Loss of Mitochondrial Genetic Diversity in Overexploited Mediterranean Swordfish (Xiphias gladius, 1759) Population

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and DNA Extraction
2.2. Mitochondrial DNA Amplification and Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Palko, B.J.; Beardsley, G.L.; Richards, W.J. Synopsis of the biology of the swordfish, Xiphias gladius (L.). U.S. Dep. Commer. NOAA Tech. Rep. NMFS Circ. 1981, 441, 21. [Google Scholar]
- Neilson, J.; Arocha, F.; Cass-Calay, S.; Mejuto, J.; Ortiz, M.; Scott, G.; Smith, C.; Travassos, P.; Tserpes, G.; Andrushchenko, I. The recovery of Atlantic swordfish: The comparative roles of the regional fisheries management organization and species biology. Rev. Fish. Sci. 2013, 21, 59–97. [Google Scholar] [CrossRef]
- Abascal, F.J.; Mejuto, J.; Quintans, M.; García-Cortés, B.; Ramos-Cartelle, A. Tracking of the broadbill swordfish, Xiphias gladius, in the central and eastern North Atlantic. Fish. Res. 2015, 162, 20–28. [Google Scholar] [CrossRef]
- Arocha, F. Swordfish Reproduction in the Atlantic Ocean: An Overview. GCR 2007, 19. [Google Scholar] [CrossRef] [Green Version]
- Marisaldi, L.; Basili, D.; Candelma, M.; Sesani, V.; Pignalosa, P.; Gioacchini, G.; Carnevali, O. Maturity assignment based on histology-validated macroscopic criteria: Tackling the stock decline of the Mediterranean swordfish (Xiphias gladius). Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 303–314. [Google Scholar] [CrossRef]
- Macías, D.; Hattour, A.; De la Serna, J.M.; Gómez-Vives, M.J.; Godoy, D. Reproductive characteristics of swordfish (Xiphias gladius) caught in the southwestern Mediterranean during 2003. Collect. Vol. Sci. Pap. ICCAT 2005, 58, 454–469. [Google Scholar]
- Anon. Report for Biennial Period, 2018–19 PART I (2018)—Volume 2; International Commission for the Conservation of Atlantic Tunas: Madrid, Spain, 2019; pp. 182–192. [Google Scholar]
- Bremer, J.A.; Mejuto, J.; Gómez-Márquez, J.; Pla-Zanuy, C.; Viñas, J.; Marques, C.; Hazin, F.; Griffiths, M.; Ely, B.; García-Cortés, B. Genetic population structure of Atlantic swordfish: Current status and future directions. Collect. Vol. Sci. Pap. ICCAT 2007, 61, 107–118. [Google Scholar]
- Viñas, J.; Bremer, J.A.; Pla, C. Inter-oceanic genetic differentiation among albacore (Thunnus alalunga) populations. Mar. Biol. 2004, 145, 225–232. [Google Scholar] [CrossRef]
- Bremer, J.R.A.; Viñas, J.; Mejuto, J.; Ely, B.; Pla, C. Comparative phylogeography of Atlantic bluefin tuna and swordfish: The combined effects of vicariance, secondary contact, introgression, and population expansion on the regional phylogenies of two highly migratory pelagic fishes. Mol. Phylogenetics Evol. 2005, 36, 169–187. [Google Scholar] [CrossRef]
