Acacia longifolia: A Host of Many Guests Even after Fire




Abstract
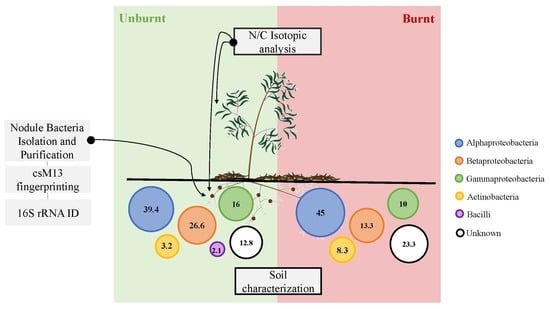
:
1. Introduction
2. Materials and Methods
2.1. Study Site Description and Nodules Collection
2.2. Soil Characteristics
2.3. Isotopic Analysis
2.4. Isolation and Phenotypic Characterization of Nodule Bacteria
2.5. DNA Fingerprinting of Bacterial Isolates
2.6. Identification of Bacterial Isolates by 16S rRNA Gene Sequencing
2.7. Statistical Analysis
3. Results
3.1. Nodulation of Young Plants
3.2. Isotopic Analysis
PCA Analysis
3.3. Bacterial Fingerprinting and Identification
4. Discussion
4.1. Nodulation: Does Fire Play a Role?
4.2. Nodule Bacteriome: Who Is Taking Part?
4.3. Bacteriome: What Could be Occurring?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Davis, M.A.; Chew, M.K.; Hobbs, R.J.; Lugo, A.E.; Ewel, J.J.; Vermeij, G.J.; Brown, J.H.; Rosenzweig, M.L.; Gardener, M.R.; Carroll, S.P.; et al. Don’t judge species on their origins. Nature 2011, 474, 153–154. [Google Scholar] [CrossRef] [PubMed]
- Hierro, J.L.; Maron, J.L.; Callaway, R.M. A biogeographical approach to plant invasions: The importance of studying exotics in their introduced and native range. J. Ecol. 2005, 93, 5–15. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P. Plant Invasions. Encycl. Biodivers. 2013, 6, 90–102. [Google Scholar] [CrossRef]
- Foyer, C.H.; Lam, H.M.; Nguyen, H.T.; Siddique, K.H.; Varshney, R.K.; Colmer, T.D.; Cowling, W.; Bramley, H.; Mori, T.A.; Hodgson, J.M.; et al. Neglecting legumes has com- promised human health and sustainable food production. Nat. Plants 2016, 2, 16112. [Google Scholar] [CrossRef]
- Maslin, B.R.; Miller, J.; Seigler, D.S. Overview of the generic status of Acacia (Leguminosae: Mimosoideae). Aust. Syst. Bot. 2003, 16, 1–18. [Google Scholar] [CrossRef]
- Whibley, D.J.E. Acacias of South Australia; Wollman, D.J., Ed.; Government Printer: Adelaide, Australia, 1980. [Google Scholar]
- Alonso, P.S.; Rodríguez, J.; González, L.; Lorenzo, P. Here to stay. Recent advances and perspectives about Acacia invasion in Mediterranean areas. Ann. For. Sci. 2017, 74, 55. [Google Scholar] [CrossRef] [Green Version]
- Peperkorn, R.; Werner, C.; Beyschlag, W. Phenotypic plasticity of an invasive acacia versus two native Mediterranean species. Funct. Plant Biol. 2005, 32, 933–944. [Google Scholar] [CrossRef]
- Morais, M.C.; Marchante, H.; Marchante, H. Big troubles are already here: Risk assessment protocol shows high risk of many alien plants present in Portugal. J. Nat. Conserv. 2017, 35, 1–12. [Google Scholar] [CrossRef]
- Stock, W.D.; Wienand, K.T.; Baker, A.C. Impacts of invading N2-fixing Acacia species on patterns of nutrient cycling in two Cape ecosystems: Evidence from soil incubation studies and 15N natural abundance values. Oecologia 1995, 101, 375–382. [Google Scholar] [CrossRef]
- Marchante, H.; Marchante, E.; Buscardo, E.; Maia, J.; Freitas, H. Recovery Potential of Dune Ecosystems Invaded by an Exotic Acacia Species (Acacia longifolia)1. Weed Technol. 2004, 18, 1427–1433. [Google Scholar]
- Yelenik, S.G.; Stock, W.D.; Richardson, D.M. Functional Group Identity Does not Predict Invader Impacts: Differential Effects of Nitrogen-fixing Exotic Plants on Ecosystem Function. Biol. Invasions 2006, 9, 117–125. [Google Scholar] [CrossRef]
