Photosynthetic Picoeukaryotes Diversity in the Underlying Ice Waters of the White Sea, Russia

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
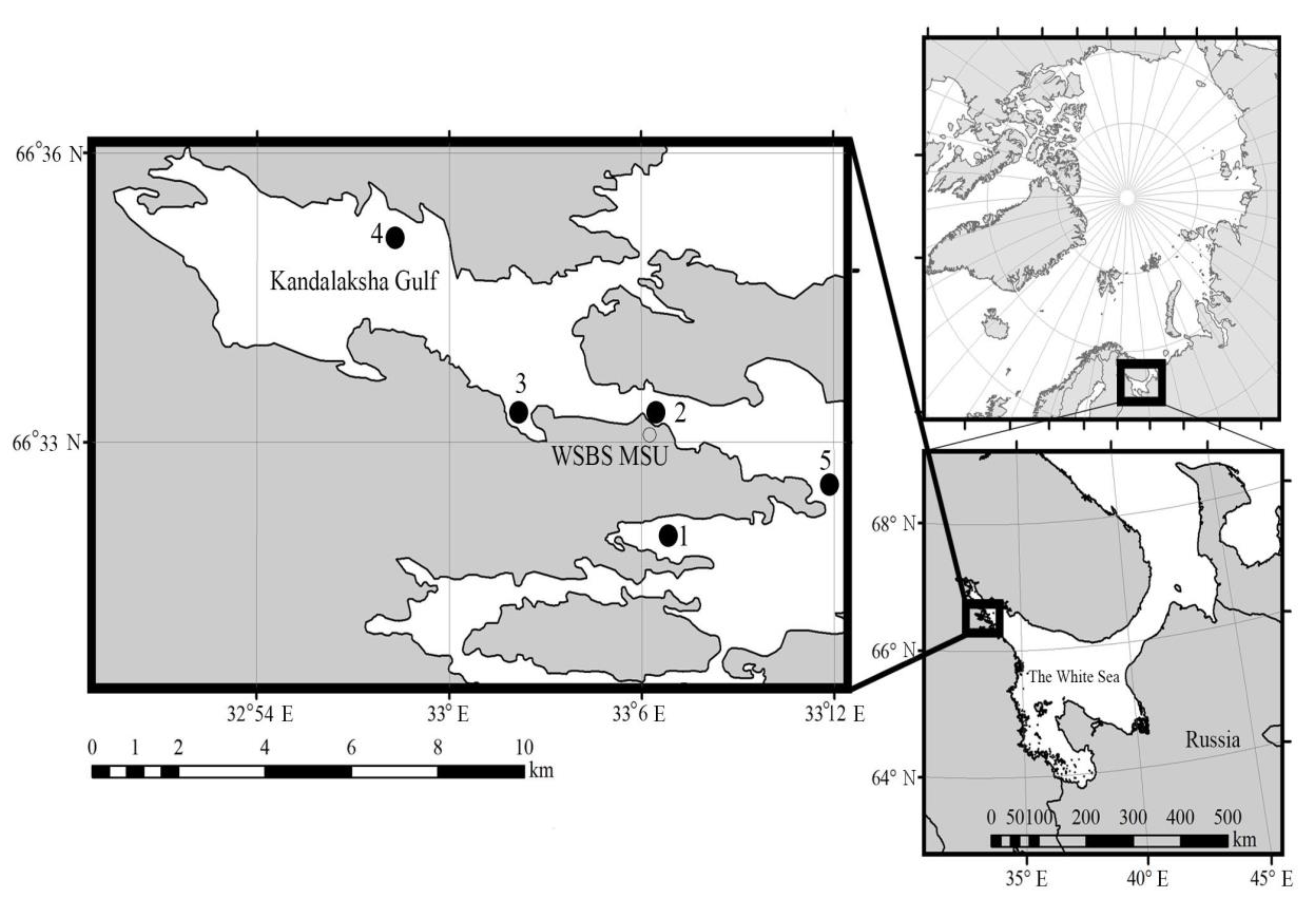
2.1. Sampling and Study Area
2.2. Chlorophylla
2.3. Enumeration of Picophototrophs
2.4. DNA Isolation of Picoplanktonic Size-fraction
2.5. DNA Amplification and Sequencing
2.6. Bioinformatics and Data Evaluation
2.7. Statistical Procedures
3. Results
3.1. Environmental Conditions
3.2. Total Chlorophyll a Biomass
3.3. The Abundance of Picophototrophs
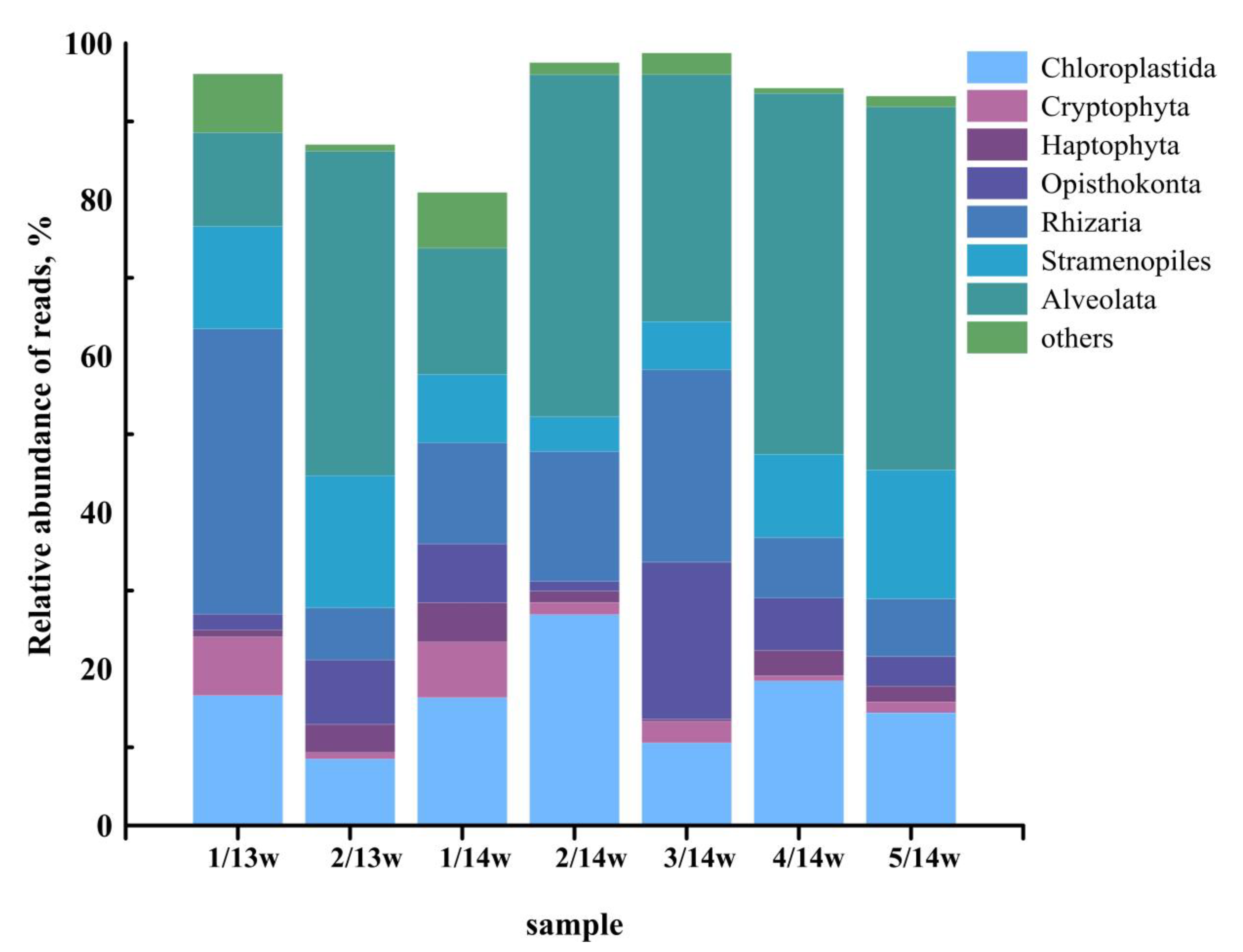
3.4. Taxonomic Composition of Eukaryotes in Samples Filtered through a 2-µm Pore Size Filter
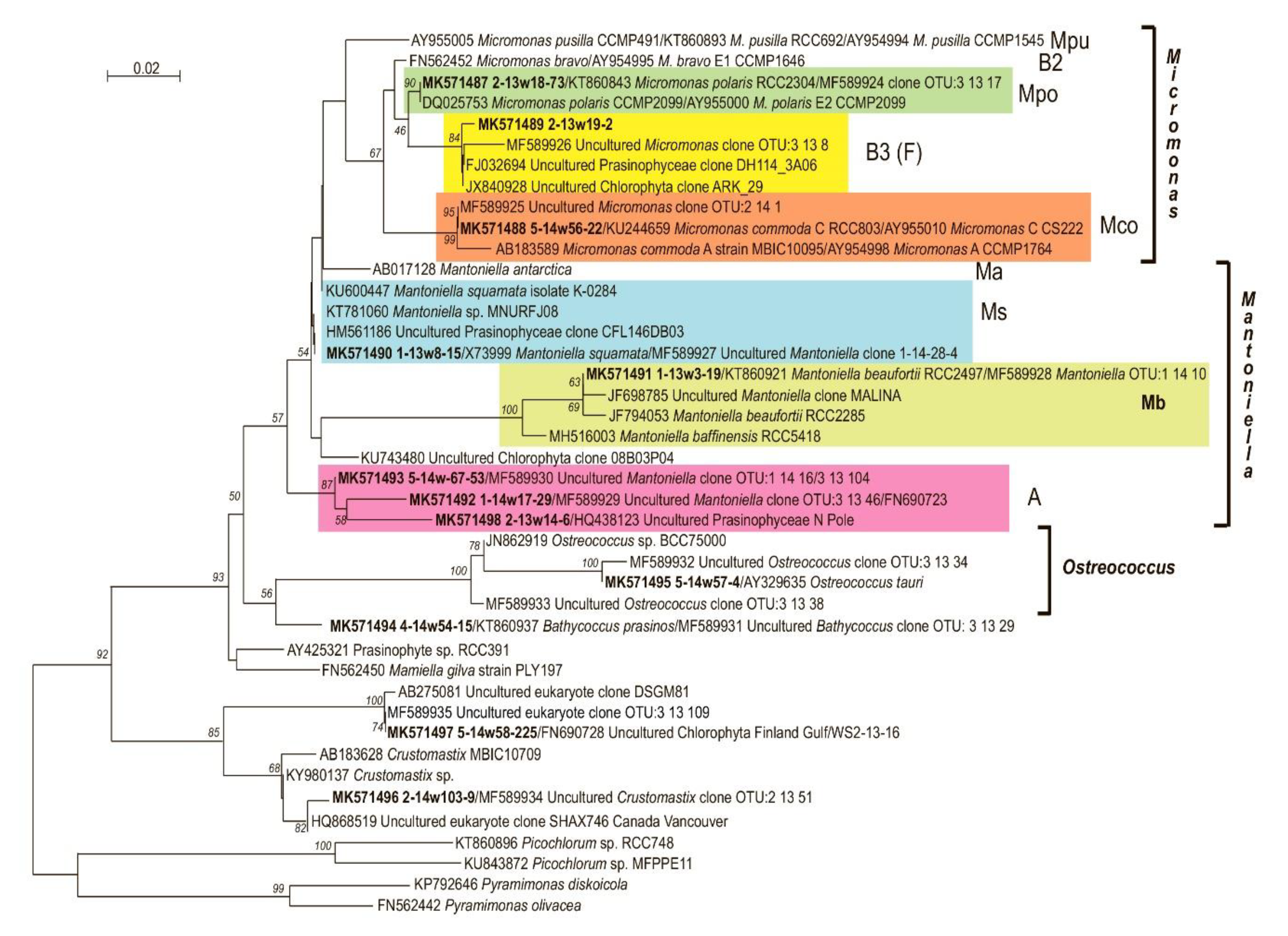
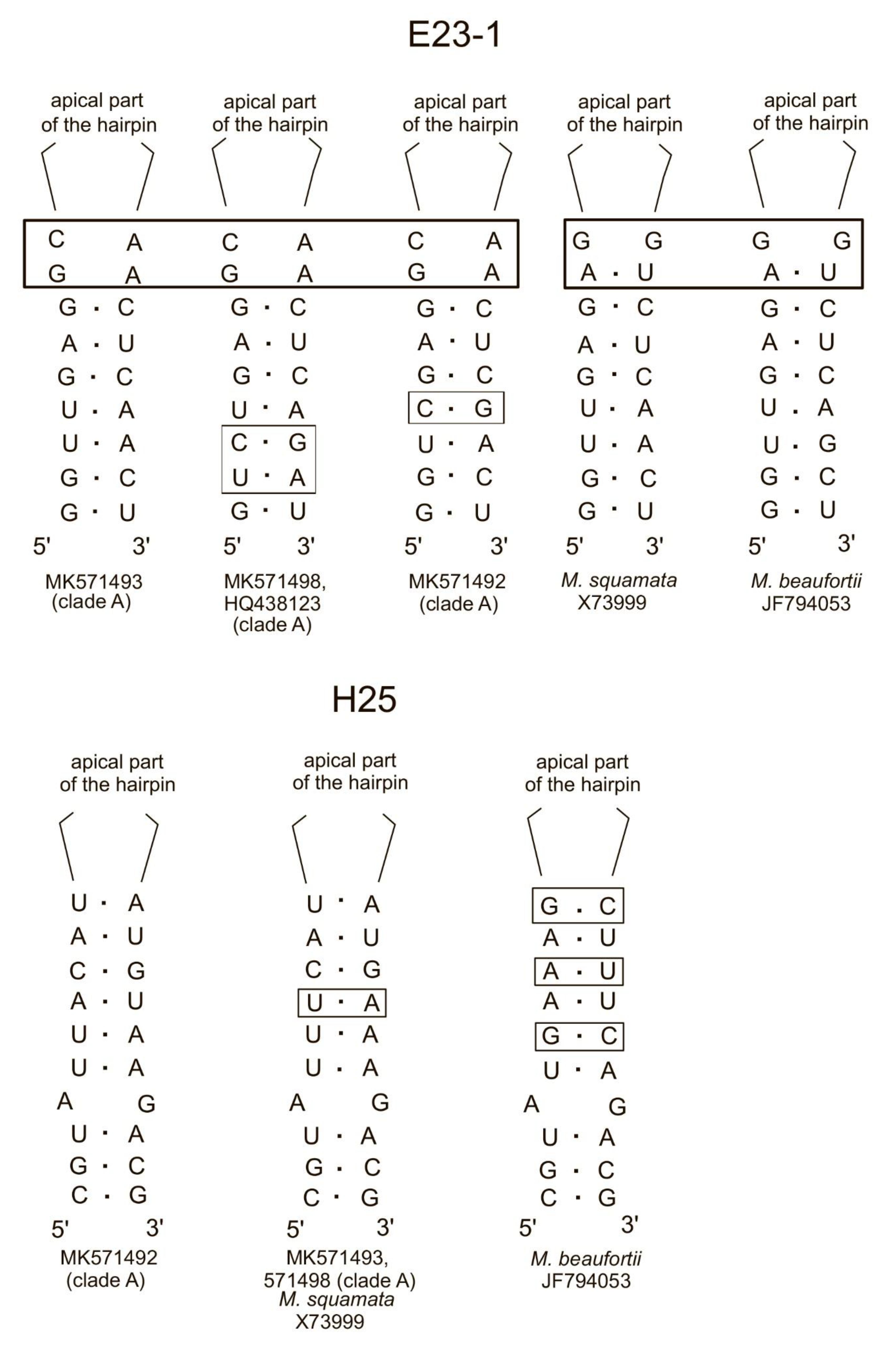
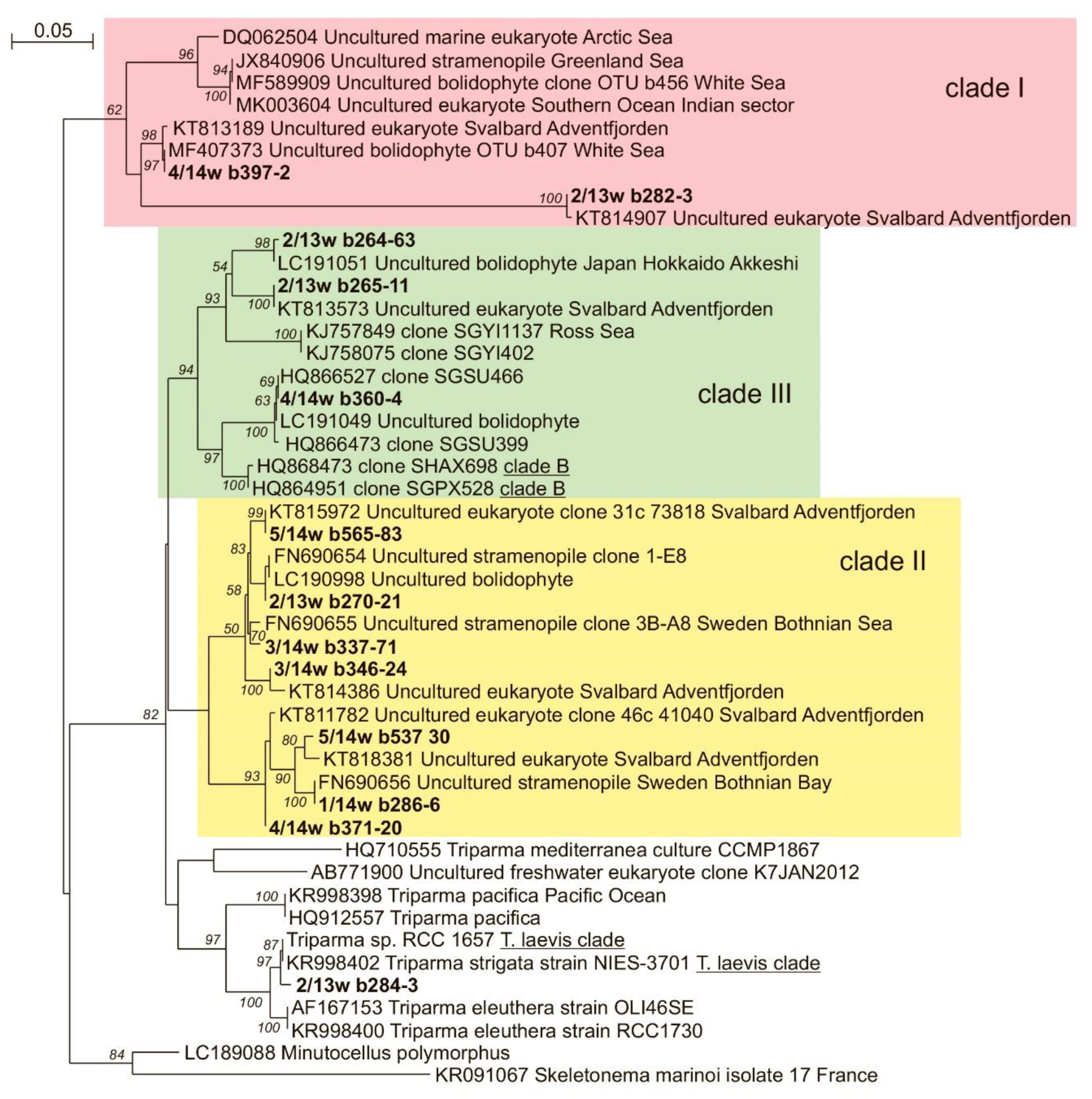
3.5. OTU Richness and Taxonomic Affiliation of the PPEs Sequences
3.6. Chlorophyta
3.7. Stramenopiles
3.8. Haptophyta
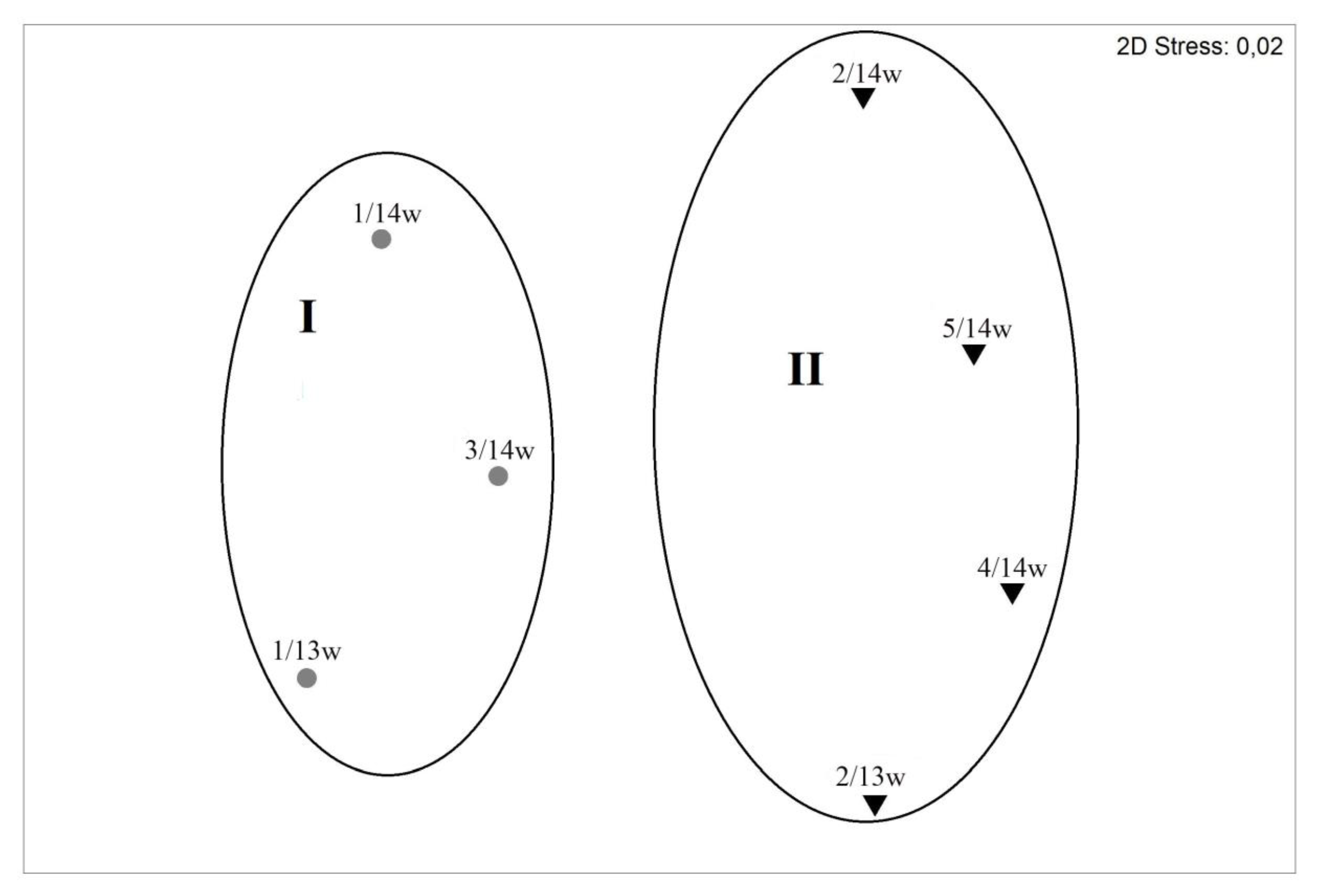
3.9. PPEs Community Structure
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Vaulot, D.; Eikrem, W.; Viprey, M.; Moreau, H. The diversity of small eukaryotic phytoplankton (<3 µm) in marine ecosystems. FEMS Microbiol. Rev. 2008, 32, 795–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massana, R. Eukaryotic Picoplankton in Surface Oceans. Annu. Rev. Microbiol. 2011, 65, 91–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon-van der Staay, S.Y.; De Wachter, R.; Vaulot, D. Oceanic 18S rDNA sequences from picoplankton reveal unsuspected eukaryotic diversity. Nature 2001, 409, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Bhavya, P.A.S.; Lee, J.H.; Lee, H.W.; Kang, J.J.; Lee, J.H.; Lee, D.; An, S.H.; Stockwell, D.A.; Whitledge, T.E.; Lee, S.H. First in situ estimations of small phytoplankton carbon and nitrogen uptake rates in the Kara, Laptev, and East Siberian seas. Biogeosciences 2018, 15, 5503–5517. [Google Scholar] [CrossRef] [Green Version]
- Proshutinsky, A.; Krishfield, R.; Timmermans, M.-L.; Toole, J.; Carmack, E.; McLaughlin, F.; Williams, W.J.; Zimmermann, S.; Itoh, M.; Shimada, K. Beaufort Gyre freshwater reservoir: State and variability from observations. J. Geophys. Res. 2009, 114, 1–25. [Google Scholar] [CrossRef]
- Carmack, E.; Mclaughlin, F. Towards recognition of physical and geochemical change in Subarctic and Arctic Seas. Prog. Oceanogr. 2011, 90, 90–104. [Google Scholar] [CrossRef]
- Li, W.K.W.; McLaughlin, F.A.; Lovejoy, C.; Carmack, E.C. Smallest algae thrive as the Arctic Ocean freshens. Science 2009, 326, 539. [Google Scholar] [CrossRef] [Green Version]
- Vincent, W.F. Microbial ecosystem responses to rapid climate change in the Arctic. ISME J. 2010, 9, 1087–1090. [Google Scholar] [CrossRef] [Green Version]
- Comeau, A.M.; Li, W.K.W.; Tremblay, J.É.; Carmack, E.C.; Lovejoy, C. Arctic ocean microbial community structure before and after the 2007 record sea ice minimum. PLoS ONE 2011, 6, e27492. [Google Scholar] [CrossRef]
- Lovejoy, C.; Massana, R.; Pedros-Alio, C. Diversity and distribution of marine microbial eukaryotes in the Arctic Ocean and adjacent seas. Appl. Environ. Microbiol. 2006, 72, 3085–3095. [Google Scholar] [CrossRef] [Green Version]
- Lovejoy, C.; Vincent, W.F.; Bonilla, S.; Roy, S.; Martineau, M.-J.; Terrado, R.; Potvin, M.; Massana, R.; Pedros-Alio, C. Distribution, phylogeny, and growth of cold-adapted picoprasinophytes in arctic seas. J. Phycol. 2007, 43, 78–89. [Google Scholar] [CrossRef]
- Hamilton, A.K.; Lovejoy, C.; Galand, P.E.; Ingram, R.G. Water masses and biogeography of picoeukaryote assemblages in a cold hydrographically complex system. Limnol. Oceanogr. 2008, 53, 922–935. [Google Scholar] [CrossRef] [Green Version]