- Šegvić-Bubić, T.; Marrone, F.; Grubišić, L.; Izquierdo-Gomez, D.; Katavić, I.; Arculeo, M.; Brutto, S.L. Two seas, two lineages: How genetic diversity is structured in Atlantic and Mediterranean greater amberjack Seriola dumerili Risso, 1810 (Perciformes, Carangidae). Fish. Res. 2016, 179, 271–279. [Google Scholar] [CrossRef]
- Bremer, J.R.A.; Mejuto, J.; Greig, T.W.; Ely, B. Global population structure of the swordfish (Xiphias gladius L.) as revealed by analysis of the mitochondrial DNA control region. J. Exp. Mar. Biol. Ecol. 1996, 197, 295–310. [Google Scholar] [CrossRef]
- Rosel, P.E.; Block, B.A. Mitochondrial control region variability and global population structure in the swordfish, Xiphias gladius. Mar. Biol. 1996, 125, 11–22. [Google Scholar] [CrossRef]
- Chow, S.; Okamoto, H.; Uozumi, Y.; Takeuchi, Y.; Takeyama, H. Genetic stock structure of the swordfish (Xiphias gladius) inferred by PCR-RFLP analysis of the mitochondrial DNA control region. Mar. Biol. 1997, 127, 359–367. [Google Scholar] [CrossRef]
- Muths, D.; Grewe, P.; Jean, C.; Bourjea, J. Genetic population structure of the Swordfish (Xiphias gladius) in the southwest Indian Ocean: Sex-biased differentiation, congruency between markers and its incidence in a way of stock assessment. Fish. Res. 2009, 97, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Reeb, C.A.; Arcangeli, L.; Block, B.A. Structure and migration corridors in Pacific populations of the Swordfish Xiphius gladius, as inferred through analyses of mitochondrial DNA. Mar. Biol. 2000, 136, 1123–1131. [Google Scholar] [CrossRef]
- Viñas, J.; Pérez-Serra, A.; Vidal, O.; Alvarado Bremer, J.R.; Pla, C. Genetic differentiation between eastern and western Mediterranean swordfish revealed by phylogeographic analysis of the mitochondrial DNA control region. ICES J. Mar. Sci. 2010, 67, 1222–1229. [Google Scholar] [CrossRef] [Green Version]
- Bremer, J.R.A.; Baker, A.J.; Mejuto, J. Mitochondrial DNA control region sequences indicate extensive mixing of swordfish (Xiphias gladius) populations in the Atlantic Ocean. Can. J. Fish. Aquat. Sci. 1995, 52, 1720–1732. [Google Scholar] [CrossRef]
- Pujolar, J.M.; Roldan, M.I.; Pla, C. A genetic assessment of the population structure of swordfish (Xiphias gladius) in the Mediterranean Sea. J. Exp. Mar. Biol. Ecol. 2002, 276, 19–29. [Google Scholar] [CrossRef]
- Kotoulas, G.; Mejuto, J.; Antoniou, A.; Kasapidis, P.; Tserpes, G.; Piccinetti, C.; Peristeraki, P.; Garcia-Cortes, B.; Oikonomaki, K.; De la Serna, J.M. Global genetic structure of swordfish (Xiphias gladius) as revealed by microsatellite DNA markers. Collect. Vol. Sci. Pap. ICCAT 2007, 61, 79–88. [Google Scholar]
- Xiphias gladius (errata version published in 2016). The IUCN Red List of Threatened Species 2011: e.T23148A88828055. Available online: https://dx.doi.org/10.2305/IUCN.UK.2011-2.RLTS.T23148A9422329.en. (accessed on 17 December 2017).
