Patterns of Co-occurrence of Rare and Threatened Species in Winter Arable Plant Communities of Italy


Abstract
:
1. Introduction
2. Materials and Methods
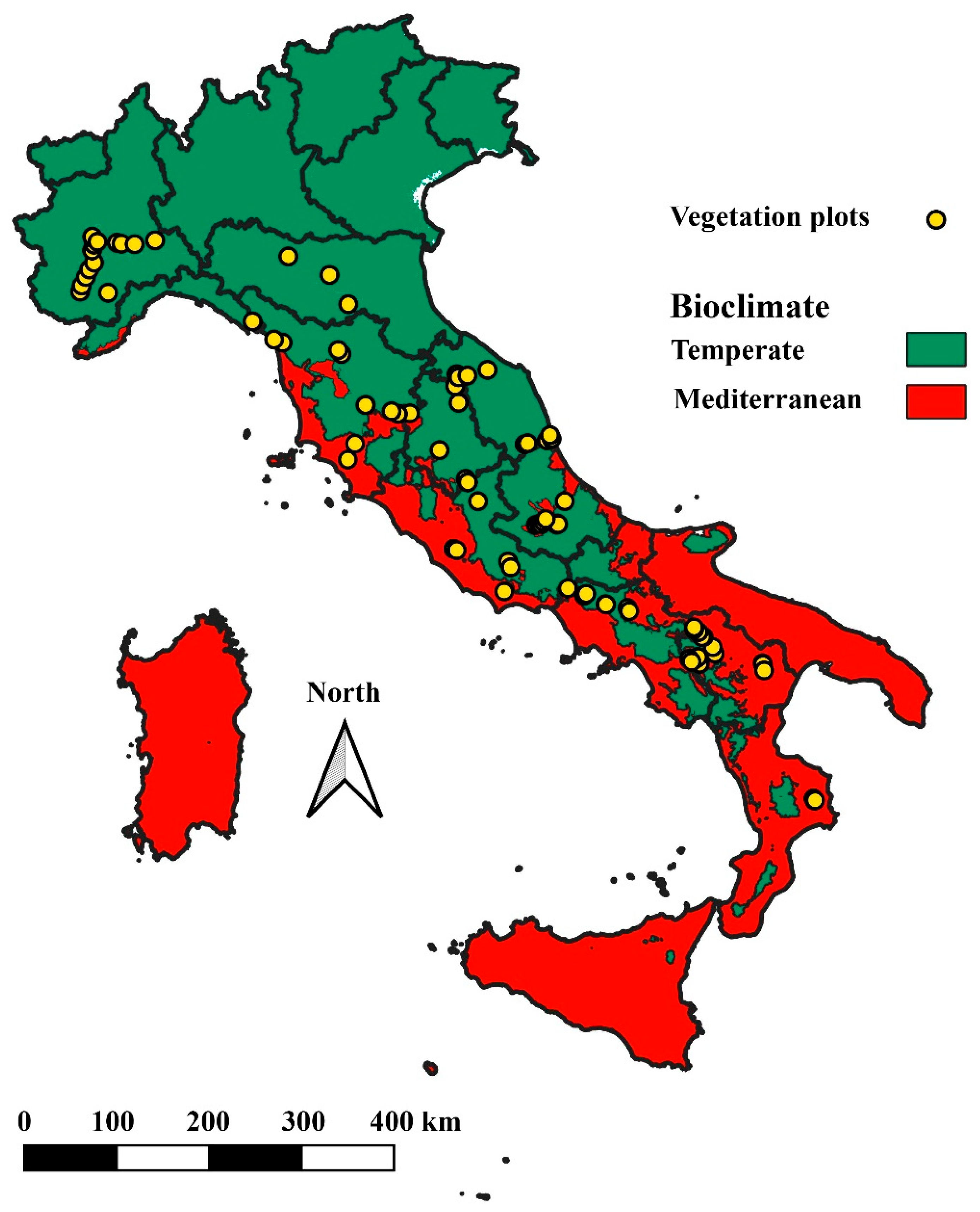
2.1. Study Area and Sampling Design
2.2. Dataset Preparation and Statistical Analyses
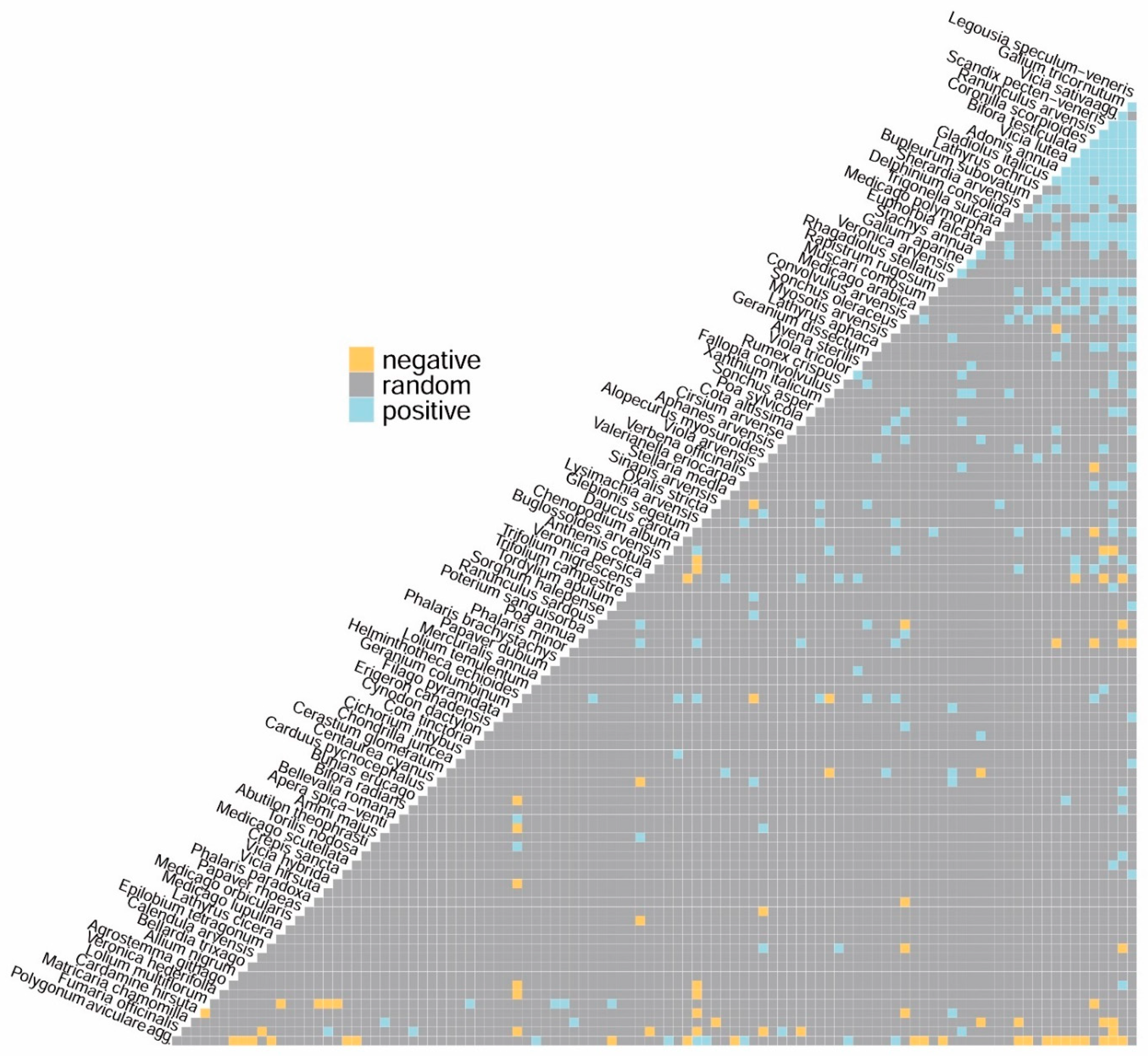
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Götzenberger, L.; de Bello, F.; Bråthen, K.A.; Davison, J.; Dubuis, A.; Guisan, A.; Lepš, J.; Lindborg, R.; Moora, M.; Pärtel, M.; et al. Ecological assembly rules in plant communities—Approaches, patterns and prospects. Biol. Rev. 2012, 87, 111–127. [Google Scholar] [CrossRef] [PubMed]
- Fridley, J.D.; Vandermast, D.B.; Kuppinger, D.M.; Manthey, M.; Peet, R.K. Co-occurrence based assessment of habitat generalists and specialists: A new approach for the measurement of niche width. J. Ecol. 2007, 95, 707–722. [Google Scholar] [CrossRef]
- Pannek, A.; Manthey, M.; Diekmann, M. Comparing resource-based and co-occurrence-based methods for estimating species niche breadth. J. Veg. Sci. 2016, 27, 596–605. [Google Scholar] [CrossRef]
- Bellanger, S.; Darmency, H.; Guillemin, J.P. Relationships between weed diversity and Centaurea cyanus. In Proceedings of the XIIIème Colloque International sur la Biologie des Mauvaises Herbes, Dijon, France, 8–10 September 2009; pp. 8–15. [Google Scholar]
- Bellanger, S.; Guillemin, J.P.; Bretagnolle, V.; Darmency, H. Centaurea cyanus as a biological indicator of segetal species richness in arable fields. Weed Res. 2012, 52, 551–563. [Google Scholar] [CrossRef]
- Petit, S.; Fried, G. Patterns of weed co-occurrence at the field and landscape level. J. Veg. Sci. 2012, 23, 1137–1147. [Google Scholar] [CrossRef]
- Holzner, W. Weed species and weed communities. Vegetatio 1978, 38, 13–20. [Google Scholar] [CrossRef]
- Marshall, E.J.P.; Brown, V.K.; Boatman, N.D.; Lutman, P.J.W.; Squire, G.R.; Ward, L.K. The role of weeds in supporting biological diversity within crop fields. Weed Res. 2003, 43, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Bretagnolle, V.; Gaba, S. Weeds for bees? A review. Agron Sustain Dev. 2015, 35, 891–909. [Google Scholar] [CrossRef] [Green Version]
