Niche Models Differentiate Potential Impacts of Two Aquatic Invasive Plant Species on Native Macrophytes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
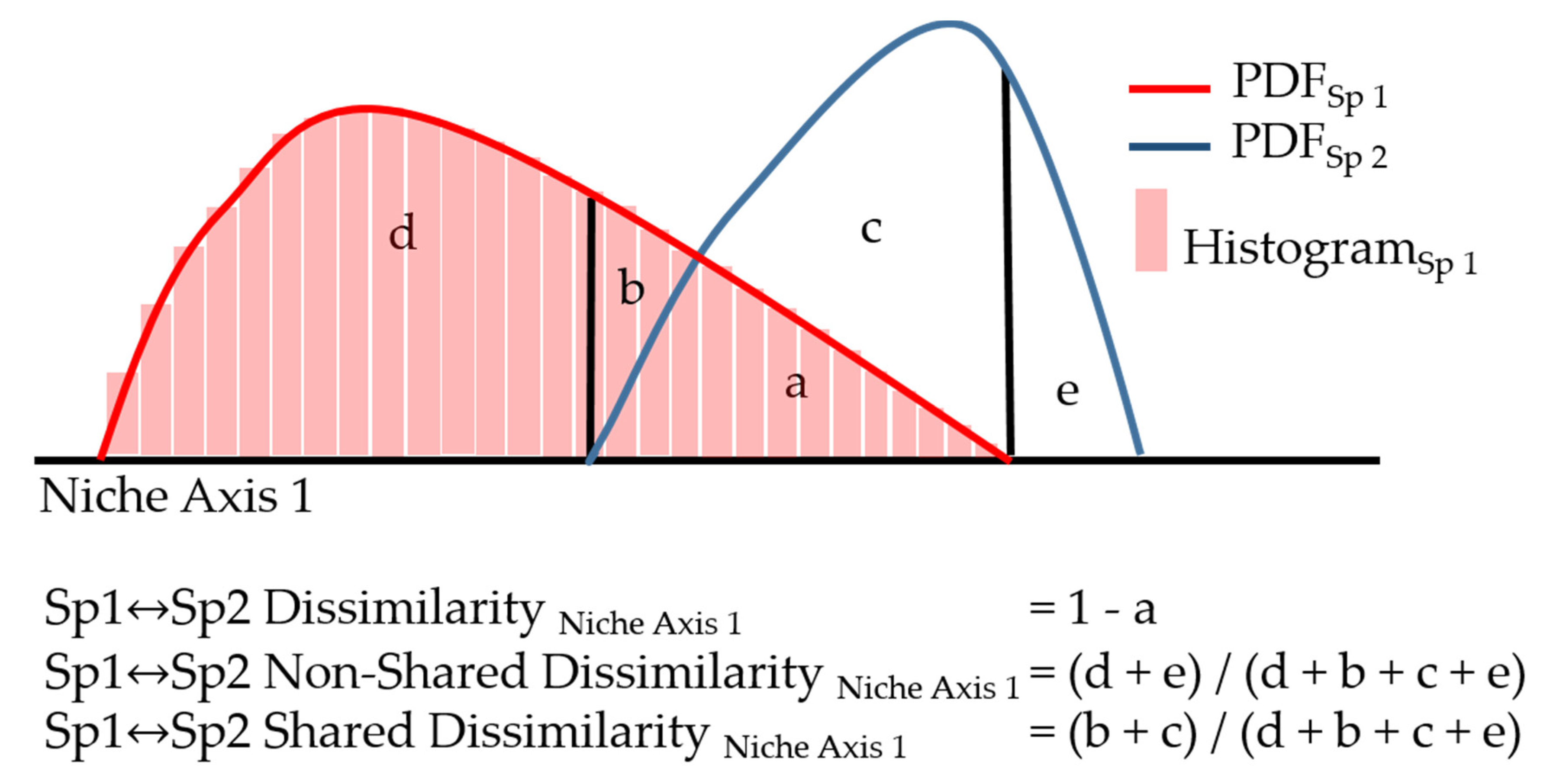
2. Materials and Methods
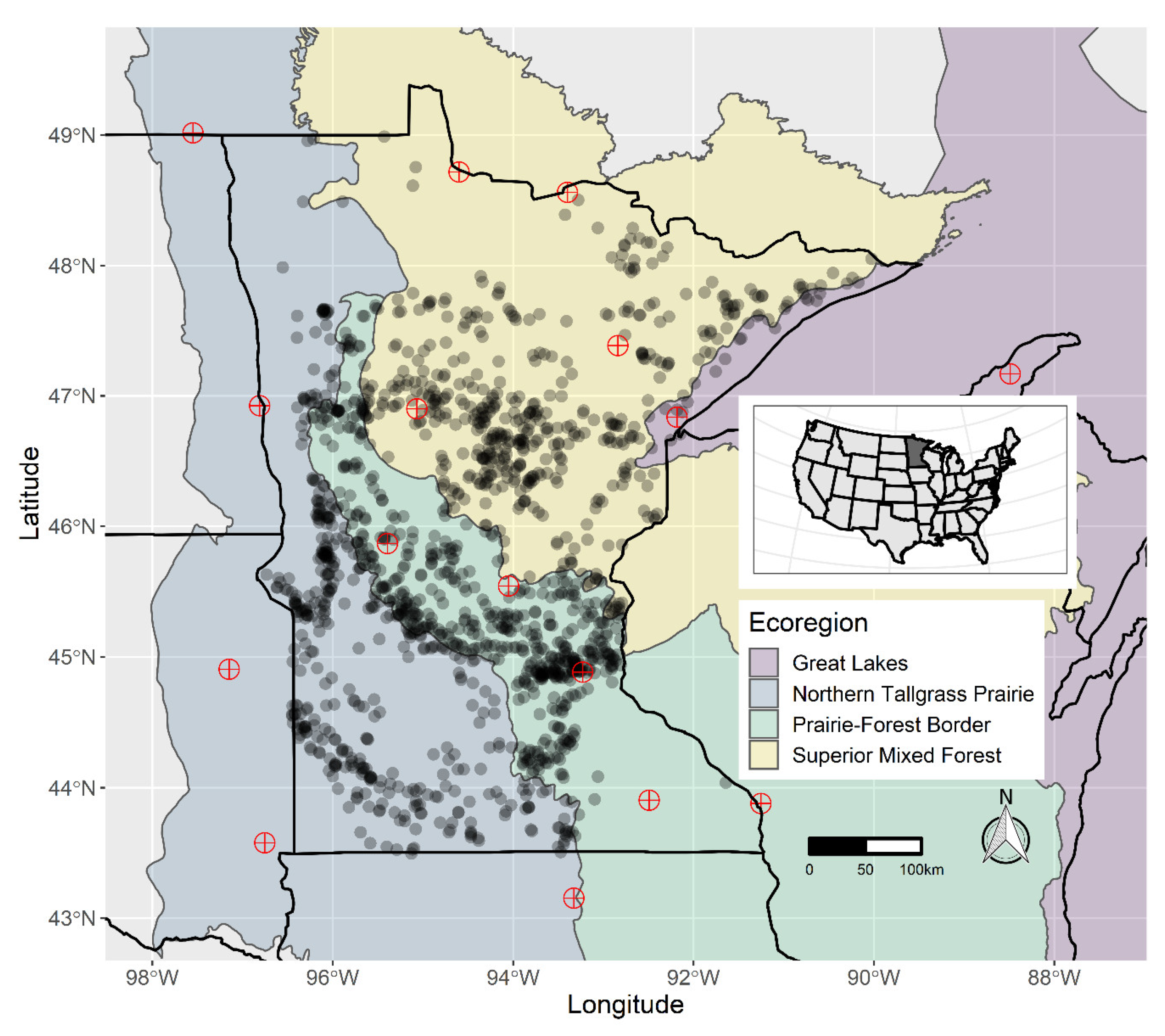
2.1. Macrophyte Occurrence Data
2.2. Environmental Data
2.3. Data Analysis
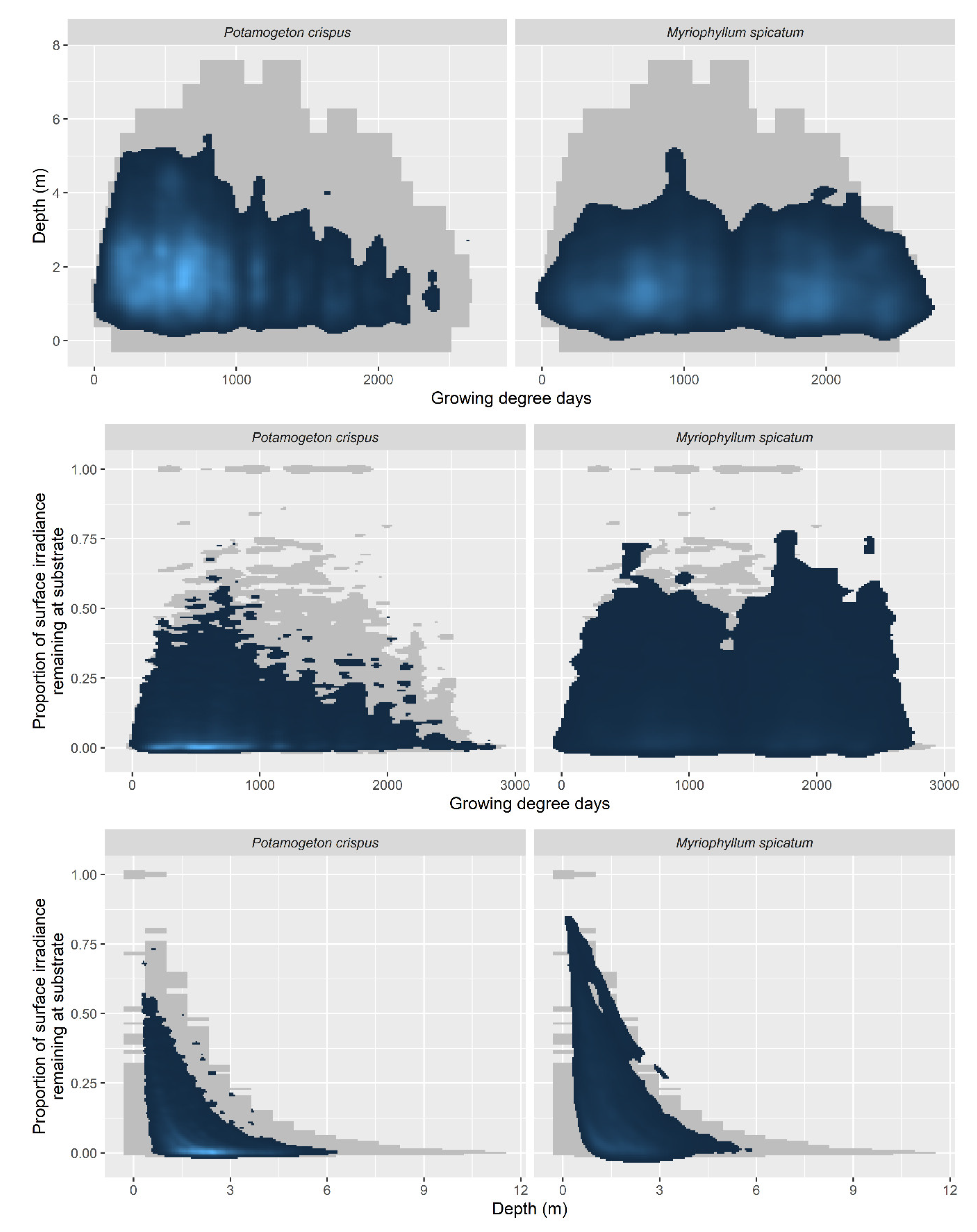
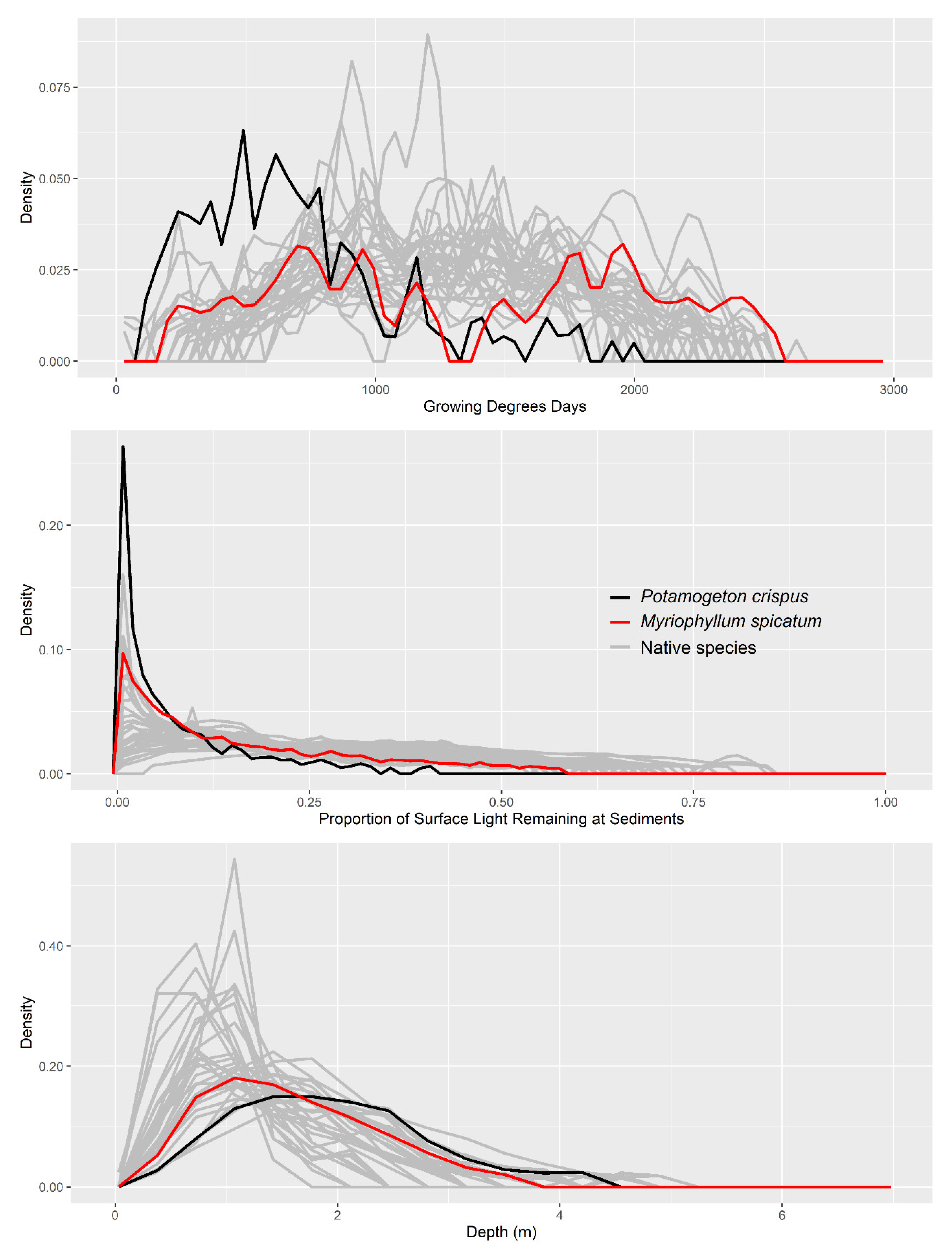
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Courchamp, F.; Fournier, A.; Bellard, C.; Bertelsmeier, C.; Bonnaud, E.; Jeschke, J.M.; Russell, J.C. Invasion biology: Specific problems and possible solutions. Trends Ecol. Evol. 2017, 32, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Bellard, C.; Cassey, P.; Blackburn, T.M. Alien species as a driver of recent extinctions. Biol. Lett. 2016, 12, 20150623. [Google Scholar] [CrossRef]
- Bradley, B.A.; Laginhas, B.B.; Whitlock, R.; Allen, J.M.; Bates, A.E.; Bernatchez, G.; Diez, J.M.; Early, R.; Lenoir, J.; Vilà, M.; et al. Disentangling the abundance-impact relationship for invasive species. Proc. Natl. Acad. Sci. USA 2019, 116, 9919–9924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moles, A.T.; Flores-Moreno, H.; Bonser, S.P.; Warton, D.I.; Helm, A.; Warman, L.; Eldridge, D.J.; Jurado, E.; Hemmings, F.A.; Reich, P.B.; et al. Invasions: The trail behind, the path ahead, and a test of a disturbing idea. J. Ecol. 2012, 100, 116–127. [Google Scholar] [CrossRef]
- Boltovskoy, D.; Sylvester, F.; Paolucci, E.M. Invasive species denialism: Sorting out facts, beliefs, and definitions. Ecol. Evol. 2018, 8, 11190–11198. [Google Scholar] [CrossRef] [Green Version]
- Schirmel, J.; Bundschuh, M.; Entling, M.H.; Kowarik, I.; Buchholz, S. Impacts of invasive plants on resident animals across ecosystems, taxa, and feeding types: A global assessment. Glob. Chang. Biol. 2016, 22, 594–603. [Google Scholar] [CrossRef]
- Seabloom, E.W.; Harpole, W.S.; Reichman, O.J.; Tilman, D. Invasion, competitive dominance, and resource use by exotic and native California grassland species. Proc. Natl. Acad. Sci. USA 2003, 100, 13384–13389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleming, J.P.; Dibble, E.D. Ecological mechanisms of invasion success in aquatic macrophytes. Hydrobiologia 2015, 746, 23–37. [Google Scholar] [CrossRef]
- Levine, J.M.; Vilà, M.; D’Antonio, C.M.; Dukes, J.S.; Grigulis, K.; Lavorel, S. Mechanisms underlying the impacts of exotic plant invasions. Proc. R. Soc. Lond. 2003, 270, 775–781. [Google Scholar] [CrossRef]
- Blumenthal, D.M.; Hufbauer, R.A. Increased plant size in exotic populations: A common-garden test with 14 invasive species. Ecology 2007, 88, 2758–2765. [Google Scholar] [CrossRef] [Green Version]
- MacDougall, A.S.; Turkington, R. Are invasive species the drivers or passengers of change in degraded ecosystems? Ecology 2005, 86, 42–55. [Google Scholar] [CrossRef] [Green Version]
