Drivers of Foliar Fungal Endophytic Communities of Kudzu (Pueraria montana var. lobata) in the Southeast United States


Abstract
:1. Introduction
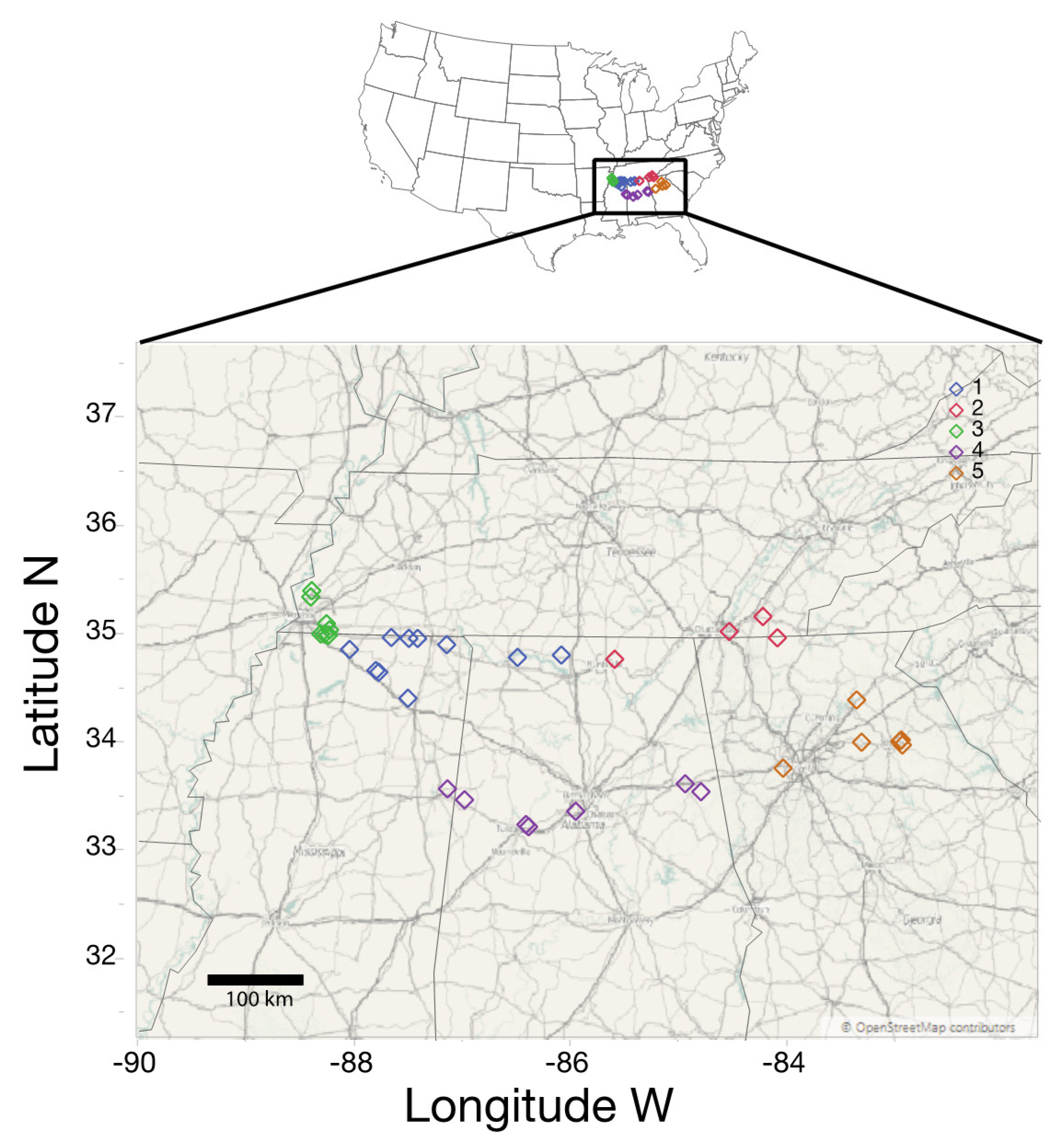
2. Materials and Methods
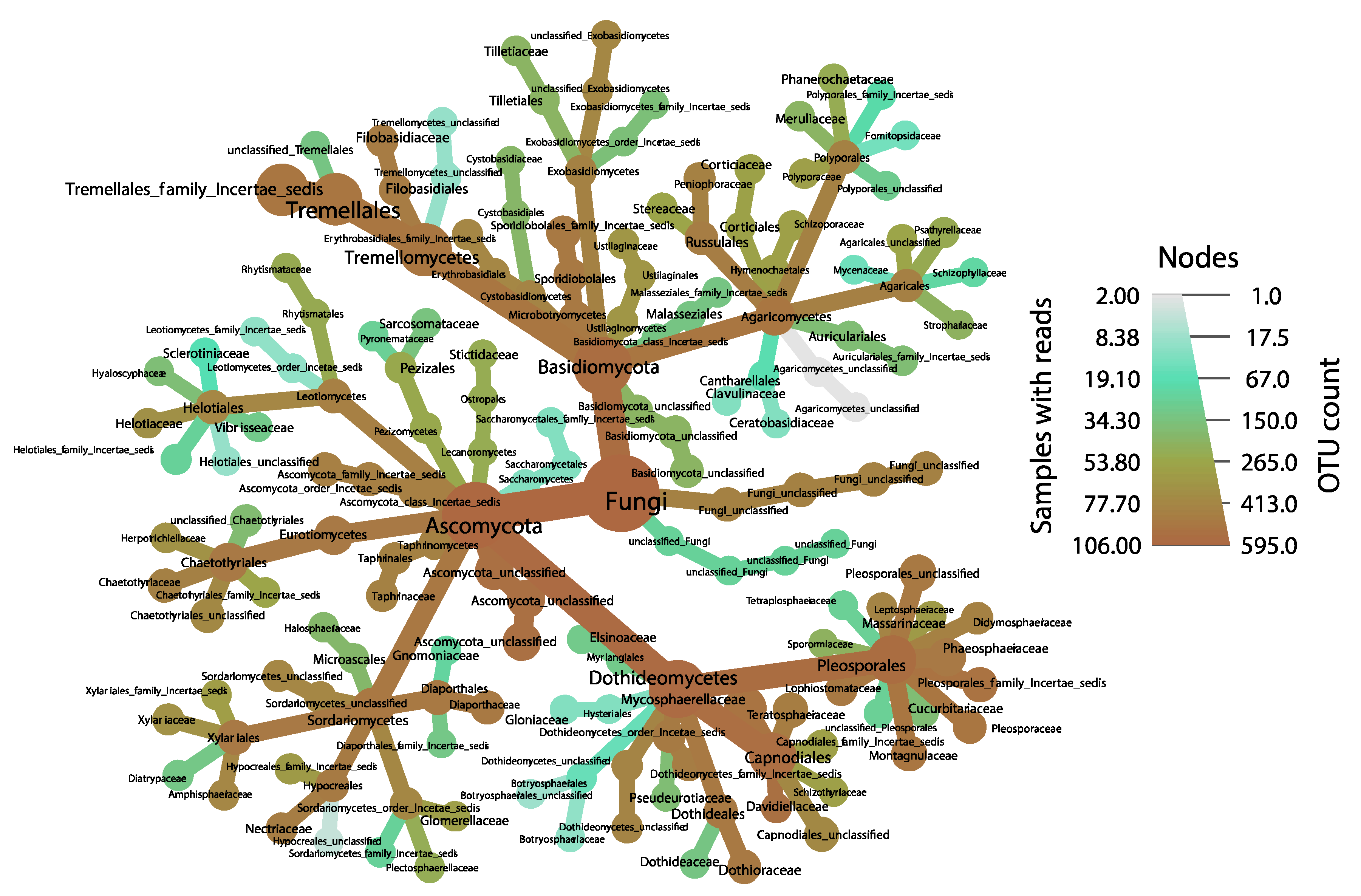
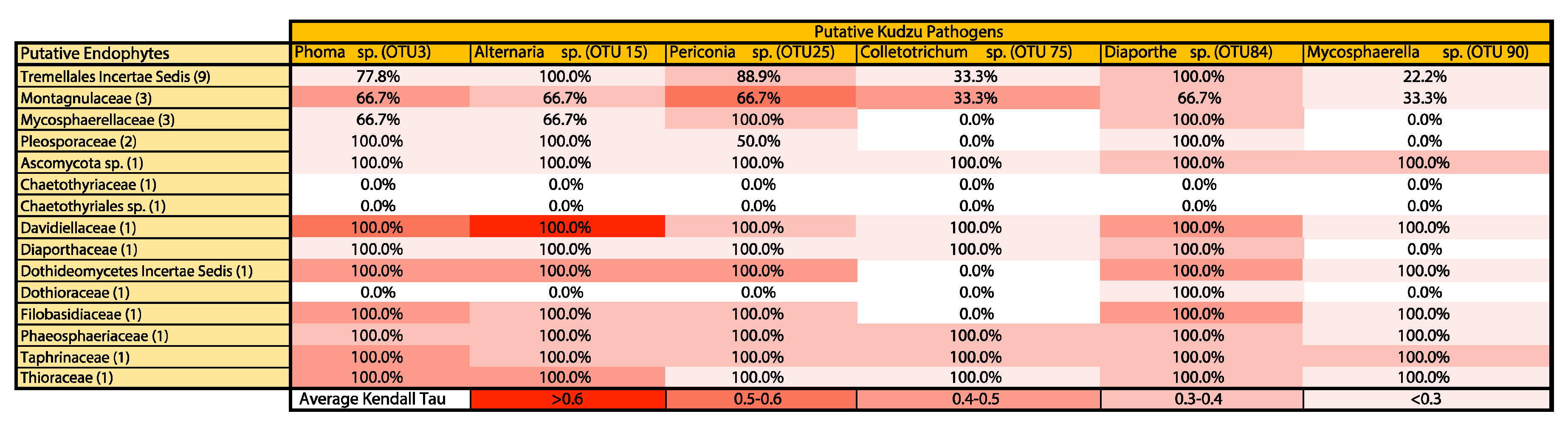
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- National Invasive Species Council. Meeting the Invasive Species Challenge; National Invasive Management Plan; U.S. Department of the Interior: Washington, DC, USA, 2001; p. 80.
- U.S. Department of Agriculture. APHIS Factsheet, Invasive Species; U.S. Department of Agriculture: Washington, DC, USA, 1999. Available online: https://www.invasive.org/publications/aphis/invasive.pdf (accessed on 13 January 2020).
