Offshore Neopycnodonte Oyster Reefs in the Mediterranean Sea



Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. General Features of Neopycnodonte cochlear Reefs
3.2. A Glimpse at Reefs’ Associated Biodiversity
4. Discussion
5. Conclusions
Supplementary Materials
Supplementary File 1Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Galtsoff, P.S. The American oyster, Crassostrea virginica Gmelin. Fish. Bull. 1964, 64, 1–488. [Google Scholar]
- Stenzel, H.B. Oyster. In Treatise on Invertebrate Paleontology; Part N, Volume 3, Mollusca 6, Bivalvia; Moore, R.C., Ed.; Geological Society of America and University of Kansas. Boulder & Lawrence: Lawrence, MA, USA, 1971; pp. 953–1224. [Google Scholar]
- Lenihan, H.S.; Peterson, C.H. How habitat degradation through fishery disturbance enhances impacts of hypoxia on oyster reefs. Ecol. Appl. 1998, 8, 28–140. [Google Scholar] [CrossRef]
- Gutierrez, A.P.; Turner, F.; Gharbi, K.; Talbot, R.; Lowe, N.R.; Peñaloza, C.; McCullough, M.; Prodöhl, P.A.; Bean, T.P.; Houston, R.D. Development 470 of a Medium Density Combined-Species SNP Array for Pacific and European oysters (Crassostrea gigas and Ostrea edulis). G3 Genes Genomes Genet. 2017, 7, 2209–2218. [Google Scholar]
- Piazza, B.P.; Bank, P.D.; La Peyre, M.K. The potential for created oyster shell reefs as a sustainable shoreline protection strategy in Louisiana. Restor. Ecol. 2005, 13, 499–506. [Google Scholar] [CrossRef]
- Ruesink, J.L.; Lenihan, H.S.; Trimble, A.C.; Heiman, K.W.; Micheli, F.; Byers, J.E.; Kay, M.C. Introduction of non-native oysters: Ecosystem effects and restoration implications. Ann. Rev. Ecol. Evol. Syst. 2005, 36, 643–689. [Google Scholar] [CrossRef] [Green Version]
- Tolley, S.G.; Volety, A.K. The role of oysters in habitat use of oyster reefs by resident fishes and decapod crustaceans. J. Shellfish Res. 2005, 24, 1007–1012. [Google Scholar]
- Kochmann, J.; Buschbaum, C.; Volkenborn, N.; Reise, K. Shift from native mussels to alien oysters: Differential effects of ecosystem engineers. J. Exp. Mar. Biol. Ecol. 2008, 364, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Markert, A.; Wehrmann, A.; Kroncke, I. Recently established Crassostrea-reefs versus native Mytilus-beds: Differences in ecosystem engineering affects the macrofaunal communities (Wadden Sea of Lower Saxony, southern German Bight). Biol. Invasions 2010, 12, 15–32. [Google Scholar] [CrossRef]
- Grabowski, J.H.; Brumbaugh, R.D.; Conrad, R.F.; Keeler, A.G.; Opaluch, J.J.; Peterson, C.H.; Piehler, M.F.; Powers, S.P.; Smyth, A.R. Economic Valuation of ecosystem services provided by oyster reefs. BioScience 2012, 62, 900–909. [Google Scholar] [CrossRef] [Green Version]
- Walles, B.; de Paiva, J.S.; van Prooijen, B.C.; Ysebaer, T.; Smaal, A.C. The ecosystem engineer Crassostrea gigas affects tidal flat morphology beyond the boundary of their reef structures. Estuaries Coasts 2015, 38, 941–950. [Google Scholar] [CrossRef]
- Beck, M.W.; Brumbaugh, R.D.; Airoldi, L.; Carranza, A.; Coen, L.D.; Crawford, C.; Defeo, O.; Edgar, G.J.; Hancock, B.; Kay, M.C.; et al. Oyster reefs at risk and recommendations for conservation, restoration, and management. BioScience 2011, 61, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Bahr, L.N.; Lanier, W.P. The Ecology of Intertidal Oyster Reefs of the South Atlantic Coast: A Community Profile; U.S. Fish and Wildlife Service, Office of Biological Services: Washington, DC, USA, 1981. [Google Scholar]