- Terrado, R.; Vincent, W.F.; Lovejoy, C. Mesopelagic protists: Diversity and succession in a coastal Arctic ecosystem. Aquat. Microb. Ecol. 2009, 56, 25–39. [Google Scholar] [CrossRef] [Green Version]
- Terrado, R.; Medrinal, E.; Dasilva, C.; Thaler, M.; Vincent, W.; Lovejoy, C. Protist community composition during spring in an Arctic flaw lead polynya. Polar Biol. 2011, 34, 1901–1914. [Google Scholar] [CrossRef]
- Lovejoy, C.; Potvin, M. Microbial eukaryotic distribution in a dynamic Beaufort Sea and the Arctic Ocean. J. Plankton Res. 2011, 33, 431–444. [Google Scholar] [CrossRef] [Green Version]
- Marie, D.; Shi, X.L.; Rigaut-Jalabert, F.; Vaulot, D. Use of flow cytometric sorting to better assess the diversity of small photosynthetic eukaryotes in the English Channel. FEMS Microbiol. Ecol. 2010, 72, 165–178. [Google Scholar] [CrossRef] [Green Version]
- Meshram, A.R.; Vader, A.; Kristiansen, S.; Gabrielsen, T.M. Microbial eukaryotes in an Arctic under-ice spring bloom north of Svalbard. Front. Microbiol. 2017, 8, 1099. [Google Scholar] [CrossRef] [PubMed]
- Kirkham, A.R.; Jardillier, L.E.; Tiganescu, A.; Pearman, J.; Zubkov, M.V.; Scanlan, D.J. Basin-scale distribution patterns of photosynthetic picoeukaryotes along an Atlantic Meridional Transect. Environ. Microbiol. 2011, 13, 975–990. [Google Scholar] [CrossRef]
- Balzano, S.; Marie, D.; Gourvil, P.; Vaulot, D. Composition of the summer photosynthetic pico and nanoplankton communities in the Beaufort Sea assessed by T-RFLP and sequences of the 18S rRNA gene from flow cytometry sorted samples. ISME J. 2012, 6, 1480–1498. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, T.; Yamaguchi, H.; Sato, M.; Watanabe, T.; Taniuchi, Y.; Kuwata, A.; Kawachi, M. Seasonal and geographical distribution of near-surface small photosynthetic eukaryotes in the western North Pacific determined by pyrosequencing of 18S rDNA. FEMS Microbiol. Ecol. 2017, 93, fiw229. [Google Scholar] [CrossRef] [Green Version]
- Diez, B.; Pedros-Alio, C.; Massana, R. Study of genetic diversity of eukaryotic picoplankton in different oceanic regions by small-subunit rRNA gene cloning and sequencing. Appl. Environ. Microbiol. 2001, 67, 2932–2941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metfies, K.; von Appen, W.-J.; Kilias, E.; Nicolaus, A.; Nöthig, E.-M. Biogeography and photosynthetic biomass of Arctic marine picoeukaroytes during summer of the record sea ice minimum 2012. PLoS ONE 2016, 11, e0148512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, V.; Dahle, S.; Galaktionov, K.; Kosobokova, X.; Naumov, A.; Rat’kova, T.; Savinov, V.; Savinova, T. White Sea: Ecology and Environment; Derzavets Publisher: St-Petersburg-Tromsø, Russia, 2001. [Google Scholar]
- Ilyash, L.V.; Belevich, T.A.; Zhitina, L.S.; Radchenko, I.G.; Ratkova, T.N. Phytoplankton of the White Sea. In Biogeochemistry of the Atmosphere, Ice and Water of the White Sea: The White Sea Environment Part I; Lisitzin, A., Gordeev, V., Eds.; Springer: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Belevich, T.A.; Ilyash, L.V.; Milyutina, I.A.; Logacheva, M.D.; Troitsky, A.V. Phototrophic picoeukaryotes of Onega Bay, the White Sea: Abundance and species composition. Mosc. Univ. Biol. Sci. Bull. 2017, 72, 109–114. [Google Scholar] [CrossRef]
- Milyutina, I.A.; Belevich, T.A.; Ilyash, L.V.; Troitsky, A.V. Insight into picophytoplankton diversity of the subarctic White Sea—The first recording of Pedinophyceae in environmental DNA. MicrobiologyOpen 2019, 8, e892. [Google Scholar] [CrossRef] [PubMed]
- Belevich, T.A.; Ilyash, L.V.; Milyutina, I.A.; Logacheva, M.D.; Goryunov, D.V.; Troitsky, A.V. Photosynthetic picoeukaryotes in the land-fast ice of the White Sea, Russia. Microb. Ecol. 2018, 75, 582–597. [Google Scholar] [CrossRef] [PubMed]