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic Invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Williams, D.G.; Mack, R.N.; Black, R.A. Ecophysiology of introduced Pennisetum setaceum on Hawaii: The role of phenotypic plasticity. Ecology 1995, 76, 1569–1580. [Google Scholar] [CrossRef]
- Bais, H.P.; Vepachedu, R.; Gilroy, S.; Callaway, R.M.; Vivanco, J.M. Allelopathy and plant invasion: From molecules and genes to species interactions. Science 2003, 301, 1377–1380. [Google Scholar] [CrossRef]
- Martin, P.A.; Newton, A.C.; Bullock, J.M. Impacts of invasive plants on carbon pools depend on both species’ traits and local climate. Ecology 2017, 98, 1026–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stinson, K.A. Plant Suppresses the Growth of Native Tree Seedlings by Disrupting Belowground Mutualisms. PLoS Biol. 2006, 4, e140. [Google Scholar] [CrossRef] [PubMed]
- Thakur, M.P.; van der Putten, W.H.; Cobben, M.M.P.; Kleunen, M.; Geisen, S. Microbial invasions in terrestrial Ecosystems. Nat. Rev. Microbiol. 2019, 17, 621–631. [Google Scholar] [CrossRef]
- Callaway, R.M.; Cipollini, D.; Barto, K.; Thelen, G.C.; Hallett, S.G.; Prati, D.; Klironomos, J. Novel weapons: Invasive plant suppresses fungal mutualists in America but not in its native Europe. Ecology 2008, 89, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Pisula, N.; Meiners, S. Relative allelopathic potential of invasive plant species in a young disturbed woodland. J. Torrey Bot. Soc. 2010, 137, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Kong, C.H.; Xuan, T.D.; Khanh, T.D.; Tran, H.D.; Trung, N.T. Allelochemicals and signaling chemicals in plants. Molecules 2019, 24, 2737. [Google Scholar] [CrossRef] [Green Version]
- Chengxu, W.Z.; Mingxing, C.; Xuhui, Q.; Bo, Q. Review on allelopathy of exotic invasive plants. Procedia Eng. 2011, 18, 240–246. [Google Scholar] [CrossRef] [Green Version]
- Hawkes, C.V.; Wren, I.F.; Herman, D.J.; Firestone, M.K. Plant invasion alters nitrogen cycling by modifying the soil nitrifying community. Ecol. Lett. 2005, 8, 976–985. [Google Scholar] [CrossRef]
- Gibbons, S.M.; Lekberg, Y.; Mummey, D.L.; Sangwan, N.; Ramsey, P.W.; Gilbert, J.A. Invasive Plants rapidly Reshape soil properties in a grassland ecosystem. MSystems 2017, 2, e178-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiTommaso, A.; Zhong, Q.; Clements, D.R. Identifying Climate Change as a factor in the establishment and persistence of invasive weeds in agricultural crops. In Invasive Species and Global Climate Change; Ziska, L.H., Dukes, J.S., Eds.; CABI Publishing: Wallingford, UK, 2014; pp. 253–270. [Google Scholar]
- Theoharides, K.A.; Dukes, J.S. Plant Invasion Across space and Time: Factors Affecting nonindigenous species success during four stages of invasion. New Phytol. 2007, 176, 256–273. [Google Scholar] [CrossRef]
- Jurskis, V. Plant invasions: Symptoms and contributors rather than causes of environmental degradation. Forests 2012, 3, 896–902. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G. Effects of Exotic Plant invasions on soil nutrient cycling processes. Ecosystems 2003, 6, 503–523. [Google Scholar] [CrossRef]
- Qi, S.S.; Dai, Z.C.; Zhai, D.L.; Chen, S.C.; Si, C.C.; Huang, P.; Wang, R.P.; Zhong, Q.X.; Du, D.L. Curvilinear effects of Invasive plants on plant diversity: Plant community invaded by Sphagneticola trilobata. PLoS ONE 2014, 9, e113964. [Google Scholar] [CrossRef]
- Arim, M.; Sebastián, R.A.; Paula, E.N.; Mauricio, L.; Pablo, A.M. Spread dynamics of invasive species. Proc. Natl. Acad. Sci. USA 2006, 103, 374–378. [Google Scholar] [CrossRef] [Green Version]
- Postic, J.; Cosic, J.; Vrandecic, K.; Jurkovic, D.; Saleh, A.A.; Leslie, J.F. Diversity of fusarium species isolated from weeds and plant debris in Croatia. J. Phytopathol. 2012, 160, 76–81. [Google Scholar] [CrossRef]
- Wisler, G.C.; Norris, R.F. Interactions between weeds and cultivated plants as related to management of plant pathogens. Weed Sci. 2005, 53, 914–917. [Google Scholar] [CrossRef]
- Wang, M.N.; Chen, X.M. Barberry does not function as an alternate host for Puccinia striiformis f. sp. tritici in the US Pacific Northwest due to teliospore degradation and barberry phenology. Plant Dis. 2015, 99, 1500–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christiano, R.S.C.; Scherm, H. Quantitative aspects of the spread of Asian Soybean Rust in the Southeastern United States, 2015–2016. Phytopathology 2007, 97, 1428–1433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hershman, D.E.; Bachi, P.R.; Harmon, C.L.; Harmon, P.F.; Palm, M.E.; McKemy, J.M.; Zeller, K.A.; Levy, L. First report of soybean rust Caused by Phakopsora pachyrhizi on Kudzu (Pueraria montana var. lobata) in Kentucky. Dis. Notes 2006, 90, 834. [Google Scholar] [CrossRef] [PubMed]