- Drinkward, A.C. Introductions and developments of oysters in the North Sea area: A review. Helgol. Meeres. 1999, 52, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Bayne, B. Biology of Oysters; Academic Press: London, UK, 2017; p. 860. [Google Scholar]
- Guo, X.; Cui, L.; Wang, H.; Xu, Z. Diversity and evolution of living oysters. J. Shellfish Res. 2018, 37, 755–771. [Google Scholar] [CrossRef]
- Fürsich, F.T. Palaeoecology and evolution of Mesozoic salinity-controlled benthic macroinvertebrate associations. Lethaia 1993, 26, 327–346. [Google Scholar] [CrossRef]
- Geyer, G.; Herbig, H.G. New Eocene oysters and the final regression at the southern rim of the central high Atlas (Morocco). Geobios 1988, 21, 663–691. [Google Scholar] [CrossRef]
- Seilacher, A.; Matyja, B.A.; Wierzbowski, A. Oyster beds: Morphologic response to changing substrate conditions. In Sedimentary and Evolutionary Cycles, Lect. Notes Earth Scie; Book Series (LNEARTH, Volume 1); Springer: Berlin/Heidelberg, Germany, 2005; pp. 421–435. [Google Scholar]
- Machalski, M. Oyster life positions and shell beds from the Upper Jurassic of Poland. Acta Paleontol. Pol. 1998, 43, 609–634. [Google Scholar]
- Beu, A.G.; Raine, J.I. Revised Descriptions of New Zealand Cenozoic Mollusca from Beu and Maxwell; GNS Science Miscellaneous Series no. 27; Publications Officer, GNS Science: Lower Hutt, New Zealand, 2009; Available online: https://www.gns.cri.nz/static/Mollusca/ (accessed on 29 December 2019).
- MacMillan, L.C. Comparative Sedimentology and Paleoecology of Fossil Giant Oyster Beds in Some Tertiary Strata of New Zealand and Argentina. Master’s Thesis, The University of Waikato, Hamilton, New Zealand, 2010. Available online: http://waikato.researchgateway.ac.nz/ (accessed on 20 December 2019).
- Harzhauser, M.; Djuricic, A.; Mandic, O.; Neubauer, T.A.; Zuschin, M.; Pfeifer, N. Age structure, carbonate production and shell loss rate in an Early Miocene reef of the giant oyster Crassostrea gryphoides. Biogeosciences 2016, 13, 1223–1235. [Google Scholar] [CrossRef] [Green Version]
- Pufahl, P.K.; James, N.P. Monospecific Pliocene oyster buildups, Murray Basin, South Australia: brackish water end member of the reef spectrum. Palaeogeogr. Palaeoclim. Palaeoecol. 2006, 233, 11–33. [Google Scholar] [CrossRef]
- Royal Commission. Report of the Commissioners: Trawl Net and Beam Trawl Fishing with Minutes of Evidence and Appendix; Eyre and Spottiswoode: London, UK, 1885. [Google Scholar]
- Pérès, J.M.; Picard, J. Nouveau manuel de bionomie benthique. Rec. Trav. Stat. Mar. Endoume. 1964, 31, 1–137. [Google Scholar]
- Parenzan, P. Storia della Crassostrea gigas (Thumb.) spontanea nel Mediterraneo. Thalass. Salent. 1969, 19, 39–48. [Google Scholar]
- Poluzzi, A.; Sabelli, B.; Taviani, M. Autosinecologia dei molluschi dei fondi mobili del Delta settentrionale del Po (estate 1980). Boll. Soc. Paleontol. Ital. 1981, 21, 169–178. [Google Scholar]
- FAO. Cultured Aquatic Species Information Programme. Crassostrea Gigas. Available online: http://www.fao.org/fishery/culturedspecies/Crassostrea_gigas/en (accessed on 31 December 2019).