- Blaix, C.; Moonen, A.C.; Dostatny, D.F.; Izquierdo, J.; Le Corff, J.; Morrison, J.; Von Redwitz, C.; Schumacher, M.; Westerman, P.R. Quantification of regulating ecosystem services provided by weeds in annual cropping systems using a systematic map approach. Weed Res. 2018, 58, 151–164. [Google Scholar] [CrossRef]
- Weisberger, D.; Nichols, V.; Liebman, M. Does diversifying crop rotations suppress weeds? A meta-analysis. PLoS ONE 2019, 14, e0219847. [Google Scholar] [CrossRef] [Green Version]
- Storkey, J.; Neve, P. What good is weed diversity? Weed Res. 2018, 58, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Šilc, U.; Vrbničanin, S.; Božić, D.; Čarni, A.; Dajić Stevanović, Z. Weed vegetation in the north-western Balkans: Diversity and species composition. Weed Res. 2009, 49, 602–612. [Google Scholar] [CrossRef]
- Mahaut, L.; Gaba, S.; Fried, G. A functional diversity approach of crop sequences reveals that weed diversity and abundance show different responses to environmental variability. J. Appl. Ecol. 2019, 56, 1400–1409. [Google Scholar] [CrossRef]
- Fanfarillo, E.; Petit, S.; Dessaint, F.; Rosati, L.; Abbate, G. Species composition, richness, and diversity of weed communities of winter arable land in relation to geo-environmental factors: A gradient analysis in mainland Italy. Botany 2020. [Google Scholar] [CrossRef]
- Lososová, Z.; Chytrý, M.; Cimalová, S.; Kropáč, Z.; Otýpková, Z.; Pyšek, P.; Tichý, L. Weed vegetation of arable land in central Europe: Gradients of diversity and species composition. J. Veg. Sci. 2004, 15, 415–422. [Google Scholar] [CrossRef]
- Fried, G.; Norton, R.L.; Reboud, X. Environmental and management factors determining weed species composition and diversity in France. Agric. Ecosyst. Environ. 2008, 128, 68–76. [Google Scholar] [CrossRef]
- Nowak, A.; Nowak, S.; Nobis, M.; Nobis, A. Crop type and altitude are the main drivers of species composition of arable weed vegetation in Tajikistan. Weed Res. 2015, 55, 525–536. [Google Scholar] [CrossRef]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; Gavilán García, R.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19 (Suppl. 1), 3–264. [Google Scholar] [CrossRef]