- Carvalho, L.; Antunes, P.M.; Martins-Loução, M.A.; Klironomos, J.N. Disturbance influences the outcome of plant-soil biota interactions in the invasive Acacia longifolia and in native species. Oikos 2010, 119, 1172–1180. [Google Scholar] [CrossRef]
- Ulm, F.; Hellmann, C.; Cruz, C.; Máguas, C. N/P imbalance as a key driver for the invasion of oligothrophic dune systems by a woody legume. Oikos 2016, 126, 231–240. [Google Scholar] [CrossRef]
- Meira-Neto, J.A.A.; Da Silva, M.C.N.A.; Tolentino, G.S.; Gastauer, M.; Buttschardt, T.; Ulm, F.; Máguas, C. Early Acacia invasion in a sandy ecosystem enables shading mediated by soil, leaf nitrogen and facilitation. Biol. Invasions 2017, 20, 1567–1575. [Google Scholar] [CrossRef]
- Andrews, M.; Andrews, M. Specificity in Legume-Rhizobia Symbioses. Int. J. Mol. Sci. 2017, 18, 705. [Google Scholar] [CrossRef] [Green Version]
- Kamutando, C.N.; Vikram, S.; Kamgan-Nkuekam, G.; Makhalanyane, T.P.; Greve, M.; Le Roux, J.J.; Richardson, D.M.; Cowan, D.A.; Valverde, A. The Functional Potential of the Rhizospheric Microbiome of an Invasive Tree Species, Acacia dealbata. Microb. Ecol. 2018, 77, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Echeverría, S. Rhizobial hitchhikers from Down Under: Invasional meltdown in a plant-bacteria mutualism? J. Biogeogr. 2010, 37, 1611–1622. [Google Scholar] [CrossRef]
- Lafay, B.; Burdon, J.J. Small-Subunit rRNA Genotyping of Rhizobia Nodulating Australian Acacia spp. Appl. Environ. Microbiol. 2001, 67, 396–402. [Google Scholar] [CrossRef] [Green Version]
- Marsudi, N. Identification and characterization of fast- and slow-growing root nodule bacteria from South-Western Australian soils able to nodulate Acacia saligna. Soil Biol. Biochem. 1999, 31, 1229–1238. [Google Scholar] [CrossRef]
- Nick, G.; De Lajudie, P.; Eardly, B.D.; Suomalainen, S.; Paulin, L.; Zhang, X.; Gillis, M.; Lindström, K. Sinorhizobium arboris sp. nov. and Sinorhizobium kostiense sp. nov., isolated from leguminous trees in Sudan and Kenya. Int. J. Syst. Evol. Microbiol. 1999, 49, 1359–1368. [Google Scholar] [CrossRef]
- Nick, G.; Jussila, M.; Hoste, B.; Niemi, M.; Kaijalainen, S.; Lajudie, P.; de Gillis, M.; Bruijn, F.J.; de Lindström, K. Rhizobia isolated from root nodules of tropical leguminous trees characterized using DNA-DNA dot-blot hybridization and rep-PCR genomic fingerprinting. Syst. Appl. Microbiol. 1999, 22, 287–299. [Google Scholar] [CrossRef]
- Rodríguez-Echeverría, S.; Crisostomo, J.; Freitas, H. Genetic Diversity of Rhizobia Associated with Acacia longifolia in Two Stages of Invasion of Coastal Sand Dunes. Appl. Environ. Microbiol. 2007, 73, 5066–5070. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Echeverría, S.; Le Roux, J.J.; Crisostomo, J.; Ndlovu, J. Jack-of-all-trades and master of many? How does associated rhizobial diversity influence the colonization success of Australian Acacia species? Divers. Distrib. 2011, 17, 946–957. [Google Scholar] [CrossRef]
- Weiss, P.W. Seed characteristics and regeneration of some species in invaded coastal communities. Austral Ecol. 1984, 9, 99–106. [Google Scholar] [CrossRef]
- Martínez-Hidalgo, P.; Hirsch, A.M. The Nodule Microbiome: N2-Fixing Rhizobia Do Not Live Alone. Phytobiomes J. 2017, 1, 70–82. [Google Scholar] [CrossRef] [Green Version]
- Boletim climatológico 2018. Available online: http://www.ipma.pt/pt/publicacoes/boletins (accessed on 30 April 2020).