- Smaal, A.C.; Kater, B.J.; Wijsman, J. Introduction, establishment and expansion of the Pacific oyster Crassostrea gigas in the Oosterschelde (SW Netherlands). Helgol. Mar. Res. 2009, 63, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Todorova, V.; Micu, D.; Klisurov, L. Unique oyster reef discovered in the Bulgarian Black Sea. CR. Acad. Bulg. Sci. 2009, 62, 871–874. [Google Scholar]
- Troost, T.K. Causes and effects of a highly successful marine invasion: Case-study of the introduced Pacific oyster Crassostrea gigas in continental NW European estuaries. J. Sea Res. 2010, 64, 145–165. [Google Scholar] [CrossRef]
- Wrange, A.-L.; Valero, J.; Harkestad, L.S.; Strand, O.; Lindegarth, S.; Christensen, H.T.; Dolmer, P.; Kristensen, P.S.; Mortensen, S. Massive settlements of the Pacific oyster, Crassostrea gigas, in Scandinavia. Biol. Inv. 2010, 12, 1145–1152. [Google Scholar] [CrossRef]
- Lejart, M.; Hilly, C. Differential response of benthic macrofauna to the formation of novel oyster reefs (Crassostrea gigas, Thunberg) on soft and rocky substrate in the intertidal of the Bay of Brest, France. J. Sea Res. 2011, 65, 84–93. [Google Scholar] [CrossRef]
- Taviani, M.; Roveri, M.; Impiccini, R.; Vigliotti, L. Segnalazione di Quaternario marino nella Val Chero (Appennino piacentino). Boll. Soc. Paleont. Ital. 1997, 36, 331–338. [Google Scholar]
- Dominici, S.; Benvenuti, M.; Forli, M.; Bogi, C.; Guerrini, A. Upper Miocene molluscs of Monti Livornesi (Tuscany, Italy): Biotic changes across environmental gradients. Palaeogeogr. Palaeoclim. Palaeoecol. 2019, 527, 103–117. [Google Scholar] [CrossRef]
- Ceregato, A.; Raffi, S.; Scarponi, D. The circalittoral/bathyal paleocommunities in the Middle Pliocene of Northern Italy: The case of the Korobkovia oblonga–Jupiteria concava paleocommunity type. Geobios 2007, 40, 555–572. [Google Scholar] [CrossRef]
- Studencka, B.; Jasionowski, M. Bivalves from Middle Miocene reefs of Poland and Ukraine: A new approach to Badenian/Sarmatian boundary in the Paratethys. Acta Geol. Pol. 2011, 61, 79–114. [Google Scholar]
- Górka, M.; Studencka, B.; Jasionowski, M.; Hara, U.; Wysocka, A.; Poberezhskyy, A. The Medobory Hills (Ukraine): Middle Miocene reef systems in the Paratethys, their biological diversity and lithofacies. Biul. Państ. In. Geol. 2012, 449, 147–174. [Google Scholar]
- Freiwald, A.; Beuck, L.; Rüggeberg, A.; Taviani, M.; Hebbeln, D.; R/V METEOR Cruise M70-1 Participants. The white coral community in the central Mediterranean revealed by ROV surveys. Oceanography 2009, 22, 58–74. [Google Scholar] [CrossRef] [Green Version]
- Wisshak, M.; López Correa, M.; Gofas, S.; Salas, C.; Taviani, M.; Jakobsen, J.; Freiwald, A. Shell architecture, element composition, and stable isotope signature of the giant deep-sea oyster Neopycnodonte zibrowii sp. n. from the NE Atlantic. Deep Sea Res. Part I 2009, 56, 374–407. [Google Scholar] [CrossRef]
- Gofas, S.; Rueda, J.L.; Salas, C.; Díaz-del-Río, V. A new record of the giant deep-sea oyster Neopycnodonte zibrowii in the Gulf of Cadiz (south-western Iberian Peninsula). Mar. Biodiv. Rec. 2010, 3, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Van Rooij, D.; DeMol, L.; LeGuilloux, E.; Wisshak, M.; Huvenne, V.A.I.; Moeremans, R.; Henriet, J.-P. Environmental setting of deep-water oysters in the Bay of Biscay. Deep Sea Res. Part I 2010, 57, 1561–1572. [Google Scholar] [CrossRef] [Green Version]
- Beuck, L.; Aguilar, R.; Fabri, M.-C.; Freiwald, A.; Gofas, S.; Hebbeln, D.; López Correa, M.; Ramos, A.; Ramil, F.; Sánchez, F.; et al. Biotope characterisation and compiled geographical distribution of the deep-water oyster Neopycnodonte zibrowii in the Atlantic Ocean and Mediterranean Sea. Rapp. Comm. Intern. Mer Médit. 2016, 41, 462. [Google Scholar]