- Belevich, T.A.; Ilyash, L.V.; Milyutina, I.A.; Logacheva, M.D.; Goryunov, D.V.; Troitsky, A.V. Metagenomic analyses of White Sea picoalgae: First data. Biochemistry 2015, 80, 1514–1521. [Google Scholar] [CrossRef]
- Pantyulin, A.N. Dynamics, structure, and water masses. In The White Sea System: Water Column and Interacting Atmosphere, Cryosphere, the River Run-Off, and Biosphere; Lisitzin, A.P., Ed.; Scientific World: Moscow, Russia, 2012; Volume 2, pp. 309–379. (In Russian) [Google Scholar]
- Arar, E.J.; Collins, G.B. Method 445.0. In In Vitro Determination of Chlorophyll A and Pheophytin A in Marine and Freshwater Algae by Fluorescence; U.S. Environmental Protection Agency: Washington, DC, USA, 1997. [Google Scholar]
- Hillebrand, H.; Dürselen, C.D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 5, 403–424. [Google Scholar] [CrossRef]
- Verity, P.G.; Robertson, C.Y.; Tronzo, C.R.; Endrews, M.G.; Nelson, J.R.; Sieracki, M.E. Relationship between cell volume and the carbon and nitrogen content of marine photosynthetic nanoplankton. Limnol. Oceanogr. 1992, 37, 1434–1446. [Google Scholar] [CrossRef]
- López-García, P.; Rodríguez-Valera, F.; Pedrós-Alió, C.; Moreira, D. Unexpected diversity of small eukaryotes in deep-sea Antarctic plankton. Nature 2001, 409, 603–607. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nuc. Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Wuyts, J.; De Rijk, P.; De Peer, V.Y.; Pison, G.; Rousseeuw, P.; De Wachter, R. Comparative analysis of more than 3000 sequences reveals the existence of two pseudoknots in area V4 of eukaryotic small subunit ribosomal RNA. Nucleic Acids Res. 2000, 28, 4698–4708. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.L.; Pollina, T.; Gourvil, P.; Corre, E.; Marie, D.; Garrido, J.L.; Rodríguez, F.; Noël, M.-H.; Vaulot, D.; Eikrem, W. Chloropicophyceae, a new class of picophytoplanktonic prasinophytes. Sci. Rep. 2017, 7, 14019. [Google Scholar] [CrossRef] [Green Version]
- Tragin, M.; Vaulot, D. Novel diversity within marine Mamiellophyceae (Chlorophyta) unveiled by metabarcoding. Sci. Rep. 2019, 9, 5190. [Google Scholar] [CrossRef] [Green Version]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. 2017. Available online: http://www.Algaebase.Org (accessed on 1 May 2019).
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; Primer-E Ltd.: Plymouth, UK, 2001. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; Primer-E Ltd.: Plymouth, UK, 2006. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Belevich, T.A.; Ilyash, L.V.; Milyutina, I.A.; Logacheva, M.D.; Troitsky, A.V. Metagenomics of Bolidophyceae in plankton and ice of the White Sea. Biochemistry 2017, 82, 1917–1928. [Google Scholar] [CrossRef] [PubMed]
- Krell, A.; Ummenhofer, C.; Kattner, G.; Naumov, A.; Evans, D.; Dieckmann, G.S.; Thomas, D.N. The biology and chemistry of land fast ice in the White Sea, Russia – a comparison of winter and spring conditions. Polar Biol. 2003, 26, 707–719. [Google Scholar] [CrossRef] [Green Version]
- Ilyash, L.V.; Zhitina, L.S.; Kudryavtseva, V.A.; Mel’nikov, I.A. Seasonal dynamics of algae species composition and biomass in the coastal ice of Kandalaksha Bay, the White Sea. Biol. Bull. Rev. 2012, 73, 459–470. (In Russian) [Google Scholar]
- Majaneva, M.; Blomster, J.; Müller, S.; Autio, R.; Majaneva, S.; Hyytiäinen, K.; Nagai, S.; Rintala, J.-M. Sea-ice eukaryotes of the Gulf of Finland, Baltic Sea, and evidence for herbivory on weakly shade-adapted ice algae. Eur. J. Protistol. 2017, 57, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Belevich, T.A.; Ilyash, L.V. Picophytoplankton abundance in the Velikaya Salma strait, White Sea. Microbiology 2012, 81, 389–395. [Google Scholar] [CrossRef]
- Terrado, R.; Lovejoy, C.; Massana, R.; Vincent, W.F. Microbial food web responses to light and nutrients beneath the coastal Arctic Ocean sea ice during the winter–spring transition. J. Mar. Syst. 2008, 74, 964–977. [Google Scholar] [CrossRef] [Green Version]