- Sankhla, I.S.; Tak, N.; Meghwal, R.R.; Choudhary, S.; Tak, A.; Rathi, S.; Sprent, J.I.; James, E.K.; Gehlot, H. Molecular characterization of nitrogen fixing microsymbionts from root nodules of Vachellia (Acacia) jacquemontii, a native legume from the Thar Desert of India. Plant Soil 2016, 410, 21–40. [Google Scholar] [CrossRef]
- Callow, J.A.; Vincent, J.M. A Manual for the Practical Study of Root-Nodule Bacteria. J. Appl. Ecol. 1971, 8, 977. [Google Scholar] [CrossRef]
- Cappuccino, J.; Sherman, N. Experiment 2, Techniques for Isolation of Pure Cultures. In Microbiology, A Laboratory Manual, 5th ed.; Benjamin-Cummings Pub Co: San Francisco, CA, 1998; p. 16. [Google Scholar]
- Crisostomo, J.; Rodríguez-Echeverría, S.; Freitas, H. Co-introduction of exotic rhizobia to the rhizosphere of the invasive legume Acacia saligna, an intercontinental study. Appl. Soil Ecol. 2013, 64, 118–126. [Google Scholar] [CrossRef]
- Pitcher, D.; Saunders, N.; Owen, R. Rapid extraction of bacterial genomic DNA with guanidium thiocyanate. Lett. Appl. Microbiol. 1989, 8, 151–156. [Google Scholar] [CrossRef]
- Vassart, G.; Georges, M.; Monsieur, R.; Brocas, H.; Lequarre, A.; Christophe, D. A sequence in M13 phage detects hypervariable minisatellites in human and animal DNA. Science 1987, 235, 683–684. [Google Scholar] [CrossRef] [Green Version]
- Krebs, C. Ecological Methodology; Harper Collins: New York, NY, USA, 1989. [Google Scholar]
- Marchesi, J.R.; Sato, T.; Weightman, A.J.; Martin, T.A.; Fry, J.C.; Hiom, S.J.; Wade, W.G. Design and Evaluation of Useful Bacterium-Specific PCR Primers That Amplify Genes Coding for Bacterial 16S rRNA. Appl. Environ. Microbiol. 1998, 64, 795–799. [Google Scholar] [CrossRef] [Green Version]
- Geneious v5.3. 2010. Available online: http://www.geneious.com (accessed on 3 April 2019).
- Covington, W.; Sackett, S. Soil mineral nitrogen changes following prescribed burning in ponderosa pine. For. Ecol. Manag. 1992, 54, 175–191. [Google Scholar] [CrossRef]
- Lim, C.W.; Lee, Y.W.; Lee, S.C.; Hwang, C.H. Nitrate inhibits soybean nodulation by regulating expression of CLE genes. Plant Sci. 2014, 229, 1–9. [Google Scholar] [CrossRef]
- Ferguson, B.; Foo, E.; Ross, J.J.; Reid, J. Relationship between gibberellin, ethylene and nodulation in Pisum sativum. New Phytol. 2010, 189, 829–842. [Google Scholar] [CrossRef]
- Streeter, J.; Wong, P.P. Inhibition of legume nodule formation and N2fixation by nitrate. Crit. Rev. Plant Sci. 1988, 7, 1–23. [Google Scholar] [CrossRef]
- Gordon, A.; Skot, L.; James, C.; Minchin, F.R. Short-term metabolic responses of soybean root nodules to nitrate. J. Exp. Bot. 2002, 53, 423–428. [Google Scholar] [CrossRef]
- Harper, J.E. Soil and symbiotic nitrogen requirements for optimum production of soybean. Crop Sci. 1974, 14, 255–260. [Google Scholar] [CrossRef]
- Shearer, G.; Bryan, B.A.; Kohl, D.H. Increase of Natural 15N Enrichment of Soybean Nodules with Mean Nodule Mass. Plant Physiol. 1984, 76, 743–746. [Google Scholar] [CrossRef] [Green Version]
- Michelsen, A.; Quarmby, C.; Sleep, D.; Jonasson, S. Vascular plant 15 N natural abundance in heath and forest tundra ecosystems is closely correlated with presence and type of mycorrhizal fungi in roots. Oecologia 1998, 115, 406–418. [Google Scholar] [CrossRef]