- Fabiszewski, A.M.; Umbanhowar, J.; Mitchell, C.S. Modeling landscape-scale pathogen spillover between domesticated and wild hosts: Asian Soybean Rust and Kudzu. Ecol. Appl. 2010, 20, 582–592. [Google Scholar] [CrossRef] [Green Version]
- Shurtleff, W.; Aoyagi, A. The Book of Kudzu: A Culinary and Healing Guide; Autumn Press: Brookline, MA, USA, 1997; p. 102. [Google Scholar]
- Sturkie, D.G.; Grimes, J.C. Kudzu Its Value and Use in Alabama; Agricultural Experiment Station of the Alabama Polytechnic Institute: Auburn, AL, USA, 1939; pp. 1–20. [Google Scholar]
- Forseth, I.N.; Innis, A.F. Kudzu (Pueraria montana): History, physiology, and ecology combine to make a major ecosystem threat. Crit. Rev. Plant Sci. 2004, 23, 401–413. [Google Scholar] [CrossRef]
- Miller, J.H.; Miller, K.V. Forest Plants of the Southeast and Their Wildlife Uses; Southern Weed Science Society: Champaign, IL, USA, 1999; pp. 284–285. [Google Scholar]
- Miller, J.H. Kudzu eradication and management. In Kudzu: The Vine to Love or Hate; Hoots, D., Baldwin, J., Eds.; Suntop Press: Kodak, TN, USA, 1996; pp. 34–40. [Google Scholar]
- Harrington, T.B.; Rader-Dixon, L.T.; Taylor, J.W. Kudzu (Pueraria montana) community responses to herbicides, burning, and high-density Loblolly Pine. Weed Sci. 2003, 51, 965–974. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.H.; Edwards, B. Kudzu: Where did it come from? And how can we stop it? South. J. Appl. For. 1983, 7, 165–169. [Google Scholar] [CrossRef] [Green Version]
- Blaustein, R.J. Kudzu’s invasion into Southern United States life and culture. In The Great Reshuffling: Human Dimensions of Invasive Species; McNeeley, J.A., Ed.; IUCN, The World Conservation Union, Gland: Cambridge, UK, 2001; pp. 55–62. [Google Scholar]
- Britton, K.O.; Orr, D.; Sun, J.H. Kudzu. In Biological Control of Invasive Plants in the Eastern United States; FHTET-2002-04; Van Driesche, R., Ed.; USDA Forest Service Publication: Morgantown, WV, USA, 2002; pp. 325–330. [Google Scholar]
- Sun, J.H.; Li, Z.C.; Jewett, D.K.; Britton, K.O.; Ye, W.H.; Ge, X.J. Genetic diversity of Pueraria lobata (Kudzu) and closely related taxa as revealed by inter-simple sequence repeat analysis. Weed Res. 2005, 45, 255–260. [Google Scholar] [CrossRef]
- Bentley, K.; Mauricio, R. High Degree of Clonal reproduction and lack of large-scale geographic patterning mark the introduced range of the invasive vine, kudzu (Pueraria montana var. lobata) in North America. Am. J. Bot. 2016, 103, 1499–1507. [Google Scholar] [CrossRef] [Green Version]
- Westbrooks, R. Invasive Plants: Changing the Landscape of America; Federal Interagency Committee for the Management of Noxious and Exotic Weeds: Washington, DC, USA, 1998; p. 109. [Google Scholar]
- Heckel, C.D. Impacts of Exotic Invasive Vines on the Ecology and Reproduction of the Endangered Trillium reliquum. Master’s Thesis, Department of Biology, Georgia Southern University, Statesboro, GA, USA, 2004. [Google Scholar]
- Hickman, J.E.; Wu, S.L.; Mickley, L.J.; Lerdau, M.T. Kudzu (Pueraria montana) invasion doubles emissions of nitric oxide and increases Ozone pollution. Proc. Natl. Acad. Sci. USA 2010, 107, 10115–10119. [Google Scholar] [CrossRef] [Green Version]
- Boyette, C.D.; Walker, H.L.; Abbas, H.K. Biological control of Kudzu (Pueraria lobata) with an isolate of Myrothecium verrucaria. Biocontrol Sci. Technol. 2002, 12, 75–82. [Google Scholar] [CrossRef]
- Weaver, M.A.; Jin, X.; Hoagland, R.E.; Boyette, C.D. Improved bioherbicidal efficacy by Myrothecium verrucaria via spray adjuvants or herbicide mixtures. Biol. Control 2009, 50, 150–156. [Google Scholar] [CrossRef]
- Weaver, M.A.; Boyette, C.D.; Hoagland, R.E. Management of Kudzu by the bioherbicide, Myrothecium verrucaria, herbicides and integrated control programs. Biocontrol Sci. Technol. 2016, 26, 136–140. [Google Scholar] [CrossRef]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttiläd, A.M.; Compant, S.; Andrea Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [Green Version]
- Stone, J.K.; Bacon, C.W.; White, J.F., Jr. An Overview of Endophytic Microbes: Endophytism Defined. In Microbial Endophytes; Bacon, C.W., White, J.F., Jr., Eds.; Marcel Dekker: New York, NY, USA, 2000; pp. 3–29. [Google Scholar]
- Newcombe, G.; Harding, A.; Ridout, M.; Busby, P.E. A Hypothetical Bottleneck in the Plant Microbiome. Front. Microbiol. 2018, 9, 1645. [Google Scholar] [CrossRef] [Green Version]
- Hodgson, S.; Cates, C.; Hodgson, J.; Morley, N.J.; Sutton, B.C.; Gange, A.C. Vertical transmission of fungal endophytes is widespread in forbs. Ecol. Evol. 2014, 4, 1199–1208. [Google Scholar] [CrossRef] [Green Version]
- Owen, N.L.; Hundley, N. Endophytes-The chemical synthesizers inside Plants. Sci. Prog. 2004, 87, 79–99. [Google Scholar] [CrossRef]