- Epstein, S.; Lopez-Garcia, P. “Missing” protists: A molecular prospective. Biodivers. Conserv. 2008, 17, 261–276. [Google Scholar] [CrossRef]
- Massana, R.; Pedros-Alio, C. Unveiling new microbial eukaryotes in the surface ocean. Curr. Opin. Microbiol. 2008, 11, 213–218. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, N.; Daugbjerg, N.; Gabrielsen, T.M. Molecular diversity and temporal variation of picoeukaryotes in two Arctic fjords, Svalbard. Polar Biol. 2012, 35, 519–533. [Google Scholar] [CrossRef]
- Nielsen, K.M.; Johnsen, P.J.; Bensasson, D.; Daffonchio, D. Release and persistence of extracellular DNA in the environment. Environ. Biosaf. Res. 2007, 6, 37–53. [Google Scholar] [CrossRef] [Green Version]
- Charvet, S.; Vincent, W.F.; Comeau, A.; Lovejoy, C. Pyrosequencing analysis of the protist communities in a high Arctic meromictic lake: DNA preservation and change. Front. Microbiol. 2012, 3, 422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Not, F.; del Campo, J.; Balagué, V.; de Vargas, C.; Massana, R. New Insights into the Diversity of Marine Picoeukaryotes. PLoS ONE 2009, 4, e7143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.; Massana, R.; Not, F.; Marie, D.; Vaulot, D. Mapping of picoeukaryotes in marine ecosystems with quantitative PCR of the 18S rRNA gene. FEMS Microb. Ecol. 2005, 52, 79–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potvin, M.; Lovejoy, C. PCR-based diversity estimates of artificial and environmental 18S rRNA gene libraries. J. Eukaryot. Microbiol. 2009, 56, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Luddington, I.; Kaczmarska, I.; Lovejoy, C. Distance and character-based evaluation of the V4 region of the 18S rRNA gene for the identification of diatoms (Bacillariophyceae). PLoS ONE 2012, 7, e45664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balzano, S.; Percopo, I.; Siano, R.; Gourvil, P.; Chanoine, M.; Dominique, M.; Vaulot, D.; Sarno, D. Morphological and genetic diversity of Beaufort Sea diatoms with high contributions from the Chaetoceros neogracilis species complex. J. Phycol. 2017, 53, 161–187. [Google Scholar] [CrossRef] [Green Version]
- Marquardt, M.; Vader, A.; Stübner, E.I.; Reigstad, M.; Gabrielsen, T.M. Strong seasonality of marine microbial eukaryotes in a high-Arctic fjord (Isfjorden, in West Spitsbergen, Norway). Appl. Environ. Microbiol. 2016, 82, 1868–1880. [Google Scholar] [CrossRef] [Green Version]
- Kilias, E.S.; Nöthig, E.M.; Wolf, C.; Metfies, K. Picoeukaryote plankton composition off West Spitsbergen at the Entrance to the Arctic Ocean. J. Eukaryot. Microbiol. 2014, 61, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Monier, A.; Comte, J.; Babin, M.; Forest, A.; Matsuoka, A.; Lovejoy, C. Oceanographic structure drives the assembly processes of microbial eukaryotic communities. ISME J. 2015, 9, 990–1002. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; He, J.; Lin, L.; Jin, H. Dominance of picophytoplankton in the newly open surface water of the central Arctic Ocean. Polar Biol. 2015, 38, 1081–1089. [Google Scholar] [CrossRef]
- Joli, N.; Monier, A.; Logares, R.; Lovejoy, C. Seasonal patterns in Arctic prasinophytes and inferred ecology of Bathycoccus unveiled in an Arctic winter metagenome. ISME J. 2017, 11, 1372–1385. [Google Scholar] [CrossRef] [Green Version]
- Kilias, E.; Wolf, C.; Nöthig, E.M.; Peeken, I.; Metfies, K. Protist distribution in the western Fram Strait in summer 2010 based on 454-pyroseqeuncing of 18S rDNA. J. Phycol. 2013, 49, 996–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balzano, S.; Gourvil, P.; Siano, R.; Chanoine, M.; Marie, D.; Lessard, S.; Sarno, D.; Vaulot, D. Diversity of cultured photosynthetic flagellates in the northeast Pacific and Arctic Oceans in summer. Biogeosciences 2012, 9, 4553–4571. [Google Scholar] [CrossRef] [Green Version]
