichous euryteles: 18–19 × 7–9; Stenoteles: 11–12 × 10–11 | Macrobasic apotrichous euryteles: 16–20 × 9–11; Stenoteles: 8–10 × 8–9 | Stenoteles: 28–29 × 24–26 |
| Manubrium nematocysts (size in µm) | Stenoteles: 5–8 × 4–6 | Stenoteles: 6–7 × 5–6 | No | No | Stenoteles: 8–10 × 7–8 | Stenoteles: 7–8 × 5–6 | No |
| Cnidophores nematocysts (size in µm) | Bean-shaped macrobasic holotrichous euryteles: 7–8 × 5–6 | Bean-shaped macrobasic holotrichous euryteles: 7–8 × 5–6 | Bean-shaped macrobasic apotrichous euryteles: 5–6 × 4–6 | Bean-shaped macrobasic apotrichous euryteles: 6–8 × 4–5 | Bean-shaped macrobasic apotrichous euryteles: 7–8 × 4–5 | Bean-shaped macrobasic apotrichous euryteles: 7–8 × 4–5 | Bean-shaped macrobasic euryteles: 7–8 × 6–7 |
| Species | Host/Substrate | Polyp GF | Medusa GF |
|---|---|---|---|
| Zanclea divergens | Celleporaria vermiformis | none | none |
| Zanclea cf. protecta | Parasmittina cf. spondylicola; Schizoporella sp. | none | Subumbrella, manubrium, bulbs |
| Zanclea sp. 1 | Celleporaria pigmentaria | none | none |
| Zanclea sp. 2 | Celleporaria sp. | none | none |
| Zanclea sango | Pavona varians | none | Manubrium (not in the middle), canals, bulbs |
| Zanclea sp. (Clade I) | Goniastrea sp. | none | Manubrium (whole), canals, bulbs |
| Asyncoryne ryniensis | Rock, sponge | base of tentacles | none |
| (1) | (2) | (3) | (4) | (5) | (6) | (7) | |
|---|---|---|---|---|---|---|---|
| (1) Z. divergens | 0 | ||||||
| (2) Z. protecta | 11.7 (1.2) | 0 | |||||
| (3) Zanclea sp. 1 | 10.7 (1.3) | 13.2 (1.4) | 0 | ||||
| (4) Zanclea sp. 2 | 12.5 (1.3) | 13.7 (1.3) | 10.7 (1.2) | 0 | |||
| (5) Z. sango | 12.9 (1.3) | 13.0 (1.3) | 14.9 (1.4) | 14.9 (1.4) | 0 | ||
| (6) Zanclea sp. (Clade I) | 12.0 (1.3) | 12.0 (1.3) | 14.0 (1.4) | 14.0 (1.3) | 4.0 (0.8) | 0 | |
| (7) A. ryniensis | 12.7 (1.3) | 11.5 (1.3) | 13.2 (1.4) | 12.9 (1.3) | 13.7 (1.4) | 12.3 (1.3) | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maggioni, D.; Saponari, L.; Seveso, D.; Galli, P.; Schiavo, A.; Ostrovsky, A.N.; Montano, S. Green Fluorescence Patterns in Closely Related Symbiotic Species of Zanclea (Hydrozoa, Capitata). Diversity 2020, 12, 78. https://doi.org/10.3390/d12020078
Maggioni D, Saponari L, Seveso D, Galli P, Schiavo A, Ostrovsky AN, Montano S. Green Fluorescence Patterns in Closely Related Symbiotic Species of Zanclea (Hydrozoa, Capitata). Diversity. 2020; 12(2):78. https://doi.org/10.3390/d12020078
Chicago/Turabian StyleMaggioni, Davide, Luca Saponari, Davide Seveso, Paolo Galli, Andrea Schiavo, Andrew N. Ostrovsky, and Simone Montano. 2020. "Green Fluorescence Patterns in Closely Related Symbiotic Species of Zanclea (Hydrozoa, Capitata)" Diversity 12, no. 2: 78. https://doi.org/10.3390/d12020078
APA StyleMaggioni, D., Saponari, L., Seveso, D., Galli, P., Schiavo, A., Ostrovsky, A. N., & Montano, S. (2020). Green Fluorescence Patterns in Closely Related Symbiotic Species of Zanclea (Hydrozoa, Capitata). Diversity, 12(2), 78. https://doi.org/10.3390/d12020078