- Frank, A.C.; Saldierna Guzmán, J.P.; Shay, J.E. Transmission of bacterial endophytes. Microorganisms 2017, 5, 70. [Google Scholar] [CrossRef] [Green Version]
- Scott, R.I.; Chard, J.M.; Hocart, M.J.; Lennard, J.H.; Graham, D.C. Penetration of Potato Tuber Lenticels by Bacteria in Relation to Biological Control of Blackleg Disease. Potato Res. 1996, 39, 333–344. [Google Scholar] [CrossRef]
- Gagné, S.; Richard, C.; Rouseau, H.; Antoun, H. Xylem-residing bacteria in Alfalfa roots. Can. J. Microbiol. 1987, 33, 996–1000. [Google Scholar] [CrossRef]
- Marques, J.M.; da Silva, T.F.; Vollu, R.E.; Blank, A.F.; Smalla, K.; Seldin, L. Bacterial endophytes of sweet potato tuberous roots Affected by the plant genotype and growth stage. Appl. Soil Ecol. 2015, 96, 273–281. [Google Scholar] [CrossRef]
- Ding, T.; Melcher, U. Influences of plant species, season and location on leaf endophytic bacterial communities of non-cultivated plants. PLoS ONE 2016, 11, e0150895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Yang, J.; Wang, E.; Li, B.; Yuan, H. Effects of growth stage and fulvic acid on the diversity and dynamics of endophytic bacterial community in Stevia rebaudiana Bertoni leaves. Front. Microbiol. 2015, 6, 867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christian, N.; Sullivan, C.; Visser, N.D.; Clay, K. Plant host and geographic location drive endophyte community composition in the face of perturbation. Microb. Ecol. 2016, 72, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.; Johnson, C.; Santos-Medellin, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl. Acad. Sci. USA 2015, 112, E911–E920. [Google Scholar] [CrossRef] [Green Version]
- Jorge, M.G.; Elba, E.; Valentín, P.; Javier, D.J. Factors influencing endophytic communities in poplar plantations. Silva Fenn. 2011, 45, 169–180. [Google Scholar]
- Rodriguez-Blanco, A.; Sicardi, M.; Frioni, L. Plant genotype and nitrogen fertilization effects on abundance and diversity of diazotrophic bacteria associated with maize (Zea mays L.). Biol. Fertil. Soils 2015, 51, 391–402. [Google Scholar] [CrossRef]
- Seghers, D.; Wittebolle, L.; Top, E.M.; Verstraete, W.; Siciliano, S.D. Impact of agricultural practices on the Zea mays L. endophytic community. Appl. Environ. Microbiol. 2004, 3, 1475–1482. [Google Scholar] [CrossRef] [Green Version]
- Richardson, D.M.; Allsopp, N.; D’Antonio, C.M.; Milton, S.J.; Rejmanek, M. Plant invasions—The role of mutualisms. Biol. Rev. 2000, 75, 65–93. [Google Scholar] [CrossRef]
- Reinhart, K.O.; Callaway, R.M. Soil biota facilitate exotic acer invasion in Europe and North America. Ecol. Appl. 2004, 14, 1737–1745. [Google Scholar] [CrossRef] [Green Version]
- Reinhart, K.O.; Callaway, R.M. Soil Biota and invasive plants. New Phytol. 2006, 170, 445–457. [Google Scholar] [CrossRef]
- Mönchgesang, S.; Strehmel, N.; Schmidt, S.; Westphal, L.; Taruttis, F.; Müller, E.; Herklotz, S.; Neumann, S.; Scheel, D. Natural variation of root exudates in Arabidopsis thaliana—Linking metabolomic and genomic data. Sci. Rep. 2016, 6, 29033. [Google Scholar] [CrossRef] [PubMed]
- Klironomos, J.N. Feedback with Soil biota contributes to plant rarity and invasiveness in communities. Nature 2002, 417, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Si, C.C.; Liu, X.Y.; Wang, C.Y.; Wang, L.; Dai, Z.C.; Qi, S.S. Different degrees of plant invasion significantly affect the richness of the soil fungal community. PLoS ONE 2013, 8, e85490. [Google Scholar] [CrossRef] [PubMed]
- Busby, P.E.; Ridout, M.; Newcombe, G. Fungal endophytes: Modifiers of plant disease. Plant Mol. Biol. 2016, 90, 645–655. [Google Scholar] [CrossRef]
- Busby, P.E.; Peay, K.G.; Newcombe, G. Common foliar fungi of Populus trichocarpa modify Melampsora rust disease severity. New Phytol. 2016, 209, 1681–1692. [Google Scholar] [CrossRef] [Green Version]
- Kurose, D.; Furuya, N.; Tsuchiya, K.; Tsushima, S.; Evans, H.C. Endophytic fungi associated with Fallopia japonica (Polygonaceae) in Japan and their interactions with Puccinia polygoniamphibii var. tovariae, a candidate for classical biological control. Fungal Biol. 2012, 116, 785–791. [Google Scholar] [CrossRef]
- Bailey, B.; Bae, H.; Strem, M.; Crozier, J.; Thomas, S.; Samuels, G.; Vinyard, B.; Holmes, K. Antibiosis, mycoparasitism, and colonization success for endophytic Trichoderma isolates with biological control potential in Theobroma cacao. Biol. Control 2008, 46, 24–35. [Google Scholar] [CrossRef]
- Kauppinen, M.; Saikkonen, K.; Helander, M.; Pirttilä, A.M.; Wäli, P.R. Epichloë grass endophytes in sustainable agriculture. Nat. Plants 2016, 2, 15224. [Google Scholar] [CrossRef]