- Foulon, E.; Not, F.; Jalabert, F.; Cariou, T.; Massana, R.; Simon, N. Ecological niche partitioning in the picoplanktonic green alga Micromonas pusilla: Evidence from environmental surveys using phylogenetic probes. Environ. Microbiol. 2008, 10, 2433–2443. [Google Scholar] [CrossRef] [Green Version]
- Simon, N.; Foulon, E.; Grulois, D.; Six, C.; Desdevises, Y.; Latimier, M.; Le Gall, F.; Tragin, M.; Houdan, A.; Derelle, E.; et al. Revision of the genus Micromonas Manton et Parke (Chlorophyta, Mamiellophyceae), of the type species M. pusilla (Butcher) Manton & Parke and of the species M. commoda van Baren, Bachy and Worden and description of two new species based on the genetic and phenotypic characterization of cultured isolates. Protist 2017, 168, 612–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slapeta, J.; López-García, P.; Moreira, D. Global dispersal and ancient cryptic species in the smallest marine eukaryotes. Mol. Biol. Evol. 2006, 23, 23–29. [Google Scholar] [CrossRef]
- Worden, A.Z.; Lee, J.-H.; Mock, T.; Rouzé, P.; Simmons, M.P.; Aerts, A.L.; Allen, A.E.; Cuvelier, M.L.; Derelle, E.; Everett, M.V.; et al. Green evolution and dynamic adaptations revealed by genomes of the marine picoeukaryotes Micromonas. Science 2009, 324, 268–272. [Google Scholar] [CrossRef] [Green Version]
- Courties, C.; Perasso, R.; Chrétiennot-Dinet, M.-J.; Gouy, M.; Guillou, L.; Troussellier, M. Phylogenetic analysis and genome size of Ostreococcus tauri (Chlorophyta, Prasinophyceae). J. Phycol. 1998, 34, 844–849. [Google Scholar] [CrossRef]
- Vannier, T.; Leconte, J.; Seeleuthner, Y.; Mondy, S.; Pelletier, E.; Aury, J.-M.; de Vargas, C.; Sieracki, M.; Iudicone, D.; Vaulot, D.; et al. Survey of the green picoalga Bathycoccus genomes in the global ocean. Sci. Rep. 2016, 6, 37900. [Google Scholar] [CrossRef] [Green Version]
- Vaulot, D.; Lepére, C.; Toulza, E.; De la Iglesia, R.; Poulain, J.; Gaboyer, F.; Moreau, H.; Vandepoele, K.; Ulloa, O.; Gavory, F.; et al. Metagenomes of the picoalga Bathycoccus from the Chile Coastal Upwelling. PLoS ONE 2012, 7, e39648. [Google Scholar] [CrossRef]
- Limardo, A.J.; Sudek, S.; Choi, C.J.; Poirier, C.; Rii, Y.M.; Blum, M.; Roth, R.; Goodenough, U.; Church, M.J.; Worden, A.Z. Quantitative biogeography of picoprasinophytes establishes ecotype distributions and significant contributions to marine phytoplankton. Environ. Microbiol. 2017, 19, 3219–3234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majaneva, M.; Rintala, J.M.; Piisila, M.; Fewer, D.P.; Blomster, J. Comparison of wintertime eukaryotic community from sea ice and open water in the Baltic Sea, based on sequencing of the 18S rRNA gene. Polar Biol. 2012, 35, 875–889. [Google Scholar] [CrossRef]
- Booth, B.C.; Larouche, P.; Belanger, S.; Klein, B.; Amiel, D.; Mei, Z.P. Dynamics of Chaetoceros socialis blooms in the North Water. Deep Sea Res. II Top. Stud. Oceanogr. 2002, 49, 5003–5025. [Google Scholar] [CrossRef]
- Degerlund, M.; Eilertsen, H.C. Main species characteristics of phytoplankton spring blooms in NE Atlantic and Arctic waters (68–80° N). Estuar. Coast. Shelf Sci. 2010, 33, 242–269. [Google Scholar] [CrossRef]
- Dasilva, C.R.; Li, W.K.W.; Lovejoy, C. Phylogenetic diversity of eukaryotic marine microbial plankton on the Scotian Shelf, Northwestern Atlantic Ocean. J. Plankton Res. 2013, 36, 344–363. [Google Scholar] [CrossRef]
- Egge, E.S.; Johannessen, T.V.; Andersen, T.; Eikrem, W.; Bittner, L.; Larsen, A.; Sandaa, R.A.; Edvardsen, B. Seasonal diversity and dynamics of haptophytes in the Skagerrak, Norway, explored by high-throughput sequencing. Mol. Ecol. 2015, 24, 3026–3042. [Google Scholar] [CrossRef]