- Pinsky, M.L.; Palumbi, S.R. Meta-analysis reveals lower genetic diversity in overfished populations. Mol. Ecol. 2014, 23, 29–39. [Google Scholar] [CrossRef]
- Frankham, R. Genetics and extinction. Biol. Conserv. 2005, 126, 131–140. [Google Scholar] [CrossRef]
- Spielman, D.; Brook, B.W.; Frankham, R. Most species are not driven to extinction before genetic factors impact them. Proc. Natl. Acad. Sci. USA 2004, 101, 15261–15264. [Google Scholar] [CrossRef] [Green Version]
- Hauser, L.; Adcock, G.J.; Smith, P.J.; Ramírez, J.H.B.; Carvalho, G.R. Loss of microsatellite diversity and low effective population size in an overexploited population of New Zealand snapper (Pagrus auratus). Proc. Natl. Acad. Sci. USA 2002, 99, 11742–11747. [Google Scholar] [CrossRef] [Green Version]
- Turner, T.F.; Wares, J.P.; Gold, J.R. Genetic effective size is three orders of magnitude smaller than adult census size in an abundant, estuarine-dependent marine fish (Sciaenops ocellatus). Genetics 2002, 162, 1329–1339. [Google Scholar]
- Hutchinson, W.F.; Oosterhout, C.; Rogers, S.I.; Carvalho, G.R. Temporal analysis of archived samples indicates marked genetic changes in declining North Sea cod (Gadus morhua). Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 2125–2132. [Google Scholar] [CrossRef] [Green Version]
- Allendorf, F.W.; England, P.R.; Luikart, G.; Ritchie, P.A.; Ryman, N. Genetic effects of harvest on wild animal populations. Trends Ecol. Evol. 2008, 23, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2003, 1, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 565–567. [Google Scholar] [CrossRef] [PubMed]
- Heck, K.L., Jr.; van Belle, G.; Simberloff, D. Explicit calculation of the rarefaction diversity measurement and the determination of sufficient sample size. Ecology 1975, 56, 1459–1461. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (H ill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Bandelt, H.-J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [PubMed]
- Kuhner, M.K. LAMARC 2.0: Maximum likelihood and Bayesian estimation of population parameters. Bioinformatics 2006, 22, 768–770. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, K.; Yandell, D.W. How sensitive is PCR-SSCP? Hum. Mutat. 1993, 2, 338–346. [Google Scholar] [CrossRef]
- Garibaldi, F. Effects of the introduction of the mesopelagic longline on catches and size structure of swordfish in the Ligurian sea (western Mediterranean). Collect. Vol. Sci. Pap. ICCAT 2015, 71, 2006–2014. [Google Scholar]
- Garibaldi, F.; Lanteri, L. Notes about a tagged/recaptured swordfish in the Ligurian Sea (western Mediterranean). Collect. Vol. Sci. Pap. ICCAT 2017, 74, 1354–1361. [Google Scholar]
- Bertolino, F.; Camolese, C.; Dell’Aquila, M.; Mariani, A.; Valastro, M. Swordfish (Xiphias gladius L.) fisheries using mesopelagic longline in the Mediterranean Sea by Italian fishing fleet. Collect. Vol. Sci. Pap. ICCAT 2015, 71, 2073–2078. [Google Scholar]