- Hillerislambers, J.; Adler, P.B.; Harpole, W.S.; Levine, J.M.; Mayfield, M.M. Rethinking community assembly through the lens of coexistence theory. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 227–248. [Google Scholar] [CrossRef] [Green Version]
- Chesson, P. Mechanisms of maintenance of species diversity. Annu. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef] [Green Version]
- Shea, K.; Chesson, P. Community ecology theory as a framework for biological invasions. Trends Ecol. Evol. 2002, 17, 170–176. [Google Scholar] [CrossRef]
- Divíšek, J.; Chytrý, M.; Beckage, B.; Gotelli, N.J.; Lososová, Z.; Pyšek, P.; Richardson, D.M.; Molofsky, J. Similarity of introduced plant species to native ones facilitates naturalization, but differences enhance invasion success. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Zwerschke, N.; van Rein, H.; Harrod, C.; Reddin, C.; Emmerson, M.C.; Roberts, D.; O’Connor, N.E. Competition between co-occurring invasive and native consumers switches between habitats. Funct. Ecol. 2018, 32, 2717–2729. [Google Scholar] [CrossRef]
- Escoriza, D.; Ruhí, A. Functional distance to recipient communities may favour invasiveness: Insights from two invasive frogs. Divers. Distrib. 2016, 22, 519–533. [Google Scholar] [CrossRef] [Green Version]
- Fournier, A.; Penone, C.; Pennino, M.G.; Courchamp, F. Predicting future invaders and future invasions. Proc. Natl. Acad. Sci. USA 2019, 116, 7905–7910. [Google Scholar] [CrossRef] [Green Version]
- Mack, R.N. Predicting the identity and fate of plant invaders: Emergent and emerging approaches. Biol. Conserv. 1996, 78, 107–121. [Google Scholar] [CrossRef]
- Darwin, C.R. On the Origin of Species by Means of Natural Selection, or Preservation of Favoured Races in the Struggle for Life; John Murray: London, UK, 1859. [Google Scholar]
- Patten, B.C.; Auble, G.T. System theory of the ecological niche. Am. Nat. 1981, 117, 893–922. [Google Scholar] [CrossRef]
- Elton, C.S. The Ecology of Invasions by Animals and Plants; Springer: Boston, MA, USA, 1958. [Google Scholar]
- Carpenter, S.R.; Mooney, H.A.; Agard, J.; Capistrano, D.; DeFries, R.S.; Diaz, S.; Dietz, T.; Duraiappah, A.K.; Oteng-Yeboah, A.; Pereira, H.M.; et al. Science for managing ecosystem services: Beyond the Millennium Ecosystem Assessment. Proc. Natl. Acad. Sci. USA 2009, 106, 1305–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grinnell, J. The niche-relationships of the California Thrasher. Auk 1917, 34, 427–433. [Google Scholar] [CrossRef]