- Zohary, M. Plant Life of Palestine, Israel and Jordan; Ronald: New York, NY, USA, 1962. [Google Scholar]
- Scholz, H. Questions about indigenous plants and anecophytes. Taxon 2007, 56, 1255–1260. [Google Scholar] [CrossRef]
- Brullo, S.; Guarino, R. The Mediterranean weedy vegetation and its origin. Ann. di Bot. 2007, 7, 101–110. [Google Scholar]
- Abbate, G.; Cicinelli, E.; Iamonico, D.; Iberite, M. Floristic analysis of the weed communities in wheat and corn crops: A case study in western-central Italy. Ann. di Bot. 2013, 3, 97–105. [Google Scholar]
- Latini, M.; Fanfarillo, E.; De Luca, E.; Iberite, M.; Abbate, G. The weed vegetation of the bean “Fagiolo Cannellino di Atina” and the red pepper “Peperone di Pontecorvo” PDO crops (Latium, central Italy). Plant Sociol. 2020, 57, 1–10. [Google Scholar]
- Janssen, J.A.M.; Rodwell, J.S.; García Criado, M.; Gubbay, S.; Haynes, T.; Nieto, A.; Sanders, N.; Landucci, F.; Loidi, J.; Ssymank, A.; et al. European Red List of Habitats—Part 2: Terrestrial and Freshwater Habitats; Publication Office of the European Union: Luxembourg, 2016; pp. 1–38. [Google Scholar]
- Fanfarillo, E.; Latini, M.; Iberite, M.; Bonari, G.; Nicolella, G.; Rosati, L.; Salerno, G.; Abbate, G. The segetal flora of winter cereals and allied crops in Italy: Species inventory with chorological, structural and ecological features. Plant Biosyst. 2020. [Google Scholar] [CrossRef]
- Albrecht, H. Suitability of arable weeds as indicator organism to evaluate species conservation effects of management in agricultural ecosystems. Agr. Ecosyst. Environ. 2003, 98, 201–211. [Google Scholar] [CrossRef]
- Fried, G.; Chauvel, B.; Reboud, X. A functional analysis of large-scale temporal shifts from 1970 to 2000 in weed assemblages of sunflower crops in France. J. Veg. Sci. 2009, 20, 49–58. [Google Scholar] [CrossRef]
- Richner, N.; Holderegger, R.; Linder, H.P.; Walter, T. Reviewing change in the arable flora of Europe: A meta-analysis. Weed Res. 2015, 55, 1–13. [Google Scholar] [CrossRef]