- Abid, N.; Laglaoui, A.; Arakrak, A.; Bakkali, M. The reproductive biology of swordfish (Xiphias gladius) in the Strait of Gibraltar. J. Mar. Biol. Assoc. U. K. 2019, 99, 649–659. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Sadovy, Y.; Reynolds, J.D. Extinction vulnerability in marine populations. Fish Fish. 2003, 4, 25–64. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Jennings, S.; Rogers, S.I.; Maxwell, D.L. Threat and decline in fishes: An indicator of marine biodiversity. Can. J. Fish. Aquat. Sci. 2006, 63, 1267–1275. [Google Scholar] [CrossRef]
- Kenchington, E.L. 14 The Effects of Fishing on Species and Genetic Diversity. Responsible Fish. Mar. Ecosyst. 2003, 235. [Google Scholar]
- Smith, P.J.; Francis, R.; McVeagh, M. Loss of genetic diversity due to fishing pressure. Fish. Res. 1991, 10, 309–316. [Google Scholar] [CrossRef]
- Ruggeri, P.; Splendiani, A.; Di Muri, C.; Fioravanti, T.; Santojanni, A.; Leonori, I.; De Felice, A.; Biagiotti, I.; Carpi, P.; Arneri, E. Coupling demographic and genetic variability from archived collections of European anchovy (Engraulis encrasicolus). PLoS ONE 2016, 11, e0151507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. The state of world fisheries and aquaculture 2016. Contributing to Food Security and Nutrition for all; FAO: Rome, Italy, 2016. [Google Scholar]




{kind=link}
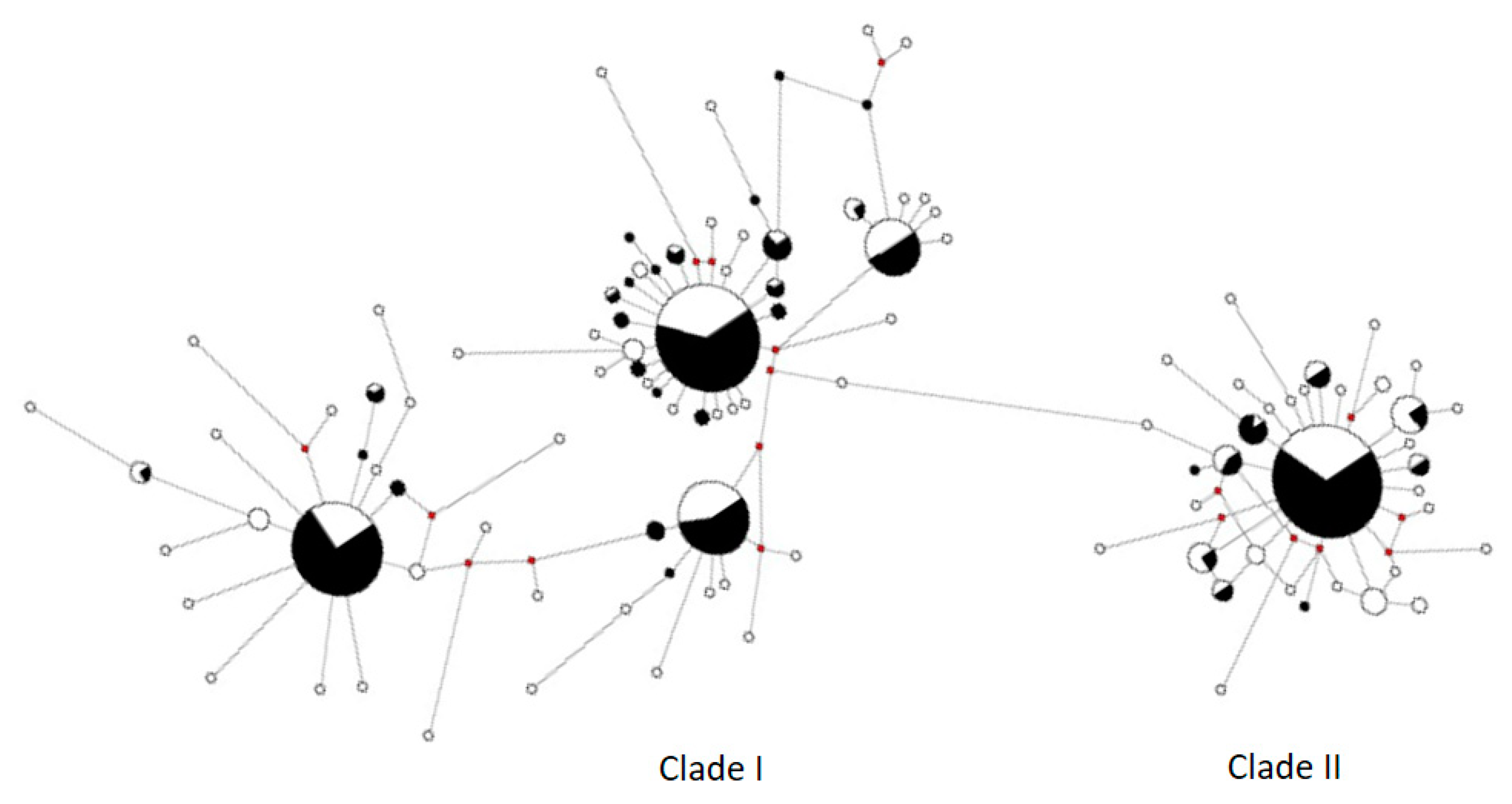{kind=link}
{kind=link}
| Sampling Area (FAO Fishing Area/Geographical Subarea) | Sampling Date | n | Mean LJFL (cm) (s.d.) |
|---|---|---|---|
| Balearic Sea (GSA 5, 6) | July 2016–August 2016–September 2016 | 84 | 122.1 (22,2) |
| September 2019 | |||
| Southern Sicily (GSA 15, 16) | July 2016 | 59 | 133.7 (32.0) |
| June 2017–July 2017 | |||
| June 2018 | |||
| Aegean Sea (GSA 22) | August 2016 | 17 | 124.2 (21.0) |
| South Adriatic Sea (GSA 18) | September 2016 | 62 | 113.5 (16.6) |
| Tyrrhenian Sea (GSA 10) | May 2017 | 16 | 120.8 (23.8) |
| Sardinian Sea (GSA 11.2) | June 2017–October 2017 | 49 | 125.0 (32.6) |
| May 2018–Jun 2018 | |||
| Total | 287 |
| Genbank Accession Number | 2 | 9 | 12 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 46 | 60 | 67 | 77 | 83 | 91 | 92 | 101 | 110 | 111 | 118 | 120 | 122 | 123 | 125 | 126 | 127 | 132 | 134 | 135 | 138 | 158 | 159 | 160 | 161 | 165 | 166 | 172 | 173 | 178 | 180 | 186 | 187 | 190 | 194 | 217 | 257 | 262 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AY650768 | G | T | A | T | A | C | A | - | - | - | - | C | A | A | C | A | T | T | A | G | T | T | A | A | C | T | G | T | T | A | T | C | A | T | T | T | A | A | A | T | C | C | - | T | G | G | T | A | G |
| MN652597 | . | . | . | . | . | . | . | T | A | C | A | . | . | . | . | . | . | . | . | . | . | C | . | . | . | . | . | . | . | . | . | T | . | . | . | . | . | . | . | . | . | . | - | . | . | . | . | . | . |