- Gallardo, B.; Clavero, M.; Sánchez, M.I.; Vilà, M. Global ecological impacts of invasive species in aquatic ecosystems. Glob. Chang. Biol. 2016, 22, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Hussner, A.; Stiers, I.; Verhofstad, M.J.J.M.; Bakker, E.S.; Grutters, B.M.C.; Haury, J.; van Valkenburg, J.L.C.H.; Brundu, G.; Newman, J.; Clayton, J.S.; et al. Management and control methods of invasive alien aquatic plants: A review. Aquat. Bot. 2017, 136, 112–137. [Google Scholar] [CrossRef]
- Hussner, A.; Van De Weyer, K.; Gross, E.M.; Hilt, S. Comments on increasing number and abundance of non-indigenous aquatic macrophyte species in Germany. Weed Res. 2010, 50, 519–526. [Google Scholar] [CrossRef]
- Capers, R.; Selsky, R.; Bugbee, G.; White, J. Aquatic plant community invasibility and scale-dependent patterns in native and invasive species richness. Ecology 2007, 88, 3135–3143. [Google Scholar] [CrossRef]
- Muthukrishnan, R.; Hansel-Welch, N.; Larkin, D.J. Environmental filtering and competitive exclusion drive biodiversity-invasibility relationships in shallow lake plant communities. J. Ecol. 2018, 106, 2058–2070. [Google Scholar] [CrossRef]
- Schultz, R.; Dibble, E. Effects of invasive macrophytes on freshwater fish and macroinvertebrate communities: The role of invasive plant traits. Hydrobiologia 2012, 684, 1–14. [Google Scholar] [CrossRef]
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Hiatt, D.; Serbesoff-King, K.; Lieurance, D.; Gordon, D.R.; Flory, S.L. Allocation of invasive plant management expenditures for conservation: Lessons from Florida, USA. Conserv. Sci. Pract. 2019, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hussner, A.; Nehring, S.; Hilt, S. From first reports to successful control: A plea for improved management of alien aquatic plant species in Germany. Hydrobiologia 2014, 737, 321–331. [Google Scholar] [CrossRef]
- Bornette, G.; Puijalon, S. Response of aquatic plants to abiotic factors: A review. Aquat. Sci. 2011, 73, 1–14. [Google Scholar] [CrossRef]
- Cronk, J.K.; Fennessy, M.S. Wetland Plants, 1st ed.; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Wetzel, R.G. Limnology; W.B. Saunders Company: Philidelphia, PA, USA, 1975. [Google Scholar]
- Hudon, C.; Lalonde, S.; Gagnon, P. Ranking the effects of site exposure, plant growth form, water depth, and transparency on aquatic plant biomass. Can. J. Fish. Aquat. Sci. 2000, 57, 31–42. [Google Scholar] [CrossRef]
- Gillefalk, M.; Herzog, C.; Hilt, S. Phosphorus availability and growth of benthic primary producers in littoral lake sediments: Are differences linked to induced bank filtration? Water 2019, 11, 1111. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, H.; Lorke, A.; Peeters, F. Wave-induced variability of the underwater light climate in the littoral zone. Verh. lnt. Verein. Limnol. 2008, 30, 627–632. [Google Scholar] [CrossRef] [Green Version]
- Chambers, P.A. Light and nutrients in the control of aquatic plant community structure. II. In situ observations. J. Ecol. 1987, 75, 621–628. [Google Scholar] [CrossRef]
