Settlement and Spreading of the Introduced Seaweed Caulacanthus okamurae (Rhodophyta) in the Mediterranean Sea




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
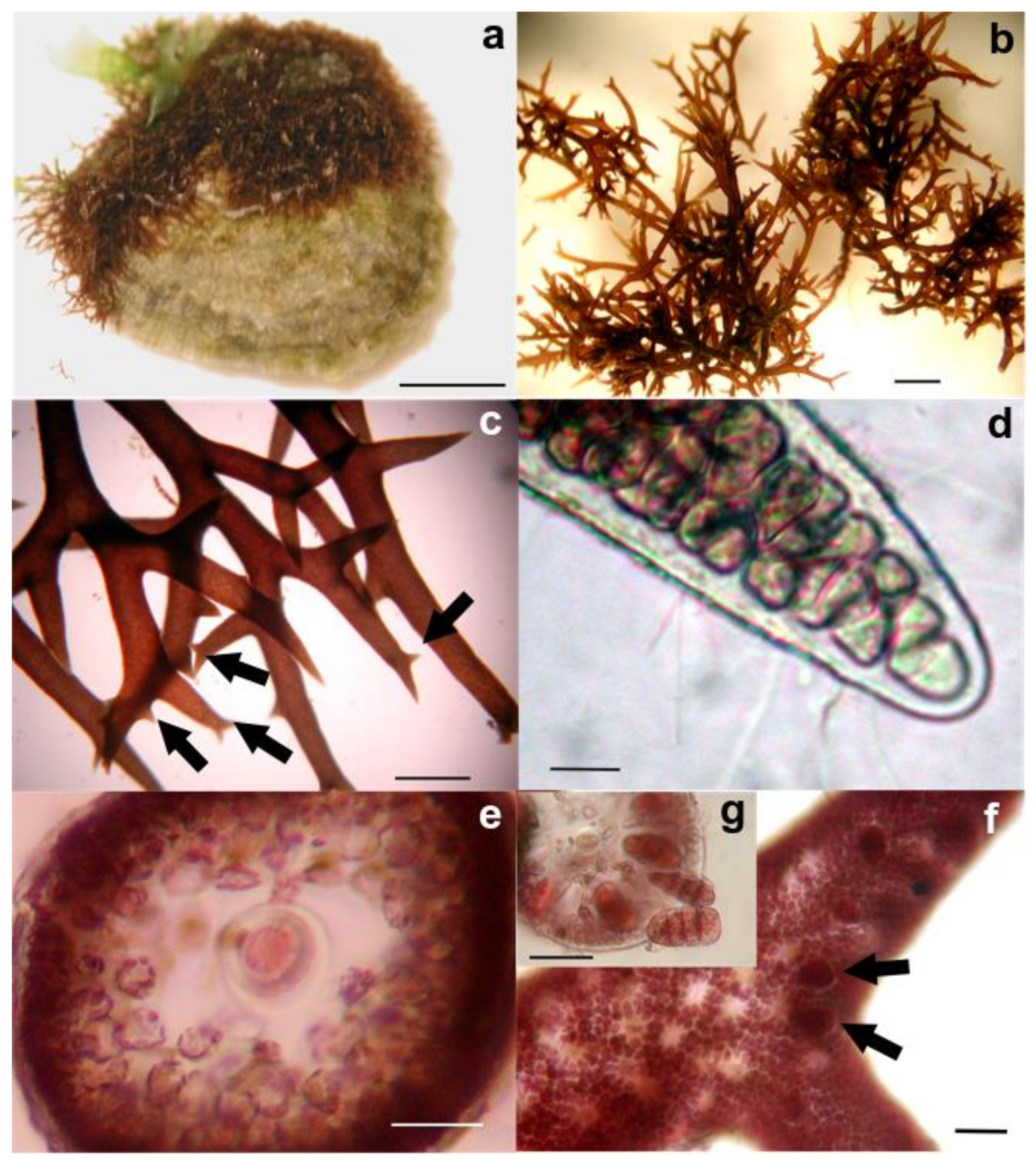
2. Materials and Methods
3. Results
3.1. Molecular Analyses
3.2. Morphological Observations
3.3. Distribution and Standing Crop Estimation
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guiry, M.D.; Guiry, G.M. AlgaeBase, National University of Ireland Galway. 2020. Available online: http://algaebase.org (accessed on 20 February 2020).