- Aguilar, R.; Pilar, M.; Gerovasileiou, V. Draft Guidelines for Inventoring and Monitoring of Dark Habitats; UNEP(DEPI)/MED WG. 431/Inf.12, Thirteenth Meeting of Focal Points for Specially Protected Areas. Agenda Item 4: Progress report on activities carried out by SPA/RAC since the twelfth meeting of Focal Points for SPAs. United Nations Environment Programme/Mediterranean Action Plan (UN Environment /MAP); Specially Protected Areas Regional Activity Centre (SPA/RAC): Alexandria, Egypt, 2017; pp. 1–55. [Google Scholar]
- Fourt, M.; Goujard, A.; Perez, T.; Chevaldonné, P. Guide de la faune profonde de la mer Méditerranée. Exploration des roches et canyons sous-marins des côtes françaises; Muséum National d’Histoire Naturelle: Paris, France, 2017; pp. 1–184. [Google Scholar]
- Taviani, M.; Angeletti, L.; Cardone, F.; Montagna, P.; Danovaro, R. A unique and threatened deep water coral-bivalve biotope new to the Mediterranean Sea offshore the Naples megalopolis. Sci. Rep. 2019, 9, 12. [Google Scholar] [CrossRef] [Green Version]
- Gofas, A.; Freiwald, A.; López Correa, M.; Remia, A.; Salas, C.C.; Taviani, M.; Wisshak, M.; Zibrowius, H. Oyster beds in the deep sea. In Abstracts World Congress of Malacology; Jordaens, K., Van Houtte, N., Van Goethem, J., Backeljau, T., Eds.; Unitas Malacologica: Antwerp, Belgium, 2007; pp. 80–82. [Google Scholar]
- Le Danois, E. Les Profondeurs De La Mer; Payot: Paris, France, 1948; pp. 1–303. [Google Scholar]
- Onorato, R.; Forti, P.; Belmonte, G.; Poto, M.; Costantini, A. La grotta sottomarina Lu Lampiune: novità esplorative. Thalassia Salent. 2003, 26, 55–64. [Google Scholar]
- Taviani, M.; Angeletti, L.; Campiani, E.; Ceregato, A.; Foglini, F.; Maselli, V.; Morsilli, M.; Parise, M.; Trincardi, F. Drowned karst landscape offshore the Apulian margin (Southern Adriatic Sea, Italy). J. Cave Karst Stud. 2012, 74, 197–212. [Google Scholar] [CrossRef]
- Corriero, G.; Pierri, C.; Mercurio, M.; Marzano, C.N.; Tarantini, S.O.; Gravina, M.F.; Lisco, S.; Moretti, M.; De Giosa, F.; Valenzano, E.; et al. Mediterranean mesophotic coral reef built by non-symbiotic scleractinians. Sci. Rep. 2019, 9, 3601. [Google Scholar] [CrossRef] [PubMed]
- Sotomayor-García, A.; Rueda, J.L.; Sánchez-Guillamón, O.; Urra, J.; Vázquez, J.T.; Palomino, D.; Fernández-Salas, L.M.; López-González, N.; González-Porto, M.; Santana-Casiano, J.M.; et al. First macro-colonizers and survivors around Tagoro Submarine Volcano, Canary Islands, Spain. Geosciences 2019, 9, 52. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.B.; Fodire, J.F.; Ridge, J.T.; Lindquist, N.L.; Theuerkauf, E.J.; Coleman, S.E.; Grabowski, J.H.; Brodeur, M.C.; Gittman, R.K.; Keller, D.A.; et al. Oyster reefs can outpace sea-level rise. Nat. Clim. Chang. 2014, 4, 493–497. [Google Scholar] [CrossRef]
- Peters, J.W.; Eggleston, D.B.; Puckett, B.J.; Theuerkauf, S.J. Oyster demographics in harvested reefs vs. no-take reserves: Implications for larval spillover and restoration success. Front. Mar. Sci. 2017, 4, 326. [Google Scholar] [CrossRef] [Green Version]
- Ridge, J.T.; Rodriguez, A.B.; Fodrie, F.J. Evidence of exceptional oyster-reef resilience to fluctuations in sea level. Ecol. Evol. 2019, 7, 10409–10420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Southwell, M.W.; Veenstra, J.J.; Adams, C.D.; Scarlett, E.V.; Payne, K.B. Changes in sediment characteristics upon oyster reef restoration, NE Florida, USA. J. Coast. Zone Manag. 2017, 20, 442. [Google Scholar] [CrossRef]
- Smaal, A.C.; Ferreira, J.G.; Grant, J.; Petersen, J.K.; Strand, Ø. Goods and Services of Marine Bivalves; Springer International Publishing, Springer Nature Switzerland AG: Cham, Switzerland, 2019; pp. 1–598. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Interpretation Manual of European Union Habitats; EUR 28; European Commission, DG Environment, Nature ENV B.3 2013. p. 144. Available online: http://ec.europa.eu/environment/nature/legislation/habitatsdirective/docs/Int_Manual_EU28.pdf (accessed on 5 December 2019).
