Current and Future Distribution of Five Timber Forest Species in Amazonas, Northeast Peru: Contributions towards a Restoration Strategy

 ,
,  ,
,  ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
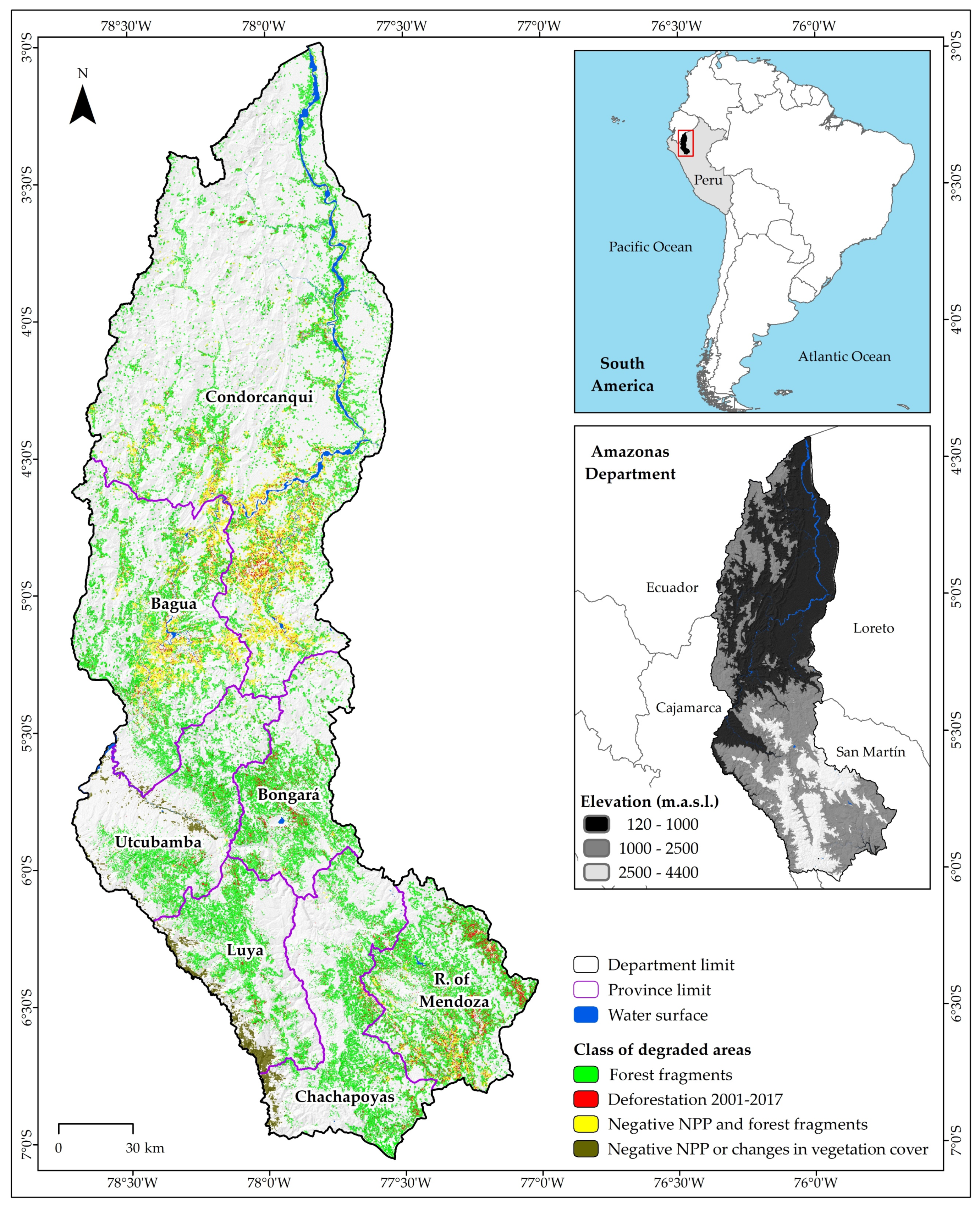
2.1. Study Area
2.2. Observed Geographical Records of Forest Species
2.3. Environmental Variables
2.4. Environmental Variable Selection
2.5. Species Distribution Modelling