- Godoy, O.; Levine, J.M. Phenology effects on invasion success: Insights from coupling field experiments to coexistence theory. Ecology 2014, 95, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Torso, K.; Scofield, B.D.; Chess, D.W. Variations in aquatic macrophyte phenology across three temperate lakes in the Coeur d’Alene Basin. Aquat. Bot. 2020, 162, 103209. [Google Scholar] [CrossRef]
- Aiken, S.G.; Newroth, R.; Wiles, I. The biology of Canadian weeds. Myriophyllum spicatum L. Can. J. Plant Sci. 1979, 59, 201–215. [Google Scholar] [CrossRef]
- Eiswerth, M.; Donaldson, S.; Johnson, W. Potential environmental impacts and economic damages of Eurasian watermilfoil (Myriophyllum spicatum) in western Nevada and northeastern California. Weed Technol. 2000, 14, 511–518. [Google Scholar] [CrossRef]
- Barko, J.W.; Smart, M.R. Comparative Influences of Light and Temperature on the Growth and Metabolism of Selected Submersed Freshwater Macrophytes. Ecol. Monogr. 1981, 51, 219–236. [Google Scholar] [CrossRef]
- Smith, C.S.; Barko, J.W. Ecology of Eurasian watermilfoil. J. Aquat. Plant Manag. 1990, 28, 55–64. [Google Scholar]
- Titus, J.E.; Adams, M.S. Coexistence and the comparative light relations of the submersed macrophytes Myriophyllum spicatum L. and Vallisneria americana Michx. Oecologia 1979, 40, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Bolduan, B.R.; Van Eeckhout, G.C.; Quade, H.W.; Gannon, J.E. Potamogeton crispus—The other invader. Lake Reserv. Manag. 1994, 10, 113–125. [Google Scholar] [CrossRef] [Green Version]
- Tobiessen, P.; Snow, P.D. Temperature and light effects on the growth of Potamogeton crispus in Collins Lake, New York State. Can. J. Bot. 1984, 62, 2822–2826. [Google Scholar] [CrossRef]
- Chambers, P.A.; Spence, D.H.N.; Weeks, D.C. Photocontrol of turion formation by Potamogeton Crispus, L. in the laboratory and natural water. New Phytol. 1985, 99, 183–194. [Google Scholar] [CrossRef]
- Woolf, T.E.; Madsen, J. Seasonal biomass and carbohydrate allocation patterns in southern Minnesota curlyleaf pondweed populations. J. Aquat. Plant Manag. 2003, 41, 113–118. [Google Scholar]
- Adamec, L. Ecophysiological characteristics of turions of aquatic plants: A review. Aquat. Bot. 2018, 148, 64–77. [Google Scholar] [CrossRef]
- Carmona, C.P.; de Bello, F.; Mason, N.W.H.; Lepš, J. Traits without borders: Integrating functional diversity across scales. Trends Ecol. Evol. 2016, 31, 382–394. [Google Scholar] [CrossRef]
- Blonder, B. Hypervolume concepts in niche- and trait-based ecology. Ecography (Cop.). 2018, 41, 1441–1455. [Google Scholar] [CrossRef]
- Blonder, B.; Lamanna, C.; Violle, C.; Enquist, B.J. The n-dimensional hypervolume. Glob. Ecol. Biogeogr. 2014, 23, 595–609. [Google Scholar] [CrossRef]