| AY650778 | . | . | . | . | . | . | . | - | - | - | - | . | . | . | . | . | . | . | . | . | . | C | . | . | . | . | . | . | . | . | . | T | . | . | . | . | . | . | . | . | . | . | - | . | . | . | . | . | . |
| AY650860 | . | . | . | . | . | . | . | - | - | - | - | . | . | . | . | . | . | . | . | . | . | C | . | . | . | . | . | . | . | . | . | T | G | . | . | . | . | . | . | . | . | . | - | . | . | . | . | . | . |
| MN652598 | A | . | . | . | . | . | . | - | - | - | - | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | - | . | . | . | . | . | . |
| MN652599 | . | . | . | . | . | . | . | - | - | - | - | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | G | . | . | . | . | . | - | . | . | . | . | . | . |
| EU827759 | . | . | . | . | . | . | . | - | - | - | - | . | . | . | . | G | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | - | . | . | . | . | . | . |
| MN652600 | . | . | . | . | . | . | . | T | A | C | A | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | G | . | . | . | . | . | . | . | . | . | - | . | . | . | . | . | . |
| MN652601 | . | . | . | . | . | . | . | - | - | - | - | . | . | . | . | . | . | . | . | . | . | . | . | . | . | C | . | . | . | . | . | . | G | . | . | . | . | . | . | . | . | . | - | . | . | . | . | . | . |
| AY650836 | . | . | . | . | . | . | . | - | - | - | - | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | G | . | . | . | . | . | . | . | . | . | - | . | . | . | . | . | . |
| AY650821 | . | . | . | . | . | . | . | - | - | - | - | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | C | . | . | . | . | . | . | . | . | . | - | . | . | . | . | . | . |
| MN652602 | . | . | . | . | . | . | . | - | - | - | - | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | C | . | . | . | . | . | . | . | . | - | . | . | . | . | . | . |
| AY650858 | . | . | . | . | . | . | . | - | - | - | - | . | . | G | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | - | . | . | . | . | . | . |
| MN652603 | . | . | . | . | . | . | . | - | - | - | - | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | G | . | . | . | . | - | . | . | . | . | . | . |
| MN652604 | . | . | . | . | . | . | . | - | - | - | - | . | . | . | . | . | . | . | . | . | C | . | . | . | T | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | - | . | . | . | . | . | . |
| MN652605 | . | . | . | . | . | . | . | - | - | - | - | . | . | . | . | . | . | . | . | . | . | . | . | . | T | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | - | . | . | . | . | . | . |
| MN652606 | . | . | . | . | . | . | . | - | - | - | - | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | G | . | . | . | . | . | . | . | . | . | . | . | . | - | . | . | . | . | . | . |
| MN652607 | . | . | . | . | . | . | . | - | - | - | - | . | . | . | T | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | - | . | . | . | . | . | . |
| MN652608 | . | - | . | . | . | . | . | - | - | - | - | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | C | . | . | T | . | . | . | . | . | . | . | . | G | T | - | . | A | . | . | . | . |