- Greenfield, M.; Pareja, R.; Ortiz, V.; Gómez-Jiménez, M.I.; Vega, F.E.; Parsa, S.A. Novel method to scale up fungal endophyte isolations. Biocontrol Sci. Technol. 2015, 25, 1208–1212. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.P.; Leopold, D.R.; Busby, P.E. Protocols for investigating the leaf mycobiome using high-throughput DNA sequencing. In Plant Pathogenic Fungi and Oomycetes: Methods and Protocols; Ma, W., Wolpert, T., Eds.; Humana Press: New York, NY, USA, 2018; pp. 39–51. [Google Scholar]
- Zhu, J.; Tremblay, N.; Liang, Y. Comparing SPAD and atLEAF values for chlorophyll assessment in crop species. Can. J. Soil Sci. 2012, 92, 645–648. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to imageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Mordukhovich, I.; Beyea, J.; Herring, A.H.; Hatch, M.; Stellman, S.D.; Teitelbaum, S.L.; Richardson, D.B.; Millikan, R.C.; Engel, L.S.; Shantakumar, S.; et al. Vehicular Traffic-Related Polycyclic Aromatic Hydrocarbon Exposure and Breast Cancer Incidence: The Long Island Breast Cancer Study Project (LIBCSP). Environ. Health Perspect. 2016, 124, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Imperato, V.; Kowalkowski, L.; Portillo-Estrada, M.; Gawronski, S.W.; Vangronsveld, J.; Thijs, S. Characterisation of the Carpinus betulus L. phyllomicrobiome in urban and forest areas. Front. Microbiol. 2019, 10, 1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barge, E.G.; Leopold, D.R.; Peay, K.G.; Newcombe, G.; Busby, P.E. Differentiating spatial from environmental effects of foliar fungal communities of Populus trichocarpa. J. Biogeogr. 2019, 46, 2001–2011. [Google Scholar] [CrossRef]
- Brown, S.P.; Ungerer, M.C.; Jumpponen, A. A Community of clones: Snow algae are diverse communities of spatially structured clones. Int. J. Plant Sci. 2016, 177, 432–439. [Google Scholar] [CrossRef] [Green Version]
- Ihrmark, K.; Bödeker, I.T.M.; Martinez, K.C.; Friberg, H.; Kubartova, A.; Schenck, J.; Strid, Y.; Stenlid, J.; Brandström-Durling, M.; Clemmensen, K.E.; et al. New primers to amplify the fungal ITS2 region–evaluation by 454-sequencing of artificial and natural communities. FEMS Microbiol. Ecol. 2012, 82, 666–677. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Brown, S.P.; Jumpponen, A. Contrasting primary successional trajectories of fungi and bacteria in retreating glacier soils. Mol. Ecol. 2014, 23, 481–497. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Opensource, platform independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microb. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- Huse, S.M.; Welch, D.W.; Morrison, H.G.; Sogin, M.L. Ironing out the wrinkles in the rare biosphere through improves OTU clustering. Environ. Microbiol. 2010, 12, 1889–1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile Open source Tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Toward a unified Paradigm for Sequence-Based Identification of Fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.P.; Veach, A.M.; Ridgon-Huss, A.R.; Grond, K.; Lickteig, S.K.; Lothamer, K.; Oliver, A.K.; Jumpponen, A. Scraping the bottom of the barrel: Are rare high throughput sequences artifacts? Fungal Ecol. 2015, 13, 221–225. [Google Scholar] [CrossRef] [Green Version]
- Oliver, A.K.; Brown, S.P.; Callaham, M.A.; Jumpponen, A. Polymerase matters: Non-proofreading enzymes inflate fungal community richness Estimates by up to 15%. Fungal Ecol. 2015, 15, 86–89. [Google Scholar] [CrossRef] [Green Version]
- Foster, Z.; Shorpton, T.; Grunwald, N. Metacoder: An R package for visualization and manipulation of community taxonomic Diversity Data. PLoS Comput. Biol. 2017, 13, e1005404. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.J. A New Method for non-parametric Multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package; R Package Version 2.4-0; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Anderson, M.J.; Walsh, D.C.I. Permanova, anosim, and the mantel test in the face of heterogeneous dispersion: What null hypothesis are you testing. Ecol. Monogr. 2013, 83, 557–574. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.W.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Shilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Stergiopoulos, I.; Gordon, T.R. Cryptic fungal infections: The hidden agenda of plant pathogens. Front. Plant Sci. 2014, 5, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Précigout, P.-A.; Claessen, D.; Makowski, D.; Robert, C. Does the latent period of leaf fungal pathogens reflect their trophic type? A meta-analysis of biotrophs, hemibiotrophs, and necrotrophs. Phytopathology 2020, 110, 354–361. [Google Scholar]
- Jumpponen, A.; Jones, K.L. Massively Parallel 454 sequencing indicates hyper diverse fungal communities in temperate Quercus macrocarpa phyllosphere. New Phytol. 2009, 184, 438–448. [Google Scholar] [CrossRef]