- Perleberg, D.; Radomski, P.; Simon, S.; Carlson, K.; Knopik, J. Minnesota Lake Plant Survey Manual, for Use by MNDNR Fisheries Section and EWR Lake Habitat Program; Minnesota Department of Natural Resources: Brainerd, MN, USA, 2016. [Google Scholar]
- Johnson, J.A.; Jones, A.R.; Newman, R.M. Evaluation of lakewide, early season herbicide treatments for controlling invasive curlyleaf pondweed (Potamogeton crispus) in Minnesota lakes. Lake Reserv. Manag. 2012, 28, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.R.; Johnson, J.A.; Newman, R.M. Effects of repeated, early season, herbicide treatments of curlyleaf pondweed on native macrophyte assemblages in Minnesota lakes. Lake Reserv. Manag. 2012, 28, 364–374. [Google Scholar] [CrossRef] [Green Version]
- Verhoeven, M.R.; Larkin, D.J.; Newman, R.M. Constraining invader dominance: Effects of repeated herbicidal management and environmental factors on curlyleaf pondweed dynamics in 50 Minnesota lakes. Freshw. Biol. 2020, 1–14. [Google Scholar] [CrossRef]
- Nault, M.; Mikulyuk, A.; Hauxwell, J.; Skogerboe, J.; Asplund, T.; Barton, M.; Wagner, K.; Hoyman, T.; Heath, E. Herbicide Treatments in Wisconsin Lakes: Building a Framework for Scientific Evaluation of Large-scale Herbicide Treatments in Wisconsin Lakes. NALMS Lake Line 2012, 32, 19–24. [Google Scholar]
- Kujawa, E.R.; Frater, P.; Mikulyuk, A.; Barton, M.; Nault, M.E.; Van Egeren, S.; Hauxwell, J. Lessons from a decade of lake management: Effects of herbicides on Eurasian watermilfoil and native plant communities. Ecosphere 2017, 8, e01718. [Google Scholar] [CrossRef]
- Frater, P.; Mikulyuk, A.; Barton, M.; Nault, M.; Wagner, K.; Hauxwell, J.; Kujawa, E. Relationships between water chemistry and herbicide efficacy of Eurasian watermilfoil management in Wisconsin lakes. Lake Reserv. Manag. 2016, 33, 1–7. [Google Scholar] [CrossRef]
- Anderson, M.; Clark, M.; Cornett, M.; Hall, K.; Olivero Sheldon, A.; Prince, J. Resilient Sites for Terrestrial Conservation in the Great Lakes and Tallgrass Prairie Region; The Nature Conservancy, Eastern Conservation Science and North America Region: Boston, MA, USA, 2018. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2019. [Google Scholar]
- Madsen, J.D.; Wersal, R.M. A review of aquatic plant monitoring and assessment methods. J. Aquat. Plant Manag. 2017, 55, 1–12. [Google Scholar]
- Chadde, S.W. Wetland Plants of Minnesota: A Complete Guide to the Wetland and Aquatic Plants of the North Star State; Createspace Independent Publishing Platform: Lexington, KY, USA, 2012; ISBN 9781460940341. [Google Scholar]
- Crow, G.E.; Hellquist, C.B. Aquatic and Wetland Plants of Northeastern North America. Volum Pteridophytes, Gymnosperms and Angiosperms: Dicotyledons; University of Wisconsin Press: Madison, WI, USA, 2000. [Google Scholar]