- Zuccarello, G.; West, J.; Rueness, J. Phylogeography of the cosmopolitan red alga Caulacanthus ustulatus (Caulacanthaceae, Gigartinales). Phycol. Res. 2002, 50, 163–172. [Google Scholar] [CrossRef]
- Yoshida, T. Marine Algae of Japan; Uchida Rokakuho Publishing Co.: Tokyo, Japan, 1998; pp. 1–1222. [Google Scholar]
- Choi, H.G.; Nam, K.W. Growth, tetrasporogenesis, and life history in culture of Caulacanthus okamurae (Gigartinales, Rhodophyta) from Korea. Bot. Mar. 2001, 44, 315–320. [Google Scholar] [CrossRef]
- Choi, H.G.; Nam, K.W.; Norton, T.A. No whirlwind romance: Typhoons, temperature and the failure of reproduction in Caulacanthus okamurae (Gigartinales, Rhodophyta). Eur. J. Phycol. 2001, 36, 353–358. [Google Scholar] [CrossRef]
- Yamada, Y. Notes on some Japanese algae V. J. Fac. Sci. Hokkaido Univ. 1933, 2, 277–285. [Google Scholar]
- Gabrielson, P.W.; Scagel, R.F. The marine algae of British Columbia, northern Washington, and southeast Alaska: Division Rhdophyta (red algae), class Rhodophyceae, order Gigartinales, family Caulacanthaceae and Plocamiaceae. Can. J. Bot. 1989, 67, 1221–1234. [Google Scholar] [CrossRef]
- Lindstrom, S.C.; Houghton, J.P.; Lees, D.C. Intertidal macroalgal community structure in southwestern Prince William Sound, Alaska. Bot. Mar. 1999, 42, 265–280. [Google Scholar] [CrossRef]
- Whiteside, K.E.; Murray, S.N. Spatial and temporal patterns of abundance in southern California populations of Caulacanthus ustulatus (Rhodophyta). J. Phycol. 2004, 40, 14. [Google Scholar]
- Maloney, E.; Fairey, R.; Lyman, A.; Reynolds, K.; Sigala, M. Introduced Aquatic Species in California Open Coastal Waters. Final Report; California Department of Fish and Game, Office of Spill Prevention and Response: Sacramento, CA, USA, 2006; p. 93. [Google Scholar]
- Miller, K.A.; Aguilar-Rosas, L.E.; Pedroche, F.F. A review of non-native seaweeds from California, USA and Baja California, Mexico. Hidrobiológica 2011, 21, 365–379. [Google Scholar]
- Fofonoff, P.W.; Ruiz, G.M.; Steves, B.; Simkanin, C.; Carlton, J.T. National Exotic Marine and Estuarine Species Information System, California. 2020. Available online: http://invasions.si.edu/nemesis/ (accessed on 20 February 2020).
- Smith, J.R.; Vogt, S.C.; Creedon, F.; Lucas, B.J.; Eernisse, D.J. The non-native turf-forming alga Caulacanthus ustulatus displaces space-occupants but increases diversity. Biol. Invasions 2014, 16, 2195–2208. [Google Scholar] [CrossRef]
- Rueness, J.; Rueness, E.K. Caulacanthus ustulatus (Gigartinales, Rhodophyta) from Brittany, France is an introduction from the Pacific Ocean. Cryptogamie Algol. 2000, 21, 355–363. [Google Scholar] [CrossRef]
- ICES Advisory Committee on the Marine Environment. Report of the Working Group on Introduction and Transfers of Marine Organisms (WGITMO); International Council for the Exploration of the Seas: Copenhagen, Denmark, 2011. [Google Scholar]
- Stegenga, H.; Karremans, M. Review of the red algal exotics in the marine waters of the Southwest Netherlands. Gorteria 2015, 27, 141–157. [Google Scholar]
- Day, F. Monitoring the marine invasive/non-native species of jetty beach, FSC Dale Fort, Pembrokeshire, Wales. Field Stud. 2018, 14, 1–7. [Google Scholar]
- Vollette, J.; Thirion, J.M.; Lahondère, C. Inventaire des macroalgues des estrans rocheux de l’estuaire de la Gironde. Bull. Soc. Bot. Centre-Ouest 2016, 46, 25–33. [Google Scholar]
- Verlaque, M.; Breton, G. Biological invasion: Long term monitoring of the macroalgal flora of a major European harbor complex. Mar. Pollut. Bull. 2019, 143, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Bárbara, I.; García-Redondo, V.; Díaz Tapia, P.; García-Fernández, A.; Piñeiro-Corbeira, C.; Peña, V.; Lugilde, J.; Cremades, J. Adiciones y correcciones a la flora bentónica marina del Atlántico ibérico norte. Acta Bot. Malac. 2019, 44, 51–60. [Google Scholar] [CrossRef]
- Verlaque, M.; Ruitton, S.; Mineur, F.; Boudouresque, C.-F. CIESM Atlas of Exotic Species in the Mediterranean 4: Macrophytes; CIESM Publishers: Monaco, 2015; p. 362. [Google Scholar]
- Cecere, E.; Petrocelli, A. The Mar Piccolo of Taranto. In Flora and Vegetation of the Italian Transitional Water Systems; Cecere, E., Petrocelli, A., Izzo, G., Sfriso, A., Eds.; CoRiLa, Stampa Multigraf Spinea: Venice, Italy, 2009; pp. 195–227. [Google Scholar]