| AY650781 | . | . | . | . | . | . | . | - | - | - | - | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | C | . | . | T | . | . | . | . | . | . | . | . | G | T | - | . | A | . | . | . | . |
| MN652609 | . | . | . | . | . | . | . | - | - | - | - | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | A | . | C | . | . | T | . | . | . | . | . | . | . | . | G | T | - | . | A | . | . | . | . |
| AY650763 | . | . | - | . | . | . | . | - | - | - | - | . | . | . | T | . | . | . | . | . | . | . | . | . | T | . | A | . | C | . | . | T | . | . | . | . | . | . | . | C | . | . | - | . | A | . | . | . | A |
| MN652595 | . | . | - | . | . | . | . | - | - | - | - | . | G | . | T | . | . | . | . | . | . | . | . | . | T | . | A | . | C | . | . | T | . | . | . | . | . | . | . | C | . | . | - | . | A | . | . | . | A |
| AY650855 | . | . | - | . | . | . | . | T | A | C | A | . | . | . | T | . | . | . | . | . | . | . | . | . | T | . | A | . | C | . | . | T | . | . | . | . | . | . | . | C | . | . | - | . | A | . | C | . | A |
| MN652596 | . | . | - | . | . | . | . | - | - | - | - | . | . | . | T | . | . | . | . | . | . | . | . | . | T | . | A | . | C | . | . | T | . | . | . | . | . | . | . | C | . | . | - | . | . | . | . | . | A |
| AY650861 | . | . | - | . | . | . | . | - | - | - | - | . | . | . | T | . | . | . | . | . | . | . | . | . | T | . | A | . | C | . | . | T | . | . | . | . | . | . | G | C | . | . | - | . | . | . | . | . | . |
| AY650809 | . | . | . | - | - | - | - | - | - | - | - | T | . | . | T | . | . | . | T | C | . | . | G | G | . | . | A | C | C | G | . | T | G | . | C | . | . | . | . | . | . | T | C | . | . | . | . | G | A |
| MN652610 | . | . | . | - | - | - | - | - | - | - | - | T | . | . | T | . | . | . | T | C | . | . | G | G | . | . | A | C | C | G | . | T | G | . | C | . | . | . | . | . | . | T | C | C | . | . | . | G | A |
| AY650762 | . | . | . | - | - | - | - | - | - | - | - | T | . | . | T | . | . | . | T | C | . | . | G | G | . | . | A | C | C | G | . | T | G | . | C | . | . | . | . | . | . | T | C | . | . | A | . | G | A |
| AY650814 | . | . | . | - | - | - | - | - | - | - | - | T | . | . | T | . | . | . | T | C | . | . | G | G | . | . | A | C | C | G | . | T | G | . | C | C | . | . | . | . | . | T | C | . | . | A | . | G | A |
| AY650805 | . | . | . | - | - | - | - | - | - | - | - | T | . | . | T | . | . | . | T | C | . | . | G | G | . | . | A | C | C | G | . | T | G | . | C | . | . | G | . | . | . | T | C | . | . | A | . | G | A |
| AY650761 | . | . | . | - | - | - | - | - | - | - | - | T | . | . | T | . | . | C | T | C | . | . | G | G | . | . | A | C | C | G | . | T | G | . | C | . | . | . | . | . | . | T | C | . | . | A | . | G | A |
| AY650829 | . | . | . | - | - | - | - | - | - | - | - | T | . | . | T | . | . | . | T | T | . | . | G | G | . | . | A | C | C | G | . | T | G | . | C | . | . | . | . | . | . | T | C | . | . | A | . | G | A |