- Crow, G.E.; Hellquist, C.B. Aquatic and Wetland Plants of Northeastern North America. Volume Angiosperms: Monocotyledons; University of Wisconsin Press: Madison, WI, USA, 2000. [Google Scholar]
- Skawinski, P.M. Aquatic Plants of the Upper Midwest; Paul Skawinski: Wausau, WI, USA, 2011; ISBN 978-1-4507-9247-9. [Google Scholar]
- Boyle, B.; Hopkins, N.; Lu, Z.; Raygoza Garay, J.A.; Mozzherin, D.; Rees, T.; Matasci, N.; Narro, M.L.; Piel, W.H.; Mckay, S.J.; et al. The taxonomic name resolution service: An online tool for automated standardization of plant names. BMC Bioinf. 2013, 14, 16. [Google Scholar] [CrossRef] [Green Version]
- Chamberlain, S.A.; Szöcs, E. Taxize: Taxonomic search and retrieval in R. F1000 Res. 2013, 2, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Stadelmann, T.H.; Brezonik, P.L.; Kloiber, S. Seasonal patterns of chlorophyll a and secchi disk transparency in lakes of East-central Minnesota: Implications for design of ground—And satellite-based monitoring programs. Lake Reserv. Manag. 2001, 17, 299–314. [Google Scholar] [CrossRef] [Green Version]
- Preisendorfer, R.W. Eyeball optics of natural waters: Secchi disk science. Limnol. Oceanogr. 1986, 31, 909–926. [Google Scholar] [CrossRef] [Green Version]
- Steele, E.A.; Neuhausser, S. Comparison of methods for measuring visual water clarity. J. North Am. Benthol. Soc. 2002, 21, 326–335. [Google Scholar] [CrossRef]
- Megard, R.O.; Settles, J.C.; Boyer, H.A.; Combs, W.S.J. Light, Secchi disks, and trophic states. Limnol. Oceanogr. 1980, 25, 373–377. [Google Scholar] [CrossRef]
- Van Duin, E.H.S.; Blom, G.; Los, F.J.; Maffione, R.; Zimmerman, R.; Cerco, C.F.; Dortch, M.; Best, E.P.H. Modeling underwater light climate in relation to sedimentation, resuspension, water quality and autotrophic growth. Hydrobiologia 2001, 444, 25–42. [Google Scholar] [CrossRef]
- McMaster, G.S.; Wilhelm, W.W. Growing degree-days: One equation, two interpretations. Agric. For. Meteorol. 1997, 87, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Cross, H.Z.; Zuber, M.S. Prediction of flowering dates in maize based on different methods of estimating thermal units. Agron. J. 1972, 64, 351–355. [Google Scholar] [CrossRef]
- Russelle, M.P.; Wilhelm, W.W.; Olson, R.A.; Power, J.F. Growth analysis based on degree days. Crop Sci. 1984, 24, 28–32. [Google Scholar] [CrossRef] [Green Version]
- Luedeling, E.; Kunz, A.; Blanke, M.M. Identification of chilling and heat requirements of cherry trees-a statistical approach. Int. J. Biometeorol. 2013, 57, 679–689. [Google Scholar] [CrossRef] [Green Version]
- NOAA. National Centers for Environmental Information Global Surface Hourly [2001–2018]; NOAA: Washington, DC, USA, 2018.
- Boissezon, A.; Auderset Joye, D.; Garcia, T. Temporal and spatial changes in population structure of the freshwater macroalga Nitellopsis obtusa (Desv.) J.Groves. Bot. Lett. 2018, 165, 103–114. [Google Scholar] [CrossRef]