- Fanfarillo, E.; Kasperski, A.; Giuliani, A.; Abbate, G. Shifts of arable plant communities after agricultural intensification: A floristic and ecological diachronic analysis in maize fields of Latium (central Italy). Bot. Lett. 2019, 166, 356–365. [Google Scholar] [CrossRef]
- Hilbig, W. Preservation of agrestal weeds. In Biology and Ecology of Weeds; Holzner, W., Numata, M., Eds.; Dr W. Junk Publishers: The Hague, The Netherlands, 1982; pp. 57–59. [Google Scholar]
- Aboucaya, A.; Jauzein, P.; Vinciguerra, L.; Virevaire, M. Plan National d’action pour la Conservation des Plantes Messicoles. Rapport Final rédigé à la Demande du Ministère de l’Aménagement du Territoire et de l’Environnement, Direction De La Nature Et Des Paysages; Conservatoire Botanique National Méditerranéen de Porquerolles: Hyères, France, 2000. [Google Scholar]
- Cambecèdes, J.; Largier, G.; Lombard, A. Plan National d’actions en Faveur des Plantes Messicoles; Conservatoire Botanique National des Pyrénées et de Midi-Pyrénées: Bagnères de Bigorre, France, 2012. [Google Scholar]
- Storkey, J.; Meyer, S.; Still, K.S.; Leuschner, C. The impact of agricultural intensification and land-use change on the European arable flora. Proc. R. Soc. B. 2012, 279, 1421–1429. [Google Scholar] [CrossRef]
- Nowak, A.; Nowak, S.; Nobis, M.; Nobis, A. A report on the conservation status of segetal weeds in Tajikistan. Weed Res. 2014, 54, 635–648. [Google Scholar] [CrossRef]
- Albrecht, H.; Cambecèdes, J.; Lang, M.; Wagner, M. Management options for the conservation of rare arable plants in Europe. Bot Lett. 2016, 163, 389–415. [Google Scholar] [CrossRef] [Green Version]
- Fanfarillo, E.; Scoppola, A.; Lososová, Z.; Abbate, G. Segetal plant communities of traditional agroecosystems: A phytosociological survey in central Italy. Phytocoenologia 2019, 49, 165–183. [Google Scholar] [CrossRef]
- Fanfarillo, E.; Latini, M.; Iberite, M.; Abbate, G. Segetal flora of Italy. Version 1.5. Laboratory of Systematic Botany and Floristics—Department of Environmental Biology, Sapienza University, 2020. Occurrence dataset. Available online: https://doi.org/10.15468/44yxfz (accessed on 30 April 2020).