- Izuno, A.; Kanzaki, M.; Artchawakom, T.; Wachrinrat, C.; Isagi, Y. Vertical Structure of Phyllosphere Fungal communities in a tropical forest in Thailand uncovered by high-throughput sequencing. PLoS ONE 2016, 11, e0166669. [Google Scholar] [CrossRef]
- Schoch, C.L.; Shoemaker, R.A.; Seifert, K.A.; Hambelton, S.; Spatafora, J.W.; Crous, P.W. A multigene phylogeny of the Dothideomycetes using four nuclear loci. Mycologia 2006, 98, 1041–1052. [Google Scholar] [CrossRef]
- Ohm, R.A.; Feau, N.; Henrissat, B.; Schoch, C.L.; Horwitz, B.A.; Barry, K.W.; Condon, B.J.; Alex, C.; Copeland, A.C.; Dhillon, B.; et al. Diverse Lifestyles and Strategies of Plant Pathogenesis Encoded in the Genomes of Eighteen Dothideomycetes Fungi. PLoS Pathog. 2012, 8, e1003037. [Google Scholar] [CrossRef] [Green Version]
- Dube, H.C. An Introduction to Fungi; Vikas Publishing House: Delhi, India, 1990; p. 608. [Google Scholar]
- Zhao, Y.; Liu, X.Z.; Bai, F.Y. Four new species of Tremella (Tremellales, Basidiomycota) based on morphology and DNA sequence data. MycoKeys 2019, 47, 75–95. [Google Scholar] [CrossRef]
- Gams, W.; Diederich, P.; Põldmaa, K. Fungicolous fungi. In Biodiversity of Fungi: Inventory and Monitoring Methods; Mueller, G.M., Bills, G.F., Foster, M.S., Eds.; Elsevier Academic Press: Cambridge, MA, USA, 2004; pp. 343–392. [Google Scholar]
- Zielinska, B.; Sagebiel, J.; Arnott, W.P.; Rogers, C.F.; Kelly, K.E.; Wagner, D.A.; Lighty, J.S.; Sarofi, A.F.; Palmer, G. Phase and size distribution of polycyclic aromatic hydrocarbons in diesel and gasoline vehicle emissions. Environ. Sci. Technol. 2004, 38, 2557–2567. [Google Scholar] [CrossRef]
- Potin, O.; Veignie, E.; Rafin, C. Biodegradation of polycyclic aromatic hydrocarbons (PAH) by Cladosporium sphaerospermum isolated from an aged PAH contaminated soil. FEMS Microbiol. Ecol. 2004, 51, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Wuyts, K.; Smets, W.; Lebeer, S.; Samson, R. Green infrastructure and atmospheric pollution shape diversity and composition of phyllosphere bacterial communities in an urban landscape. FEMS Microbiol. Ecol. 2020, 96, fiz173. [Google Scholar] [CrossRef]
- Postma, J.W.M.; Olsson, P.A.; Falkengren-Grerup, U. Root colonisation by arbuscular mycorrhizal, fine endophytic and dark septate fungi across a pH gradient in acid beech forests. Soil Biol. Biochem. 2007, 39, 400–408. [Google Scholar] [CrossRef]
- Kemmitt, S.J.; Wright, D.; Goulding, K.W.T.; Jones, D.L. pH regulation of carbon and nitrogen dynamics in two agricultural soils. Soil Biol. Biochem. 2006, 38, 898–911. [Google Scholar] [CrossRef]
- Darlison, J.; Mogren, L.; Rosberg, A.K.; Grudén, M.; Minet, A.; Liné, C.; Mieli, M.; Bengtsson, T.; Håkansson, A.; Uhlig, E.; et al. Leaf mineral content govern microbial community structure in the phyllosphere of Spinach (Spinacia oleracea) and Rocket (Diplotaxis tenuifolia). Sci. Total Environ. 2019, 675, 501–512. [Google Scholar] [CrossRef]
- González-Teuber, M.; Vilo, C.; Bascuñán-Godoy, L. Molecular characterization of endophytic fungi associated with the roots of Chenopodium quinoa inhabiting the Atacama Desert, Chile. Genom. Data 2017, 11, 109–112. [Google Scholar] [CrossRef]
- Brundrett, M.C. Coevolution of roots and mycorrhizas of land plants. New Phytol. 2002, 154, 275–304. [Google Scholar] [CrossRef]
- Berg, G.; Smalla, K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microb. Ecol. 2009, 68, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wu, X.; Chen, T.; Wang, W.; Liu, G.; Zhang, W.; Li, S.; Wang, M.; Zhao, C.; Zhou, H.; et al. Plant phenotypic traits eventually shape its microbiota: A common garden test. Front. Microbiol. 2018, 9, 2479. [Google Scholar] [CrossRef] [Green Version]
- Bakker, P.A.H.M.; Berendsen, R.L.; Doornbos, R.F.; Wintermans, P.C.A.; Pieterse, C.M.J. The Rhizosphere revisited: Root microbiomics. Front. Plant Sci. 2013, 4, 165. [Google Scholar] [CrossRef] [Green Version]
- Burns, J.H.; Anacker, B.L.; Strauss, S.Y.; Burke, D.J. Soil microbial community variation correlates most strongly with plant species identity, followed by soil chemistry, spatial location and plant genus. AoB Plants 2015, 7, plv030. [Google Scholar] [CrossRef] [Green Version]
- Docherty, K.M.; Borton, H.M.; Espinosa, N.; Gebhardt, M.; Gil-Loaiza, J.; Gutknecht, J.L.M.; Maes, P.W.; Mott, B.M.; Parnell, J.J.; Purdy, G.; et al. Key edaphic properties largely explain temporal and geographic variation in soil microbial communities across four biomes. PLoS ONE 2015, 10, e0135352. [Google Scholar] [CrossRef]