- Liu, H.; Probert, I.; Uitz, J.; Claustre, H.; Aris-Brosou, S.; Frada, M.; Not, F.; Vargas, C. Extreme diversity in noncalcifying haptophytes explains a major pigment paradox in open oceans. Proc. Natl. Acad. Sci. USA 2009, 106, 12803–12808. [Google Scholar] [CrossRef] [Green Version]
- Orsi, W.; Song, Y.C.; Hallam, S.; Edgcomb, V. Effect of oxygen minimum zone formation on communities of marine protist. ISME J. 2012, 6, 1586–1601. [Google Scholar] [CrossRef]
- Ichinomiya, M.; Yoshikawa, S.; Kamiya, M.; Ohki, K.; Takaichi, S.; Kuwata, A. Isolation and characterization of Parmales (Heterokonta/Heterokontophyta/Stramenopiles) from the Oyashio region, Western North Pacific. J. Phycol. 2011, 47, 144–151. [Google Scholar] [CrossRef]
- Ichinomiya, M.; dos Santos, A.; Gourvil, P.; Yoshikawa, S.; Kamiya, M.; Ohki, K.; Audic, S.; de Vargas, C.; Noël, M.-H.; Vaulot, D.; et al. Diversity and oceanic distribution of the Parmales (Bolidophyceae), a picoplanktonic group closely related to diatoms. ISME J. 2016, 10, 2419–2434. [Google Scholar] [CrossRef]
- Kuwata, A.; Yamada, K.; Ichinomiya, M.; Yoshikawa, S.; Tragin, M.; Vaulot, D.; dos Santos, A. Bolidophyceae, a Sister Picoplanktonic Group of Diatoms—A Review. Front. Mar. Sci. 2018, 5, 370. [Google Scholar] [CrossRef]
- Komuro, C.; Narita, H.; Imai, K.; Nojiri, Y.; Jordan, R.W. Microplankton assemblages at Station KNOT in the subarctic western Pacific, 1999–2000. Deep Sea Res. II Top. Stud. Oceanogr. 2005, 52, 2206–2217. [Google Scholar] [CrossRef]
- Ichinomiya, M.; Nakamachi, M.; Shimizu, Y.; Kuwata, A. Growth characteristics and vertical distribution of Triparma laevis (Parmales) during summer in the Oyashio region, western North Pacific. Aquat. Microb. Ecol. 2013, 68, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Piwosz, K.; Wiktor, J.M.; Niemi, A.; Tatarek, A.; Michel, C. Mesoscale distribution and functional diversity of picoeukaryotes in the first-year sea ice of the Canadian Arctic. ISME J. 2013, 7, 1461–1471. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Latitude (N) Longitude (E) | Date | Ice Thickness (cm) | Under Ice Water | Chl-a (μg/L) | N (cells/mL) | B (µg C/L) | N (%) | B (%) | |
|---|---|---|---|---|---|---|---|---|---|---|
| Salinity (psu) | Temperature (°C) | |||||||||
| 1/13w | 66°32.01’ | 19 Mar 2013 | 71 | 14.9 | –1.0 | 0.16 | 140 | 0.17 | 11 | 25 |
| 1/14w | 33°6.54’ | 16 Mar 2014 | 58 | 15.6 | –1.1 | 0.22 | 50 | 0.07 | 15 | 35 |
| 2/13w | 66°33.20’ | 23 Mar 2013 | 22 | 24.5 | –0.7 | 0.05 | 10 | 0.06 | 5 | 43 |
| 2/14w | 33°6.28’ | 17 Mar 2014 | 49 | 24.5 | –1.2 | 0.73 | 20 | 0.07 | 33 | 78 |
| 3/14w | 66°33.12’ 33°2.11’ | 17 Mar 2014 | 52 | 21.9 | –0.9 | 0.32 | 20 | 0.03 | 100 | 100 |
| 4/14w | 66°34.87’ 32°58.89’ | 19 Mar 2014 | 45 | 26.7 | –1.1 | 0.10 | 80 | 0.19 | 53 | 86 |
| 5/14w | 66°32.14’ 33°13.17’ | 15 Mar 2014 | 26 | 25.5 | –1.2 | 0.31 | 10 | 0.01 | 5 | 11 |
| Sample | Total Number of V4 Tag Sequences | Number of V4 Sequences of Eukaryotes Groups after Quality Filtering | Number of Eukaryotes OTUs (97% Similarity) | Number of PPE Reads | Number of PPE OTUs |
|---|---|---|---|---|---|
| 1/13w | 26,493 | 7398 | 609 | 384 | 34 |
| 2/13w | 24,600 | 19,080 | 1227 | 1542 | 98 |
| 1/14w | 27,019 | 12,334 | 822 | 1347 | 72 |
| 2/14w | 28,525 | 7645 | 618 | 1216 | 141 |
| 3/14w | 39,578 | 17,845 | 1393 | 1620 | 83 |
| 4/14w | 27,227 | 7974 | 738 | 1101 | 140 |
| 5/14w | 94,682 | 50,227 | 3856 | 5947 | 186 |
| Total | 268,124 | 122,503 | 9263 | 13,157 | 754 |
| Taxonomic Group | Reads (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| 1/13w | 2/13w | 1/14w | 2/14w | 3/14w | 4/14w | 5/14w | ||
| Chloroplastida | Mamiellophyceae | 81.2 (59) | 27.3 (23) | 71.3 (50) | 91.8 (89) | 82.4 (67) | 60.8 (52) | 70.2 (56) |