- Pesaresi, S.; Biondi, E.; Casavecchia, S. Bioclimates of Italy. J. Maps 2017, 13, 955–960. [Google Scholar] [CrossRef]
- Costantini, E.A.C.; Barbetti, R.; Fantappiè, M.; L’Abate, G.; Lorenzetti, R.; Magini, S. Pedodiversity. In The Soils of Italy; Dazzi, C., Ed.; Springer: Berlin, Germany, 2013. [Google Scholar]
- Chytrý, M.; Otýpková, Z. Plot sizes used for phytosociological sampling of European vegetation. J. Veg. Sci. 2003, 14, 563–570. [Google Scholar] [CrossRef]
- Güler, B.; Jentsch, A.; Apostolova, I.; Bartha, S.; Bloor, J.M.G.; Campetella, G.; Canullo, R.; Házi, J.; Kreyling, J.; Poittier, J.; et al. How plot shape and spatial arrange-affect plant species richness counts: Implications for sampling design and rarefaction analyses. J. Veg. Sci. 2016, 27, 692–703. [Google Scholar] [CrossRef] [Green Version]
- Braun-Blanquet, J. Pflanzensoziologie; Springer: Vienna, Austria, 1964. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia; Edagricole-New Business Media: Bologna, Italy, 2017; Volume 2, pp. 1065–2260. [Google Scholar]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Galasso, G.; Bartolucci, F.; Peruzzi, L.; Ardenghi, N.M.G.; Albano, A.; Alessandrini, A.; Bacchetta, G.; Ballelli, S.; Bandini Mazzanti, M.; Barberis, G.; et al. An updated checklist of the vascular flora alien to Italy. Plant Biosyst. 2018, 152, 556–592. [Google Scholar] [CrossRef]
- Fried, G.; Petit, S.; Reboud, X. A specialist-generalist classification of the arable flora and its response to changes in agricultural practices. BMC Ecol. 2010, 10, 20. [Google Scholar] [CrossRef] [Green Version]
- Pignatti, S.; Menegoni, P.; Pietrosanti, S. Bioindicazione attraverso le piante vascolari. Valori di indicazione secondo Ellenberg (zeigerwerte) per le specie della Flora d’Italia. Braun-Blanquetia 2005, 39, 97. [Google Scholar]
- Griffith, M.D.; Veech, J.A.; Marsh, C.J. Cooccur: Probabilistic Species Co-Occurrence Analysis in R. J. Stat. Softw. 2016, 69. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 12 May 2020).