- Cecere, E.; Alabiso, G.; Carlucci, R.; Petrocelli, A.; Verlaque, M. Fate of two invasive or potentially invasive alien seaweeds in a central Mediterranean transitional water system: Failure and success. Bot. Mar. 2016, 59, 451–462. [Google Scholar] [CrossRef]
- Sfriso, A.; Facca, C. Distribution and production of macrophytes in the lagoon of Venice. Comparison of actual and past abundance. Hydrobiologia 2007, 577, 71–85. [Google Scholar] [CrossRef]
- Sfriso, A.; Buosi, A.; Mistri, M.; Munari, C.; Franzoi, P.; Sfriso, A.A. Long-term changes of the trophic status in transitional ecosystems of the northern Adriatic Sea, key parameters and future expectations: The lagoon of Venice as a study case. Nat. Conserv. 2019, 34, 193–215. [Google Scholar] [CrossRef]
- Freshwater, D.W.; Rueness, J. Phylogenetic relationships of some European Gelidium (Gelidiales, Rhodophyta) species, based on rbcL nucleotide sequence analysis. Phycologia 1994, 33, 187–194. [Google Scholar] [CrossRef]
- Wolf, M.A.; Sciuto, K.; Maggs, C.A.; Barros-Barreto, M.B.; Andreoli, C.; Moro, I. Ceramium Roth (Ceramiales, Rhodophyta) from Venice lagoon (Adriatic Sea, Italy): Comparative studies of Mediterranean and Atlantic taxa. Taxon 2011, 60, 1584–1595. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, G. Estimating the dimension of a model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Sfriso, A. Flora and vertical distribution of macroalgae in the lagoon of Venice: A comparison with previous studies. Plant Biosyst. 1987, 121, 69–85. [Google Scholar] [CrossRef]
- Petrocelli, A.; Cecere, E.; Rubino, F. Successions of phytobenthos species in a Mediterranean transitional water system: The importance of long term observations. Nat. Conserv. 2019, 34, 217–246. [Google Scholar] [CrossRef] [Green Version]
- Petrocelli, A.; Alabiso, G.; Cecere, E.; Ricci, P.; Carlucci, R. Invasive or not? The case of Grateloupia turuturu (Rhodophyta, Halymeniales) in the Northern Ionian Sea (Mediterranean Sea). Estuar. Coast. Shelf Sci. Under review.
- Rueness, J. A culture study of Caulacanthus ustulatus (Caulacanthaceae, Gigartinales, Rhodophyta) from Europe and Asia. Cryptogamie Algol. 1997, 2, 175–185. [Google Scholar]
- Marchini, A.; Ferrario, J.; Sfriso, A.; Occhipinti-Ambrogi, A. Current status and trends of biological invasions in the Lagoon of Venice, a hotspot of marine NIS introductions in the Mediterranean Sea. Biol. Invasions 2015, 17, 2943–2962. [Google Scholar] [CrossRef] [Green Version]
- Sfriso, A.; Marchini, A. Updating of non-indigenous macroalgae in the Italian Coasts. New introductions and cryptic species. Biol. Mar. Mediterr. 2014, 21, 60–69. [Google Scholar]
- Sfriso, A.; Buosi, A.; Wolf, M.A.; Sfriso, A.A. Invasion of alien macroalgae in the Venice Lagoon, a pest or a resource? Aquat. Invasions 2020, 15. Available online: https://www.reabic.net/aquaticinvasions/2020/ACCEPTED/AI_2020_Sfriso_etal_correctedproof.pdf.
- Sfriso, A.; Curiel, D. Check-list of marine seaweeds recorded in the last 20 years in Venice lagoon and a comparison with the previous records. Bot. Mar. 2007, 50, 22–58. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrocelli, A.; Wolf, M.A.; Cecere, E.; Sciuto, K.; Sfriso, A. Settlement and Spreading of the Introduced Seaweed Caulacanthus okamurae (Rhodophyta) in the Mediterranean Sea. Diversity 2020, 12, 129. https://doi.org/10.3390/d12040129
Petrocelli A, Wolf MA, Cecere E, Sciuto K, Sfriso A. Settlement and Spreading of the Introduced Seaweed Caulacanthus okamurae (Rhodophyta) in the Mediterranean Sea. Diversity. 2020; 12(4):129. https://doi.org/10.3390/d12040129
Chicago/Turabian StylePetrocelli, Antonella, Marion A. Wolf, Ester Cecere, Katia Sciuto, and Adriano Sfriso. 2020. "Settlement and Spreading of the Introduced Seaweed Caulacanthus okamurae (Rhodophyta) in the Mediterranean Sea" Diversity 12, no. 4: 129. https://doi.org/10.3390/d12040129
APA StylePetrocelli, A., Wolf, M. A., Cecere, E., Sciuto, K., & Sfriso, A. (2020). Settlement and Spreading of the Introduced Seaweed Caulacanthus okamurae (Rhodophyta) in the Mediterranean Sea. Diversity, 12(4), 129. https://doi.org/10.3390/d12040129