| Pyramimonadophyceae | 3.9 (6) | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Palmophyllophyceae | 0.0 | 3.6 (3) | 0.0 | 0.0 | 0.0 | 1.8 (2) | 0.1 (1) | |
| Trebouxiophyceae | 5.0 (6) | 1.4 (4) | 6.8 (8) | 0.2 (1) | 4.5 (7) | 3.0 (6) | 0.2 (1) | |
| Stramenopile | Bolidophyceae | 6.8 (21) | 58.1 (67) | 21.9 (42) | 8.1 (12) | 13.1 (24) | 29.4 (36) | 19.6 (39) |
| Mediophyceae | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.2 (2) | |
| Haptophyta | Coccolithophyceae | 3.1 (9) | 9.5 (3) | 0.0 | 0.0 | 0.0 | 5.0 (4) | 9.7 (2) |
| Expected OTUs richness of PPEs | 36 | 71 | 57 | 105 | 63 | 110 | 103 | |
| Taxonomic Group | Reads (%) | ||||||
|---|---|---|---|---|---|---|---|
| 1/13w | 2/13w | 1/14w | 2/14w | 3/14w | 4/14w | 5/14w | |
| Micromonas polaris | 1.6 | 17.1 | 1.7 | 54.9 | 5.4 | 66.7 | 53.1 |
| Micromonas commoda A2 | 0.0 | 1.2 | 0.0 | 0.0 | 0.0 | 0.6 | 0.5 |
| Micromonas clade F (B3) | 0.0 | 0.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.4 |
| total Micromonas | 1.6 | 19.0 | 1.7 | 54.9 | 5.4 | 67.3 | 54.1 |
| Mantoniella squamata | 85.2 | 36.4 | 11.2 | 7.9 | 1.4 | 4.9 | 14.6 |
| Mantoniella clade 1 | 7.7 | 4.2 | 42.4 | 25.8 | 5.4 | 8.7 | 7.1 |
| Mantoniella clade 2 | 1.3 | 0.0 | 13.0 | 0.6 | 11.7 | 0.0 | 0.0 |
| Mantoniella clade 3 | 3.5 | 24.6 | 30.8 | 1.4 | 75.9 | 8.7 | 4.5 |
| Mantoniella clade 4 | 0.0 | 1.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Total Mantoniella | 97.7 | 66.6 | 97.4 | 35.8 | 94.4 | 22.3 | 26.2 |
| Bathycoccus prasinos | 0.0 | 4.7 | 0.0 | 0.3 | 0.0 | 3.3 | 2.1 |
| Ostreococcus tauri | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.1 |
| uncultured eukaryotic clone DSGM-81 | 0.6 | 9.2 | 0.9 | 8.1 | 0.2 | 6.7 | 14.0 |
| Crustomastix sp. MBIC10709 | 0.0 | 0.5 | 0.0 | 1.0 | 0.0 | 0.4 | 3.6 |
| OTU | Closest Match | Identity | Origin |
|---|---|---|---|
| Mamiellophyceae | |||
| 2/13w18-73 MK571487 | Micromonas pusilla CCMP2099 (clade Ea) AY955000 (Micromonas polaris) | 100% | Baffin Bay, Canada |
| 5/14w56-22 MK571488 | Micromonas pusilla strain CS222 (clade C) AY955010 (Micromonas commoda A2) | 100% | South Pacific Ocean |
| 2/13w19-2 MK571489 | Uncultured Micromonas clade F MF589926 | 100% | the White Sea ice |
| Uncultured Prasinophyceae clone DH114_3A06 FJ032694 (Micromonas clade B3) | 100% | South Atlantic Ocean | |
| 1/13w8–15 MK571490 | Mantoniella squamata X73999 | 100% | - |
| Uncultured Prasinophyceae clone CFL146DB03 HM561186 | 100% | the Beaufort Sea | |
| 1/13w3–19 MK571491 | Mantoniella beaufortii KT860921 | 100% | the Beaufort Sea |
| Uncultured Mantoniella clade 1 MF589928 | 100% | the White Sea ice | |
| 1/14w17-29 MK571492 | Uncultured Mantoniella clade 2 MF589929 | 100% | the White Sea ice |
| Uncultured Chlorophyta clone 5-D5 FN690723 | 100% | the Baltic Sea | |
| 5/14w67-53 MK571493 | Uncultured Mantoniella clade 3 MF589930 | 100% | the White Sea ice |
| Uncultured Chlorophyta clone 5-D5 FN690723 | 98.4% | the Baltic Sea | |
| 2/13w14-6 MK571498 | Uncultured Prasinophyceae clone North_Pole_SI120_29 HQ438123 (uncultured Mantoniella clade 4) | 100% | North Pole sea ice |
| 4/14w54–15 MK571494 | Bathycoccus prasinos strain RCC801 KT860937 | 100% | English Channel, Atlantic Ocean |
| 5/14w57-4 MK571495 | Ostreococcus tauri Y15814 | 100% | the Mediterranean Sea |
| 2/14w103-9 MK571496 | Uncultured Crustomastix MF589934 | 100% | the White Sea ice |
| Uncultured eukaryote clone SHAX746 HQ868519 | 99.4% | Pacific Ocean, Canada | |
| 5/14w58-225 MK571497 | Uncultured eukaryotic clone DSGM81 AB275081 | 99.2% | methane cold seep sediment (Japan) |
| Uncultured Chlorophyta FN690728 | 100% | the Baltic Sea | |
| Pyramimonadophyceae | |||
| 1/13w72–12 MK571500 | Pyramimonas sp. RCC2009 JF794047 | 98.1% | the Beaufort Sea |