- Carmona, C.P.; Bello, F.; Mason, N.W.H.; Lepš, J. Trait probability density (TPD): Measuring functional diversity across scales based on TPD with R. Ecology 2019, 100, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Blonder, B.; Lamanna, C.; Violle, C.; Enquist, B.J. Using n-dimensional hypervolumes for species distribution modelling: A response to Qiao et al. (). Glob. Ecol. Biogeogr. 2017, 26, 1071–1075. [Google Scholar] [CrossRef] [Green Version]
- Blonder, B.; Morrow, C.B.; Maitner, B.; Harris, D.J.; Lamanna, C.; Violle, C.; Enquist, B.J.; Kerkhoff, A.J. New approaches for delineating n -dimensional hypervolumes. Methods Ecol. Evol. 2018, 9, 305–319. [Google Scholar] [CrossRef] [Green Version]
- Blonder, B. Do hypervolumes have holes? Am. Nat. 2016, 187, E93–E105. [Google Scholar] [CrossRef]
- Zedler, J.B.; Kercher, S. Causes and consequences of invasive plants in wetlands: Opportunities, opportunists, and outcomes. CRC. Crit. Rev. Plant Sci. 2004, 23, 431–452. [Google Scholar] [CrossRef]
- MacDougall, A.S.; Gilbert, B.; Levine, J.M. Plant invasions and the niche. J. Ecol. 2009, 97, 609–615. [Google Scholar] [CrossRef]
- Mitchell, C.E.; Agrawal, A.A.; Bever, J.D.; Gilbert, G.S.; Hufbauer, R.A.; Klironomos, J.N.; Maron, J.L.; Morris, W.F.; Parker, I.M.; Power, A.G.; et al. Biotic interactions and plant invasions. Ecol. Lett. 2006, 9, 726–740. [Google Scholar] [CrossRef]
- Seabloom, E.W.; Borer, E.T.; Boucher, V.L.; Burton, R.S.; Cottingham, K.L.; Goldwasser, L.; Gram, W.K.; Kendall, B.E.; Micheli, F. Competition, seed limitation, disturbance, and reestablishment of California native annual forbs. Ecol. Appl. 2003, 13, 575–592. [Google Scholar] [CrossRef] [Green Version]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 436–443. [Google Scholar] [CrossRef]
- Hilt, S.; Henschke, I.; Rücker, J.; Nixdorf, B. Can submerged macrophytes influence turbidity and trophic state in deep lakes? Suggestions from a case study. J. Environ. Qual. 2010, 39, 725. [Google Scholar] [CrossRef]
- James, W.F.; Dechamps, A.; Turyk, N.; Mcginley, P. Contribution of Potamogeton Crispus Decay to the Phosphorus Budget of McGinnis Lake, Wisconsin; U.S. Army Engineer Research and Development Center: Vicksburg, MS, USA, 2007. [Google Scholar]
- Boylen, C.W.; Eichler, L.W.; Madsen, J.D. Loss of native aquatic plant species in a community dominated by Eurasian watermilfoil. Hydrobiologia 1999, 415, 207–211. [Google Scholar] [CrossRef]
- Scheele, B.C.; Foster, C.N.; Banks, S.C.; Lindenmayer, D.B. Niche contractions in declining species: Mechanisms and consequences. Trends Ecol. Evol. 2017, 32, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.; Newman, M. A comparison of two methods for sampling biomass of aquatic plants. J. Aquat. Plant Manag. 2011, 49, 1–8. [Google Scholar]