- La Peyre, M.K.; Marshall, D.A.; Miller, L.S.; Humpries, A.T. Oyster reefs in Northern Gulf of Mexico estuaries harbor diverse fish and decapod crustacean assemblages: a meta-synthesis. Front. Mar. Sci. 2019, 6, 666. [Google Scholar] [CrossRef] [Green Version]
- EUNIS. 1997. Available online: https://eunis.eea.europa.eu/habitats-code-browser.jsp (accessed on 28 December 2019).
- EMODnet Bathymetry Consortium: EMODnet Digital Bathymetry (DTM). 2018. Available online: http://www.emodnet-bathymetry.eu (accessed on 20 December 2019).
- Foglini, F.; Grande, V.; Marchese, F.; Bracchi, V.A.; Prampolini, M.; Angeletti, L.; Castellan, G.; Chimienti, G.; Hansen, I.M.; Gudmunsen, M.; et al. Application of hyperspectral imaging to underwater habitat mapping, Southern Adriatic Sea. Sensors 2019, 19, 2261. [Google Scholar] [CrossRef] [Green Version]
- Santín, A.; Grynó, J.; Ambroso, S.; Uriz, M.J.; Dominguez-Carrió, C.; Gili, J.-M. Distribution patterns and demographic trends of demosponges at the Menorca Channel (Northwestern Mediterranean Sea). Prog. Oceanogr. 2019, 173, 9–25. [Google Scholar]
- WoRMS Editorial Board. 2019. Available online: http://www.marinespecies.org (accessed on 25 December 2019).
- Bortoluzzi, G.; Gasperini, L.; Cagatay, N.; Gorur, N. Geophysical and Geological Studies in the Sea of Marmara and Gulf of Saros. ISMAR Bologna Technical Report, 98; Bologna, Italy, 2005; pp. 1–33. Available online: http://ricerca.ismar.cnr.it/CRUISE_REPORTS/2000-2009/MARM05_REP/MARM05_REP.pdf (accessed on 20 December 2019).
- Taviani, M.; Franchi, F.; Angeletti, L.; Correggiari, A.; López Correa, M.; Maselli, V.; Mazzoli, C.; Peckmann, J. Biodetrital carbonates on the Adriatic continental shelf imprinted by oxidation of seeping hydrocarbons. Mar. Petrol. Geol. 2015, 66, 511–531. [Google Scholar] [CrossRef]
- Harding, J.M.; Mann, R. Oyster reefs as fish habitat: opportunistic use of restored reefs by transient fishes. J. Shelf Res. 2001, 20, 951–959. [Google Scholar]
- Ingrosso, G.; Abbiati, M.; Badalamenti, F.; Bavestrello, G.; Belmonte, G.; Cannas, R.; Benedetti-Cecchi, L.; Bertolino, M.; Bevilacqua, S.; Bianchi, C.N.; et al. Mediterranean bioconstructions along the Italian coast. Adv. Mar. Biol. 2018, 79, 61–136. [Google Scholar] [PubMed]
- Carlucci, R.; Lembo, G.; Maiorano, P.; Capezzuto, F.; Marano, C.A.; Sion, L.; Spedicato, M.T.; Ungaro, N.; Tursi, A.; D’Onghia, G. Nursery areas of red mullet (Mullus barbatus), hake (Merluccius merluccius) and deep-water rose shrimp (Parapenaeus longirostris) in Eastern-Central Mediterranean Sea. Estuar. Coast. Shelf Sci. 2009, 83, 529–538. [Google Scholar] [CrossRef]
- FAO. The State of Mediterranean and Black Sea Fisheries; General Fisheries Commission for the Mediterranean: Rome, Italy, 2018; pp. 1–172. Available online: http://www.fao.org/3/ca2702en/CA2702EN.pdf (accessed on 5 February 2020).