- Veech, J.A. A Probabilistic Model for Analysing Species Co-Occurrence: Probabilistic Model. Global Ecol. Biogeogr. 2013, 22, 252–260. [Google Scholar] [CrossRef]
- Isbell, F.; Reich, P.B.; Tilman, D.; Hobbie, S.E.; Polasky, S.; Binder, S. Nutrient enrichment, biodiversity loss, and consequent declines in ecosystem productivity. PNAS 2013, 110, 11911–11916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chytrý, M. Phytosociological data give biased estimates of species richness. J. Veg. Sci. 2001, 12, 439–444. [Google Scholar] [CrossRef]
- Perrino, E.V.; Calabrese, G. Endangered segetal species in southern Italy: Distribution, conservation status, trends, actions and ethnobotanical notes. Genet. Resour. Crop. Evol. 2018, 65, 2107–2134. [Google Scholar] [CrossRef]
- Condit, R.S.; Ashton, P.S.; Baker, P.J.; Bunyavejchewin, S.; Gunatilleke, S.; Gunatilleke, N.; Hubbell, S.P.; Foster, R.B.; Itoh, A.; LaFrankie, J.V.; et al. Spatial patterns in the distribution of tropical tree species. Science 2000, 288, 1414–1418. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Huang, Z.; Ye, W.; Cao, H.; Wei, S.; Wang, Z.; Lian, J.; Sun, I.-F.; Ma, K.; He, F. Spatial distributions of tree species in a subtropical forest of China. Oikos 2009, 118, 495–502. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Rare and Threatened Taxa | Positively Associated Taxa | Negatively Associated Taxa |
|---|---|---|
| Adonis annua | Aphanes arvensis; Bifora testiculata; Bupleurum subovatum; Coronilla scorpioides; Cota altissima; Delphinium consolida; Euphorbia falcata; Galium tricornutum; Geranium dissectum; Gladiolus italicus; Legousia speculum-veneris; Medicago polymorpha; Muscari comosum; Ranunculus arvensis; Scandix pecten-veneris; Vicia sativa agg.; Vicia lutea | Poa annua*; Polygonum aviculare agg.; Sonchus oleraceus* |
| Agrostemma githago | Galium tricornutum | Helminthotheca echioides; Lysimachia arvensis |
| Bifora radians | Galium tricornutum | |
| Centaurea cyanus | Veronica arvensis; Viola arvensis | Veronica persica |
| Filago pyramidata | Legousia speculum-veneris | |
| Galium tricornutum | Adonis annua; Agrostemma githago; Bifora radians; Bifora testiculata; Buglossoides arvensis; Bupleurum subovatum; Carduus pycnocephalus; Convolvulus arvensis; Coronilla scorpioides; Euphorbia falcata; Gladiolus italicus; Lathyrus aphaca; Lathyrus ochrus; Legousia speculum-veneris; Medicago arabica; Medicago polymorpha; Muscari comosum; Ranunculus arvensis; Rhagadiolus stellatus; Scandix pecten-veneris; Sonchus asper*; Torilis nodosa; Trigonella sulcata; Valerianella eriocarpa; Vicia sativa agg.; Vicia lutea | Epilobium tetragonum; Poa annua*; Polygonum aviculare agg.; Ranunculus sardous; Veronica persica |