- Mortimer, P.E.; Le Roux, M.R.; Perez-Fernandez, M.; Benedito, V.A.; Kleinert, A.; Xu, J.-C.; Valentine, A. The dual symbiosis between arbuscular mycorrhiza and nitrogen fixing bacteria benefits the growth and nutrition of the woody invasive legume Acacia cyclops under nutrient limiting conditions. Plant Soil 2012, 366, 229–241. [Google Scholar] [CrossRef]
- Israel, D.W. Investigation of the Role of Phosphorus in Symbiotic Dinitrogen Fixation. Plant Physiol. 1987, 84, 835–840. [Google Scholar] [CrossRef] [Green Version]
- Jensen, H.H.; Schjoerring, J.K.; Soussana, J.-F. The Influence of Phosphorus Deficiency on Growth and Nitrogen Fixation of White Clover Plants. Ann. Bot. 2002, 90, 745–753. [Google Scholar] [CrossRef] [Green Version]
- Taylor, B.; Simms, E.L.; La Pierre, K.J. More Than a Functional Group: Diversity within the Legume–Rhizobia Mutualism and Its Relationship with Ecosystem Function. Diversity 2020, 12, 50. [Google Scholar] [CrossRef] [Green Version]
- Barnet, Y.; Catt, P.; Hearne, D. Biological Nitrogen Fixation and Root-Nodule Bacteria (Rhizobium Sp. and Bradyrhizobium Sp.) In Two Rehabilitating Sand Dune Areas Planted With Acacia Spp. Aust. J. Bot. 1985, 33, 595–610. [Google Scholar] [CrossRef]
- Barnet, Y.; Catt, P. Distribution and characteristics of root-nodule bacteria isolated from Australian Acacia spp. Plant Soil 1991, 135, 109–120. [Google Scholar] [CrossRef]
- Lafay, B.; Burdon, J.J. Molecular Diversity of Rhizobia Occurring on Native Shrubby Legumes in Southeastern Australia. Appl. Environ. Microbiol. 1998, 64, 3989–3997. [Google Scholar] [CrossRef] [Green Version]
- Leary, J.; Singleton, P.; Scowcroft, P.; Borthakur, D. Symbiotic diversity in the cosmopolitan genus Acacia. Symbiosis 2006, 41, 107–117. [Google Scholar]
- Birnbaum, C.; Barrett, L.G.; Thrall, P.H.; Leishman, M.R. Mutualisms are not constraining cross?continental invasion success of Acacia species within Australia. Divers. Distrib. 2012, 18, 962–976. [Google Scholar] [CrossRef]
- Velázquez, E.; Martínez-Hidalgo, P.; Carro, L.; Alonso, P.; Peix, A.; Trujillo, M.E.; Martínez-Molina, E. Chapter 10, Nodular endophytes: An untapped diversity. In Beneficial Plant Microbial Interactions: Ecology and Applications, 1st ed.; Bélen Rodelas Gonzalez and Jésus Gonzalez-Lopez: Boca Ratón, FL, USA, 2014; pp. 215–236. [Google Scholar]
- Dourado, A.C.; Alves, P.I.L.; Tenreiro, T.; Ferreira, E.M.; Tenreiro, R.; Fareleira, P.; Crespo, M.T.B. Identification of Sinorhizobium (Ensifer) medicae based on a specific genomic sequence unveiled by M13-PCR fingerprinting. Int. Microbiol. 2009, 12, 215–225. [Google Scholar]
- Diouf, D.; Samba-Mbaye, R.; Lesueur, D.; Ba, A.T.; Dreyfus, B.; De Lajudie, P.; Neyra, M. Genetic Diversity of Acacia seyal Del. Rhizobial Populations Indigenous to Senegalese Soils in Relation to Salinity and pH of the Sampling Sites. Microb. Ecol. 2007, 54, 553–566. [Google Scholar] [CrossRef]
- Shiraishi, A.; Matsushita, N.; Hougetsu, T. Nodulation in black locust by the γ-proteobacteria Pseudomonas sp. and the β-proteobacteria Burkholderia sp. Systematic Appied. Microbiology 2010, 33, 269–274. [Google Scholar]
- Saïdi, S.; Chebil, S.; Gtari, M.; Mhamdi, R. Characterization of root-nodule bacteria isolated from Vicia faba and selection of plant growth promoting isolates. World J. Microbiol. Biotechnol. 2013, 29, 1099–1106. [Google Scholar] [CrossRef]
- Mårtensson, A.M.; Brutti, L.; Ljunggren, H. Competition between strains ofBradyrhizobium japonicum for nodulation of soybeans at different nitrogen fertilizer levels. Plant Soil 1989, 117, 219–225. [Google Scholar] [CrossRef]