- Marshall, C.T.; Peters, R.H. General patterns in the seasonal development of chlorophyll a for temperate lakes. Limnol. Oceanogr. 1989, 34, 856–867. [Google Scholar] [CrossRef]
- Titus, J.; Goldstein, R.A.; Adams, M.S.; Mankin, J.B.; O’neill, R.V.; Weiler, P.R., Jr.; Shugart, H.H.; Booth, R.S. A production model for Myriophyllum spicatum L. Ecology 1997, 56, 1129–1138. [Google Scholar] [CrossRef]
- Grace, J.B.; Wetzel, R.G. The production biology of Eurasian watermilfoil (Myriophyllum spicatum L.): A Review. J. Aquat. Plant. Manag. 1978, 16, 1–11. [Google Scholar]
- Adams, M.S.; McCracken, M.D. Seasonal Production of the Myriophyllum Component of the Littoral of Lake Wingra, Wisconsin. J. Ecol. 1974, 62, 457. [Google Scholar] [CrossRef]
- Galatowitsch, S.M.; Anderson, N.O.; Ascher, P.D. Invasiveness in wetland plants in temperate North America. Wetlands 1999, 19, 733–755. [Google Scholar] [CrossRef]
- Valley, R.D.; Bremigan, M.T. Effects of macrophyte bed architecture on largemouth bass foraging: Implications of exotic macrophyte invasions. Trans. Am. Fish. Soc. 2002, 131, 234–244. [Google Scholar] [CrossRef]
- Wu, J.; Cheng, S.; Liang, W.; He, F.; Wu, Z. Effects of sediment anoxia and light on turion germination and early growth of Potamogeton crispus. Hydrobiologia 2009, 628, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Gioria, M.; Osborne, B.A. Resource competition in plant invasions: Emerging patterns and research needs. Front. Plant Sci. 2014, 5, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Hautier, Y.; Niklaus, P.; Hector, A. Competition for light causes plant biodiversity loss after eutrophication. Science 2009, 324, 636–638. [Google Scholar] [CrossRef] [Green Version]
- Kolar, C.S.; Lodge, D.M. Progress in invasion biology: Predicting invaders. Trends Ecol. Evol. 2001, 16, 199–204. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verhoeven, M.R.; Glisson, W.J.; Larkin, D.J. Niche Models Differentiate Potential Impacts of Two Aquatic Invasive Plant Species on Native Macrophytes. Diversity 2020, 12, 162. https://doi.org/10.3390/d12040162
Verhoeven MR, Glisson WJ, Larkin DJ. Niche Models Differentiate Potential Impacts of Two Aquatic Invasive Plant Species on Native Macrophytes. Diversity. 2020; 12(4):162. https://doi.org/10.3390/d12040162
Chicago/Turabian StyleVerhoeven, Michael R., Wesley J. Glisson, and Daniel J. Larkin. 2020. "Niche Models Differentiate Potential Impacts of Two Aquatic Invasive Plant Species on Native Macrophytes" Diversity 12, no. 4: 162. https://doi.org/10.3390/d12040162
APA StyleVerhoeven, M. R., Glisson, W. J., & Larkin, D. J. (2020). Niche Models Differentiate Potential Impacts of Two Aquatic Invasive Plant Species on Native Macrophytes. Diversity, 12(4), 162. https://doi.org/10.3390/d12040162