| EU827771 | . | . | . | - | - | - | - | - | - | - | - | T | . | . | T | . | . | . | T | C | . | . | G | G | . | . | A | . | C | G | C | T | G | . | C | . | . | . | . | . | . | T | C | . | . | A | . | G | A |
| MN652611 | . | . | . | - | - | - | - | - | - | - | - | T | . | . | T | . | C | . | T | C | . | . | G | G | . | . | A | C | C | G | . | T | G | . | C | . | . | . | . | . | . | T | C | . | . | A | . | . | A |
| EU827787 | . | . | . | - | - | - | - | - | - | - | - | T | . | . | T | . | . | . | T | T | . | . | G | G | . | . | A | C | C | G | . | T | G | . | C | . | . | . | . | . | . | T | C | . | . | A | . | G | . |
| Title | N | H | h (s.d.) | π (s.d.) | θ (95% C.I.) |
|---|---|---|---|---|---|
| Viñas et al., 2010 | |||||
| Clade I + II | 251 | 93 | 0.946 (0.007) | 0.160 (0.080) | 0.142 (0.121–0.188) |
| Clade I | 156 | 60 | 0.910 (0.014) | 0.034 (0.020) | 0.076 (0.060–0.107) |
| Clade II | 95 | 33 | 0.861 (0.031) | 0.006 (0.004) | 0.044 (0.0.29–0.062) |
| This study | |||||
| Clade I + II | 286 | 36 | 0.844 (0.010) | 0.130 (0.070) | 0.033 (0.028–0.044) |
| Clade I | 190 | 26 | 0.793 (0.018) | 0.029 (0.015) | 0.019 (0.015–0.030) |
| Clade II | 96 | 10 | 0.418 (0.063) | 0.003 (0.002) | 0.010 (0.006–0.017) |
| Source of Variation | Variance Component | Percentage of Variation | Fixation Index (ФST) | p-Value |
|---|---|---|---|---|
| Whole dataset | ||||
| Among temporal samples | 0.008 | 1.77 | 0.0177 | 0.0000 |
| Within samples | 0.446 | 98.23 | ||
| Clade I | ||||
| Among temporal samples | 0.008 | 1.85 | 0.0184 | 0.0005 |
| Within samples | 0.423 | 98.15 | ||
| Clade II | ||||
| Among temporal samples | 0.044 | 12.10 | 0.121 | 0.0000 |
| Within samples | 0.319 | 87.90 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Righi, T.; Splendiani, A.; Fioravanti, T.; Casoni, E.; Gioacchini, G.; Carnevali, O.; Caputo Barucchi, V. Loss of Mitochondrial Genetic Diversity in Overexploited Mediterranean Swordfish (Xiphias gladius, 1759) Population. Diversity 2020, 12, 170. https://doi.org/10.3390/d12050170
Righi T, Splendiani A, Fioravanti T, Casoni E, Gioacchini G, Carnevali O, Caputo Barucchi V. Loss of Mitochondrial Genetic Diversity in Overexploited Mediterranean Swordfish (Xiphias gladius, 1759) Population. Diversity. 2020; 12(5):170. https://doi.org/10.3390/d12050170
Chicago/Turabian StyleRighi, Tommaso, Andrea Splendiani, Tatiana Fioravanti, Elia Casoni, Giorgia Gioacchini, Oliana Carnevali, and Vincenzo Caputo Barucchi. 2020. "Loss of Mitochondrial Genetic Diversity in Overexploited Mediterranean Swordfish (Xiphias gladius, 1759) Population" Diversity 12, no. 5: 170. https://doi.org/10.3390/d12050170
APA StyleRighi, T., Splendiani, A., Fioravanti, T., Casoni, E., Gioacchini, G., Carnevali, O., & Caputo Barucchi, V. (2020). Loss of Mitochondrial Genetic Diversity in Overexploited Mediterranean Swordfish (Xiphias gladius, 1759) Population. Diversity, 12(5), 170. https://doi.org/10.3390/d12050170