| Lathyrus aphaca | Alopecurus myosuroides*; Galium tricornutum; Legousia speculum-veneris; Ranunculus arvensis; Scandix pecten-veneris | |
| Legousia speculum-veneris | Adonis annua; Anthemis cotula; Avena sterilis; Bifora testiculata; Bupleurum subovatum; Chondrilla juncea; Convolvulus arvensis; Coronilla scorpioides; Cota altissima; Crepis sancta; Delphinium consolida; Euphorbia falcata; Fallopia convolvulus; Filago pyramidata; Galium tricornutum; Gladiolus italicus; Glebionis segetum; Lathyrus aphaca; Lathyrus ochrus; Medicago polymorpha; Muscari comosum; Myosotis arvensis; Ranunculus arvensis; Rhagadiolus stellatus; Scandix pecten-veneris; Stachys annua; Tordylium apulum; Trigonella sulcata; Vicia lutea; Viola arvensis | Poa annua* |
| Lolium temulentum | Scandix pecten-veneris | |
| Ranunculus arvensis | Adonis annua; Anthemis cotula; Bifora testiculata; Bupleurum subovatum; Convolvulus arvensis; Coronilla scorpioides; Delphinium consolida; Euphorbia falcata; Fallopia convolvulus; Galium tricornutum; Helminthotheca echioides; Lathyrus aphaca; Lathyrus ochrus; Legousia speculum-veneris; Medicago polymorpha; Rhagadiolus stellatus; Scandix pecten-veneris; Stachys annua; Trigonella sulcata; Valerianella eriocarpa; Vicia sativa agg.; Vicia lutea | Glebionis segetum; Lolium multiflorum; Poa annua*; Viola arvensis |
| Scandix pecten-veneris | Adonis annua; Aphanes arvensis; Bifora testiculata; Buglossoides arvensis; Bupleurum subovatum; Coronilla scorpioides; Delphinium consolida; Euphorbia falcata; Galium tricornutum; Gladiolus italicus; Medicago arabica; Medicago polymorpha; Muscari comosum; Lathyrus aphaca; Lathyrus ochrus; Legousia speculum-veneris; Lolium temulentum; Ranunculus arvensis; Rhagadiolus stellatus; Sherardia arvensis; Trigonella sulcata; Vicia lutea; Vicia sativa agg. | Chenopodium album*; Matricaria chamomilla; Polygonum aviculare agg.; Veronica persica |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fanfarillo, E.; Latini, M.; Abbate, G. Patterns of Co-occurrence of Rare and Threatened Species in Winter Arable Plant Communities of Italy. Diversity 2020, 12, 195. https://doi.org/10.3390/d12050195
Fanfarillo E, Latini M, Abbate G. Patterns of Co-occurrence of Rare and Threatened Species in Winter Arable Plant Communities of Italy. Diversity. 2020; 12(5):195. https://doi.org/10.3390/d12050195
Chicago/Turabian StyleFanfarillo, Emanuele, Marta Latini, and Giovanna Abbate. 2020. "Patterns of Co-occurrence of Rare and Threatened Species in Winter Arable Plant Communities of Italy" Diversity 12, no. 5: 195. https://doi.org/10.3390/d12050195
APA StyleFanfarillo, E., Latini, M., & Abbate, G. (2020). Patterns of Co-occurrence of Rare and Threatened Species in Winter Arable Plant Communities of Italy. Diversity, 12(5), 195. https://doi.org/10.3390/d12050195