| Palmophyllophyceae | |||
| 4/14w79–16 MK571499 | Prasinoderma coloniale strain RCC854 KT860905 | 97.0% | Pacific Ocean |
| Uncultured eukaryote clone SHAX501 HQ868998 | 98.9% | Pacific Ocean, Canada | |
| Trebouxiophyceae | |||
| 1/14w39-6 MK571501 | Picochlorum sp. RCC748 KT860896 | 100% | Atlantic Ocean |
| 1/14w40-46 MK571502 | Choricystis minor X89012 | 100% | lake in Germany |
| Bolidophyceae | |||
| 2/13w b284-3 MK571511 | Triparma strigata KR998402 | 100% | Pacific Ocean, Japan |
| 2/13w b270-21 MK571512 | Uncultured bolidophyte LC190998 | 99.0% | Pacific Ocean, Japan |
| 2/13w b264-63 MK571513 | Uncultured bolidophyte LC191051 | 99.0% | Pacific Ocean, Japan |
| 3/14w b337-71 MK571514 | Uncultured stramenopile FN690655 | 99.0% | the Baltic Sea ice |
| 1/14w b286-6 MN684208 | Uncultured stramenopile FN690656 | 100% | the Baltic Sea ice |
| 5/14w b537-30 MK571516 | Uncultured eukaryote KT818381 | 97.8% | the Greenland Sea |
| 4/14w b371-20 MK571518 | Uncultured eukaryote KT811782 | 99.3% | the Greenland Sea |
| 3/14w b346-24 MK571515 | Uncultured eukaryote KT814386 | 98.8% | the Greenland Sea |
| 5/14w b565-83 MK571517 | Uncultured eukaryote KT815972 | 97.8% | the Greenland Sea |
| 2/13w b265-11 MK571519 | Uncultured eukaryote KT813573 | 99.5% | the Greenland Sea |
| 4/14w b360-4 MK571520 | Uncultured bolidophyte OTU:b474 MF589906 | 100% | The White Sea summer water |
| Uncultured bolidophyte LC191049 | 100% | Pacific Ocean | |
| 2/13w b282-3 MK571522 | Uncultured bolidophyte isolate OTU:b55 MF407369 | 100% | The White Sea ice |
| Uncultured eukaryote clone 52c_105508 KT814907 | 99.5% | the Greenland Sea | |
| 4/14w b397-2 MK571523 | Uncultured bolidophyte isolate OTU:b407 MF407373 | 100% | The White Sea summer water |
| Mediophyceae | |||
| 5/14w120-8 MN541095 | Chaetoceros cf. neogracilis strain RCC2318 JN934684 | 100% | the Beaufort Sea |
| 5/14w103-6 MK571504 | Skeletonema marinoi isolate 17 KR091067 | 100% | Atlantic Ocean |
| 5/14w121-3 MK571505 | Minutocellus polymorphus NIES-3970 LC189088 | 100% | - |
| Arcocellulus cornucervis strain RCC2270 JN934677 | 100% | the Beaufort Sea | |
| Haptophyta | |||
| 5/14w130-550 MK571506 | Phaeocystis pouchetii isolate AJ01 KR091066 | 100% | the North Sea |
| 5/14w125-4 MK571507 | Chrysochromulina clone MALINA JF698782 | 98.4% | the Beaufort Sea |
| Uncultured eukaryote KP405041 | 99.2% | the South China Sea | |
| 2/13w29-6 MK571508 | Chrysochromulina simplex AM491021 | 99.4% | - |
| Uncultured haptophyte Ma135-Pry1-C55 JX680441 | 100% | the Marmara Sea | |
| 4/14w91-6 MK571509 | Uncultured haptophyte FN690514 | 98.7% | the Baltic Sea |
| 5/14w134–14 MK571510 | Uncultured haptophyte KC488456 | 99.2% | the North Atlantic Ocean |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belevich, T.A.; Ilyash, L.V.; Milyutina, I.A.; Logacheva, M.D.; Troitsky, A.V. Photosynthetic Picoeukaryotes Diversity in the Underlying Ice Waters of the White Sea, Russia. Diversity 2020, 12, 93. https://doi.org/10.3390/d12030093
Belevich TA, Ilyash LV, Milyutina IA, Logacheva MD, Troitsky AV. Photosynthetic Picoeukaryotes Diversity in the Underlying Ice Waters of the White Sea, Russia. Diversity. 2020; 12(3):93. https://doi.org/10.3390/d12030093
Chicago/Turabian StyleBelevich, Tatiana A., Ludmila V. Ilyash, Irina A. Milyutina, Maria D. Logacheva, and Aleksey V. Troitsky. 2020. "Photosynthetic Picoeukaryotes Diversity in the Underlying Ice Waters of the White Sea, Russia" Diversity 12, no. 3: 93. https://doi.org/10.3390/d12030093
APA StyleBelevich, T. A., Ilyash, L. V., Milyutina, I. A., Logacheva, M. D., & Troitsky, A. V. (2020). Photosynthetic Picoeukaryotes Diversity in the Underlying Ice Waters of the White Sea, Russia. Diversity, 12(3), 93. https://doi.org/10.3390/d12030093