- Spedicato, M.T.; Zupa, W.; Carbonara, P.; Casciaro, L.; Bitetto, I.; Facchini, M.T.; Gaudio, P.; Palmisano, M.; Lembo, G. Lo stato delle risorse biologiche e della pesca nel Basso Adriatico e nello Ionio nord occidentale. In Atti del Convegno il Mare Adriatico e le sue Risorse; Marini, M., Bombace, G., Iacobone, G., Eds.; Carlo Saladino Editore: Palermo, Italy, 2017; pp. 177–208. [Google Scholar]
- Council Directive 92/43/EEC—EUR-Lex. Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:1992:206:0007:0050:EN:PDF (accessed on 5 December 2019).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
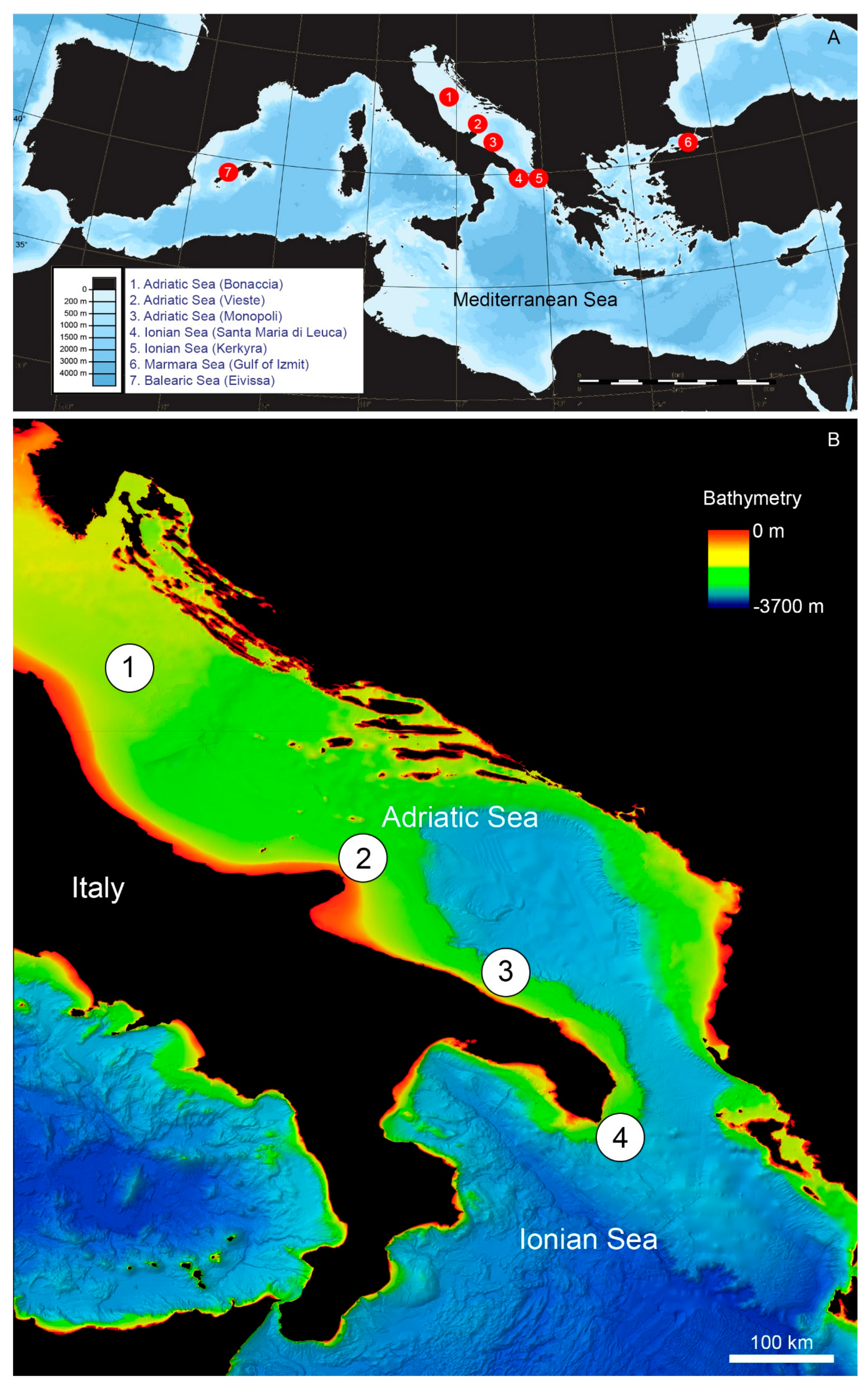
| Site | Collection Date | Station | Lat N-Long E Start | Lat N-Long E End | Depth Min–Max (m) | Dive Length (m) |
|---|---|---|---|---|---|---|
| Bonaccia | 13 August 2017 | MS17_III_110 | 43°35.50′-14°20.11′ | 43°35.23′-14°20.44′ | 77–83 | 1079.93 |
| Vieste | 10 November 2015 | MS15_47 | 41°59,62′-16°15,14′ | 41°59,68′-16°15,13′ | 55–75 | 804.72 |
| Vieste | 10 November 2015 | MS15_48 | 42°00.81′-16°11.13′ | 42°00.78′-16°11.13′ | 48–49 | 58.70 |
| Vieste | 11 November 2015 | MS15_62 | 42°01.94′-16°10.24′ | 42°01.84′-16°10.25′ | 60–66 | 139.52 |
| Vieste | 11 November 2015 | MS15_63 | 42°01.02′-16°11.75′ | 42°00.95′-16°11.79′ | 50–52 | 316.75 |
| Vieste | 11 November 2015 | MS15_67 | 41°59.60′-16°15.14′ | 41°59.48′-16°15.23′ | 52–72 | 255.95 |
| Monopoli | 5 August 2017 | MS17_II_180 | 41°00.19′-17°24.25′ | 40°59.85′-17°24.89′ | 95–102 | 1685.84 |
| Monopoli | 15 August 2017 | MS17_III_115 | 41°04.17′-17°18.15′ | 41°04.38′-17°18.24′ | 85–87 | 1039.10 |
| Santa Maria di Leuca | 31 July 2017 | MS17_II_115 | 39°44.02′-18°22.26′ | 39°44.27′-18°22.12′ | 70–113 | 1179.90 |
| Santa Maria di Leuca | 31 July 2017 | MS17_II_117 | 39°42.31′-18°21.31′ | 39°42.48′-18°21.68′ | 90–125 | 1200.00 |
| Kerkyra Island | 3 May 2006 | CR83 (Epibenthic haul) | 39°46.00′-19°25.52′ | 39°47.22′-19°25.42′ | 77–99 | n/a |
| Gulf of Izmit | 19 September 2005 | GRA02 (Grab) | 40°44.00′-29°27.00′ | - | 56 | n/a |
| Eivissa Island | 15 April 2004 | COBAS-78 (Epibenthic haul) | 38°45.99′-01°18.05′ | 38°46.74′-01°19.34′ | 94–96 | n/a |
| n. | Phylum | Class | Taxon | Auctores | Protection |
|---|---|---|---|---|---|
| 1 | Porifera | Demospongiae | spp. | ||
| 2 | Aplysina aerophoba | (Nardo, 1833) | 1 | ||