- Helander, M.L.; Neuvonen, S.; Sieber, T.; Petrini, O. Simulated acid rain affects birch leaf endophyte populations. Microb. Ecol. 1993, 26, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Malinowski, D.P.; Alloush, G.A.; Belesky, D.P. Leaf endophyte Neotyphodium coenophialum modifies mineral uptake in tall fescue. Plant Soil. 2000, 227, 116–126. [Google Scholar] [CrossRef]
- D’Jonsiles, M.F.; Carmarán, C.C.; Robles, C.A.; Ceriani-Nakamurakare, E.D.; Novas, M.V. Mycorrhizal colonization and soil parameters affected by foliar endophytes in Jatropha curcas L. J. Soil Sci. Plant Nutr. 2019, 19, 332–341. [Google Scholar]
- Compant, S.; Saikkonen, K.; Mitter, B.; Campisano, A.; Mercado-Blanco, J. Editorial special issue: Soil, plants, and endophytes. Plant Soil 2016, 405, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cai, G.; Schneider, R.W. Population Structure of Cercospora kikuchii, the causal agent of Cercospora leaf blight and purple seed stain in soybean. Phytopathology 2008, 98, 823–829. [Google Scholar] [CrossRef]
- Rotem, J. The Genus Alternaria, Biology, Epidemiology and Pathogenicity; APS Press: St. Paul, MN, USA, 1994; p. 326. [Google Scholar]
- Tu, J.C. Epidemiology of Anthracnose caused by Colletotrichum lindemuthianum on white and stems with lesions were cut into Bean (Phaseolus vulgaris) in Southern Ontario: Survival of the pathogen. Plant Dis. 1983, 67, 402–404. [Google Scholar] [CrossRef] [Green Version]
- Higgins, B.B. “Halo Spot” of Beans and Kudzu. In Bulletin of the Georgia Agricultural Experimental Station; No. 161; Bulletin: Athens, GA, USA, 1930; p. 20. [Google Scholar]
- Pan, J.J.; May, G. Fungal-fungal associations affect the assembly of endophyte communities in Maize (Zea mays). Microb. Ecol. 2009, 58, 668–678. [Google Scholar] [CrossRef]
- Morin, L.; Auld, B.A.; Brown, J.F. Interaction between Puccinia xanthii and facultative parasitic fungi on Xanthium occidentale. Biol. Control 1993, 3, 288–295. [Google Scholar] [CrossRef]
- Leone, M.R.; Lackner, G.; Silipo, A.; Lanzetta, R.; Molinaro, A.; Hertweck, C. An unusual galactofuranose lipopolysaccharide that ensures the intracellular survival of toxin-producing bacteria in their fungal host. Angew. Chem. Int. Ed. Engl. 2010, 49, 7476–7480. [Google Scholar] [CrossRef]
- Partida-Martinez, L.P.; Hertweck, C. Pathogenic fungus harbours endosymbiotic bacteria for toxin production. Nature 2005, 437, 884–888. [Google Scholar] [CrossRef]
- Lackner, G.; Moebius, N.; Hertweck, C. Endofungal bacterium controls its host by an hrp type III secretion system. ISME J. 2011, 5, 252–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Tested Variable. | Diversity (1-D) | Richness (Sobs) | Evenness (ED) |
|---|---|---|---|
| Chlorophyll (mg/cm2) | NS | NS | NS |
| Genotype | NS | NS | NS |
| Leaf area (cm2) | NS | NS | NS |
| Location | F4,88 = 12.884, p < 0.001 | F4,88 = 7.280, p < 0.001 | F4,88 = 10.423, p < 0.001 |
| Nitrate (ppm) | t = −3.27, p = 0.0015 | NS | t = −2.21, p = 0.0292 |
| Potassium (ppm) | NS | NS | NS |
| Soil pH | NS | t = 2.50, p = 0.0143 | NS |
| Traffic Intensity | F4,89 = 4.059, p = 0.0045 | F4,89 = 3.530, p = 0.010 | NS |
| Environmental Factor | Percentage of Significant OTUs (Positive %, Negative %) |
|---|---|
| Traffic Intensity | 37% |
| Location Cluster | 29% |
| Kudzu Genotype | 12% |
| Soil pH | 10% (8%, 2%) |
| Leaf Chlorophyll | 10% (10%, 0%) |
| Leaf Area | 9% (4%, 5%) |
| Stem and Leaf K+ | 5% (0%, 5%) |
| Stem and Leaf NO3− | 2% (2%, 0%) |
| Model Factor | Df | Pseudo-F | R2 | p Value |
|---|---|---|---|---|
| Location | 4 | 5.729 | 0.15299 | 0.001 |
| Genotype | 5 | 3.8823 | 0.12959 | 0.001 |
| Traffic Intensity | 4 | 4.7323 | 0.12637 | 0.001 |
| Soil pH | 1 | 6.1513 | 0.04107 | 0.001 |
| Leaf NO3− | 1 | 4.7655 | 0.03182 | 0.001 |
| Chlorophyll | 1 | 4.4625 | 0.02979 | 0.001 |
| Leaf Area | 1 | 4.0106 | 0.02678 | 0.001 |
| Leaf K+ | 1 | 2.7078 | 0.01808 | 0.003 |
| Residuals | 95 | 0.44351 | ||
| Total | 113 | 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahrtash, M.; Brown, S.P. Drivers of Foliar Fungal Endophytic Communities of Kudzu (Pueraria montana var. lobata) in the Southeast United States. Diversity 2020, 12, 185. https://doi.org/10.3390/d12050185
Shahrtash M, Brown SP. Drivers of Foliar Fungal Endophytic Communities of Kudzu (Pueraria montana var. lobata) in the Southeast United States. Diversity. 2020; 12(5):185. https://doi.org/10.3390/d12050185
Chicago/Turabian StyleShahrtash, Maryam, and Shawn P. Brown. 2020. "Drivers of Foliar Fungal Endophytic Communities of Kudzu (Pueraria montana var. lobata) in the Southeast United States" Diversity 12, no. 5: 185. https://doi.org/10.3390/d12050185
APA StyleShahrtash, M., & Brown, S. P. (2020). Drivers of Foliar Fungal Endophytic Communities of Kudzu (Pueraria montana var. lobata) in the Southeast United States. Diversity, 12(5), 185. https://doi.org/10.3390/d12050185