- Kiers, E.T.; Rousseau, R.A.; West, S.A.; Denison, R.F. Host sanctions and the legume–rhizobium mutualism. Nature 2003, 425, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, B.; Mens, C.; Hastwell, A.; Zhang, M.; Su, H.; Jones, C.M.; Chu, X.; Gresshoff, P.M. Legume nodulation: The host controls the party. Plant Cell Environ. 2018, 42, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwivedi, S.L.; Sahrawat, K.L.; Upadhyaya, H.D.; Mengoni, A.; Galardini, M.; Bazzicalupo, M.; Biondi, E.G.; Hungria, M.; Kaschuk, G.; Blair, M.W.; et al. Advances in Host Plant and Rhizobium Genomics to Enhance Symbiotic Nitrogen Fixation in Grain Legumes. Adv. Agron. 2015, 129, 1–116. [Google Scholar] [CrossRef]
- Fox, J. Interpreting the “selection effect” of biodiversity on ecosystem function. Ecol. Lett. 2005, 8, 846–856. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P.; Rejmánek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Franche, C.; Lindström, K.; Elmerich, C. Nitrogen-fixing bacteria associated with leguminous and non-leguminous plants. Plant Soil 2008, 321, 35–59. [Google Scholar] [CrossRef]
- Ding, H.; Hynes, M.F. Plasmid transfer systems in the rhizobia. Can. J. Microbiol. 2009, 55, 917–927. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Texture | Water pH | OM (%) | Egner-Riehm Extraction | Mineral N – N mg·kg−1 | |||
|---|---|---|---|---|---|---|---|
| P2O5 mg·kg−1 | Total N g·kg−1 | N-NO3− | N-NH4+ | ||||
| UBZ | Coarse | 5.5 | 1.02 | 3 | 0.054* | 2.8 | 2.8 |
| BZ | Coarse | 5.4 | 2.08 | 16 | 0.124 | 4.3 | 6.3 |
| Leaves | Nodules | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| δ15N | δ13C | %N | %C | C/N | δ15N | δ13C | %N | %C | C/N | |
| UBZ | −1.0 | −30.2 | 2.4 | 44.3 | 19.1 | 7.9 | −29.5 | 5.3 | 36.5 | 7.0 |
| BZ | 0.8 | −29.5 | 3.3 | 43.5 | 15.2 | 7.4 | −28.8 | 6.3 | 42.3 | 7.0 |
| Genera/Zones | UBZ 1 | UBZ 2 | UBZ 3 | BZ 1 | BZ 2 | BZ 3 |
|---|---|---|---|---|---|---|
| Althererythrobacter sp. | 0 | 0 | 1 | 0 | 0 | 0 |
| Bradyrhizobium sp. | 9 | 12 | 13 | 6 | 16 | 5 |
| Paracoccus sp. | 0 | 1 | 0 | 0 | 0 | 0 |
| Rhizobium sp. | 0 | 1 | 0 | 0 | 0 | 0 |
| Caballeronia sp. | 2 | 0 | 0 | 1 | 0 | 0 |
| Duganella sp. | 8 | 0 | 0 | 0 | 0 | 0 |
| Paraburkholderia sp. | 4 | 6 | 5 | 4 | 2 | 1 |
| Moraxella sp. | 2 | 0 | 0 | 1 | 1 | 2 |
| Pseudomonas sp. | 12 | 0 | 1 | 1 | 4 | 1 |
| Micrococcus sp. | 2 | 0 | 0 | 1 | 1 | 2 |
| Nocardioides sp. | 1 | 0 | 0 | 1 | 0 | 0 |
| Paenibacillus sp. | 0 | 0 | 2 | 0 | 0 | 0 |
| Unknown | 1 | 10 | 1 | 9 | 3 | 2 |
| Total | 41 | 30 | 23 | 23 | 26 | 10 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jesus, J.G.d.; Tenreiro, R.; Máguas, C.; Trindade, H. Acacia longifolia: A Host of Many Guests Even after Fire. Diversity 2020, 12, 250. https://doi.org/10.3390/d12060250
Jesus JGd, Tenreiro R, Máguas C, Trindade H. Acacia longifolia: A Host of Many Guests Even after Fire. Diversity. 2020; 12(6):250. https://doi.org/10.3390/d12060250
Chicago/Turabian StyleJesus, Joana Guedes de, Rogério Tenreiro, Cristina Máguas, and Helena Trindade. 2020. "Acacia longifolia: A Host of Many Guests Even after Fire" Diversity 12, no. 6: 250. https://doi.org/10.3390/d12060250
APA StyleJesus, J. G. d., Tenreiro, R., Máguas, C., & Trindade, H. (2020). Acacia longifolia: A Host of Many Guests Even after Fire. Diversity, 12(6), 250. https://doi.org/10.3390/d12060250