| 3 | Axinella cannabina | (Esper, 1794) | 1, 2 (EN) | ||
| 4 | Axinella polypoides | Schmidt, 1862 | 1, 2 (EN), 6 | ||
| 5 | Cliona celata | Grant, 1826 | |||
| 6 | Haliclona mediterranea | Griessinger, 1971 | |||
| 7 | Hexadella racovitzai | Topsent, 1896 | |||
| 8 | Ircinia variabilis | Schmidt, 1862 | |||
| 9 | Mycale tunicata | Schmidt, 1862 | |||
| 10 | cf. Petrosia sp. | ||||
| 11 | Poecillastra compressa | (Bowerbanck, 1866) | 2 (VU) | ||
| 12 | Spongia spp. | ||||
| 13 | cf. Spongia agaricina | Pallas, 1766 | 1, 6 | ||
| 14 | Spongia lamella | (Schulze, 1879) | 2 (EN) | ||
| 15 | Spongia officinalis | Linnaeus, 1759 | 1, 2 (EN), 6 | ||
| 16 | Suberites domuncula | (Olivi, 1792) | |||
| 17 | Tethya aurantium | (Pallas, 1766) | |||
| 18 | Tethya cf. citrina | Sarà and Melone, 1965 | 1 | ||
| 19 | Ulosa stuposa | (Esper, 1794) | |||
| 20 | Cnidaria | Hydrozoa | cf. Halecium sp. | ||
| 21 | cf. Halecium halecinum | (Linnaeus, 1758) | |||
| 22 | Lytocarpia myriophyllum | (Linnaeus, 1758) | |||
| 23 | cf. Sertularella sp. | ||||
| 24 | Anthozoa | Alcyonium palmatum | Pallas, 1766 | ||
| 25 | Caryophyllidae spp. | ||||
| 26 | Caryophyllia cf. smithii | Stokes and Broderip, 1828 | 4 | ||
| 27 | Dendrophyllia cornigera | (Lamarck, 1816) | 2 (VU), 3 (EN), 4 | ||
| 28 | Paralcyonium spinulosum | (Delle Chiaje, 1822) | |||
| 29 | Phyllangia americana | Milne Edwards and Haime, 1849 | 4 | ||
| 30 | Virgularia mirabilis | (Müller, 1776) | 2 (VU), | ||
| 31 | Annelida | Polychaeta | Bonellia viridis | Rolando, 1822 | |
| 32 | Filograna-Salmacina complex | ||||
| 33 | Protula tubularia | (Montagu, 1803) | |||
| 34 | Sabella spallanzanii | (Gmelin, 1791) | |||
| 35 | Serpula vermicularis | Linnaeus, 1767 | |||
| 36 | Terebellides stroemii | Sars, 1835 | |||
| 37 | Vermiliopsis sp. | ||||
| 38 | Mollusca | Bivalvia | Atrina fragilis | (Pennant, 1777) | |
| 39 | Neopycnodonte cochlear | (Poli, 1795) | |||
| 40 | Pecten jacobaeus | (Linnaeus, 1758) | |||
| 41 | Gastropoda | cf. Calliostoma zizyphinum | (Linnaeus, 1758) | ||
| 42 | cf. Fusinus sp. | ||||
| 43 | Hexaplex trunculus | (Linnaeus, 1758) | |||
| 44 | cf. Naticidae | ||||
| 45 | Nudibranchia | cf. Caloria elegans | (Alder and Hancock, 1845) | ||
| 46 | Flabellina affinis | (Gmelin, 1791) | |||
| 47 | Hypselodoris tricolor | (Cantraine, 1835) | |||
| 48 | Marionia blainvillea | (Risso, 1818) | |||
| 49 | Cephalopoda | cf. Octopus vulgaris | Cuvier, 1797 | ||
| 50 | Teuthida sp. | ||||
| 51 | Arthropoda | Malacostraca | Maja squinado | (Herbst, 1788) | 1, 6 |
| 52 | Munida sp. | ||||
| 53 | Pagurus cf. excavatus | (Herbst, 1791) | |||
| 54 | Palinurus elephas | (Fabricius, 1787) | 1, 3 (VU), 6 | ||
| 55 | Bryozoa | Gymnolaemata | Reteporella grimaldii | (Jullien, 1903) | |
| 56 | Smittina cervicornis | (Pallas, 1766) | |||
| 57 | Echinodermata | Asteroidea | Chaetaster longipes | (Bruzelius, 1805) | |
| 58 | Echinaster sepositus | (Retzius, 1783) | |||
| 59 | Marthasterias glacialis | (Linnaeus, 1758) | |||
| 60 | Peltaster placenta | (Müller and Troschel, 1842) | |||
| 61 | Crinoidea | Antedon mediterranea | (Lamarck, 1816) | ||
| 62 | Echinoidea | Centrostephanus longispinus | (Philippi, 1845) | 1, 5, 6 | |
| 63 | Cidaris cidaris | (Linnaeus, 1758) | |||
| 64 | Echinus melo | Lamarck, 1816 | |||
| 65 | Holothuroidea | Holothuria forskali | Delle Chiaje, 1823 | ||
| 66 | Ophiuroidea | cf. Ophiothrix fragilis | (Abildgaard in Müller, 1789) | ||
| 67 | Chordata | Ascidiacea | sp. | ||
| 68 | Botrylloides sp. | ||||
| 69 | Didemnidae sp. | ||||
| 70 | Diplosoma spongiforme | (Giard, 1872) | |||
| 71 | Halocynthia papillosa | (Linnaeus, 1767) | |||
| 72 | Actinopterygii | Callanthias ruber | (Rafinesque, 1810) | ||
| 73 | Chelidonichthys lastoviza | (Bonnaterre, 1788) | |||
| 74 | Conger conger | (Linnaeus, 1758) | |||
| 75 | Muraena helena | Linnaeus, 1758 | |||
| 76 | Phycis phycis | (Linnaeus, 1766) | |||
| 77 | Serranus cabrilla | (Linnaeus, 1758) | |||
| 78 | Scorpaena scrofa | Linnaeus, 1758 | |||
| 79 | Spicara maena | (Linnaeus, 1758) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angeletti, L.; Taviani, M. Offshore Neopycnodonte Oyster Reefs in the Mediterranean Sea. Diversity 2020, 12, 92. https://doi.org/10.3390/d12030092
Angeletti L, Taviani M. Offshore Neopycnodonte Oyster Reefs in the Mediterranean Sea. Diversity. 2020; 12(3):92. https://doi.org/10.3390/d12030092
Chicago/Turabian StyleAngeletti, Lorenzo, and Marco Taviani. 2020. "Offshore Neopycnodonte Oyster Reefs in the Mediterranean Sea" Diversity 12, no. 3: 92. https://doi.org/10.3390/d12030092
APA StyleAngeletti, L., & Taviani, M. (2020). Offshore Neopycnodonte Oyster Reefs in the Mediterranean Sea. Diversity, 12(3), 92. https://doi.org/10.3390/d12030092