2.6. Identifying Areas with Potential for Forest Restoration
3. Results
3.1. Performance of the Species Distribution Models
3.2. Environmental Variables Contributions
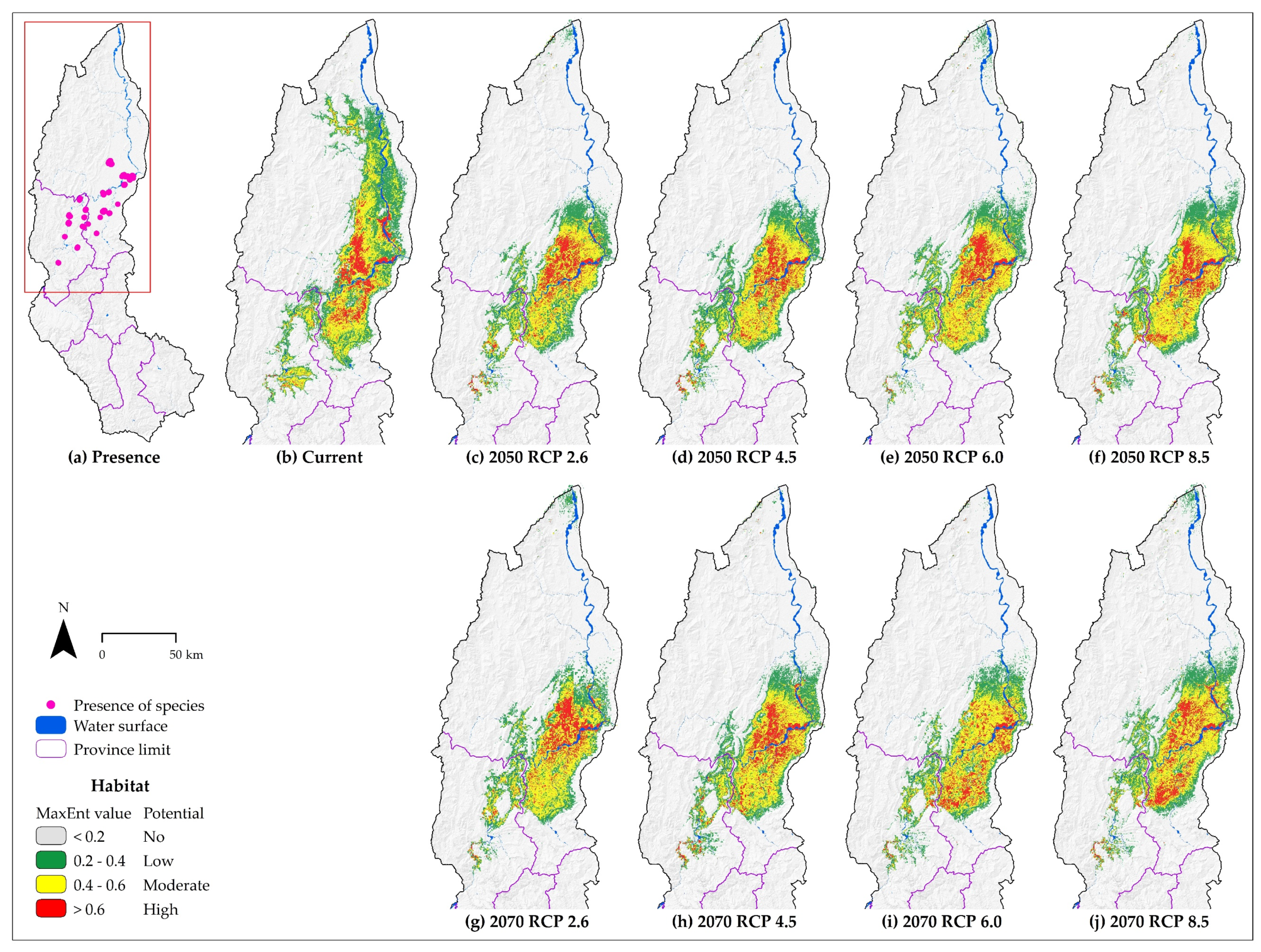
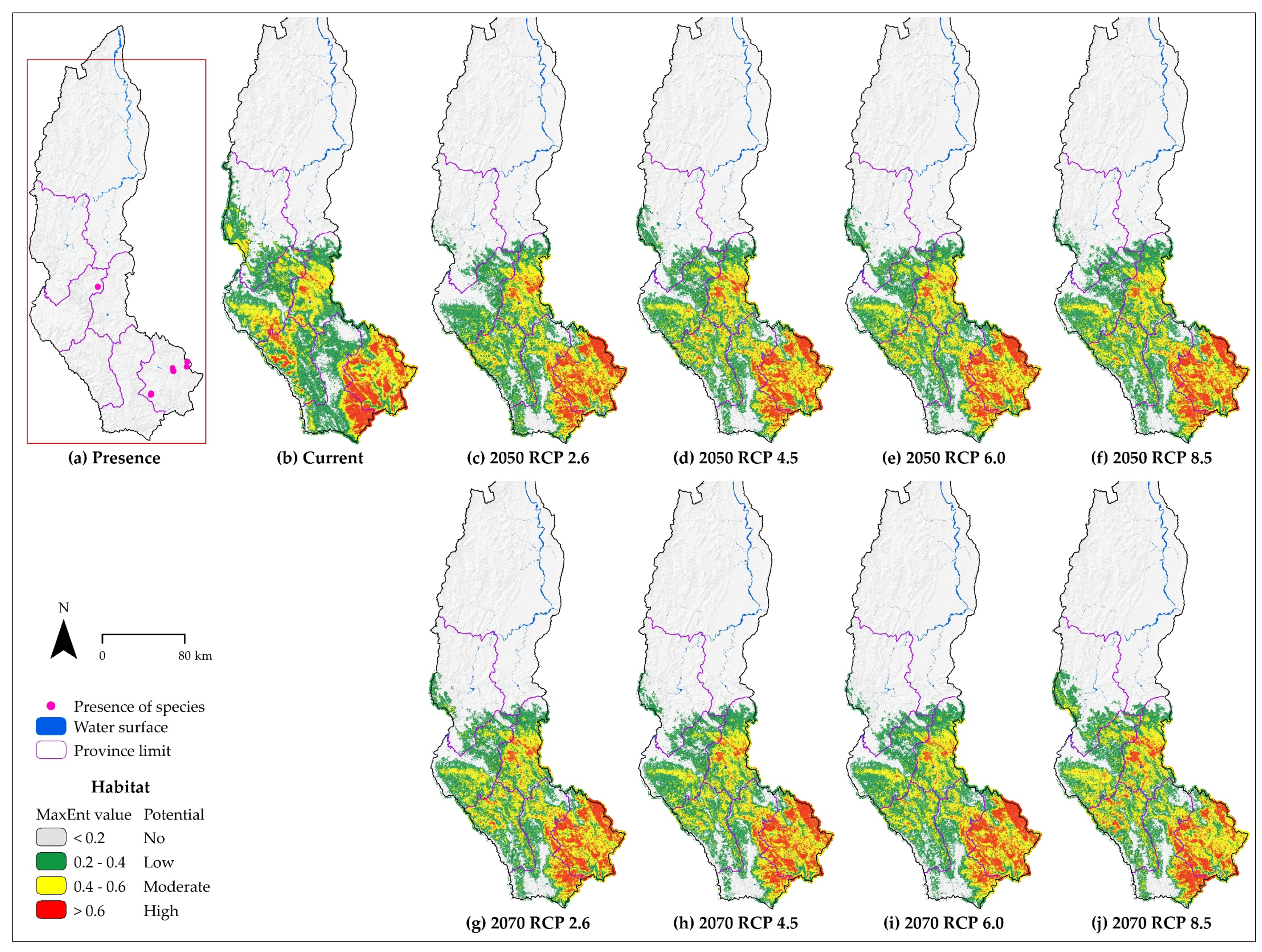
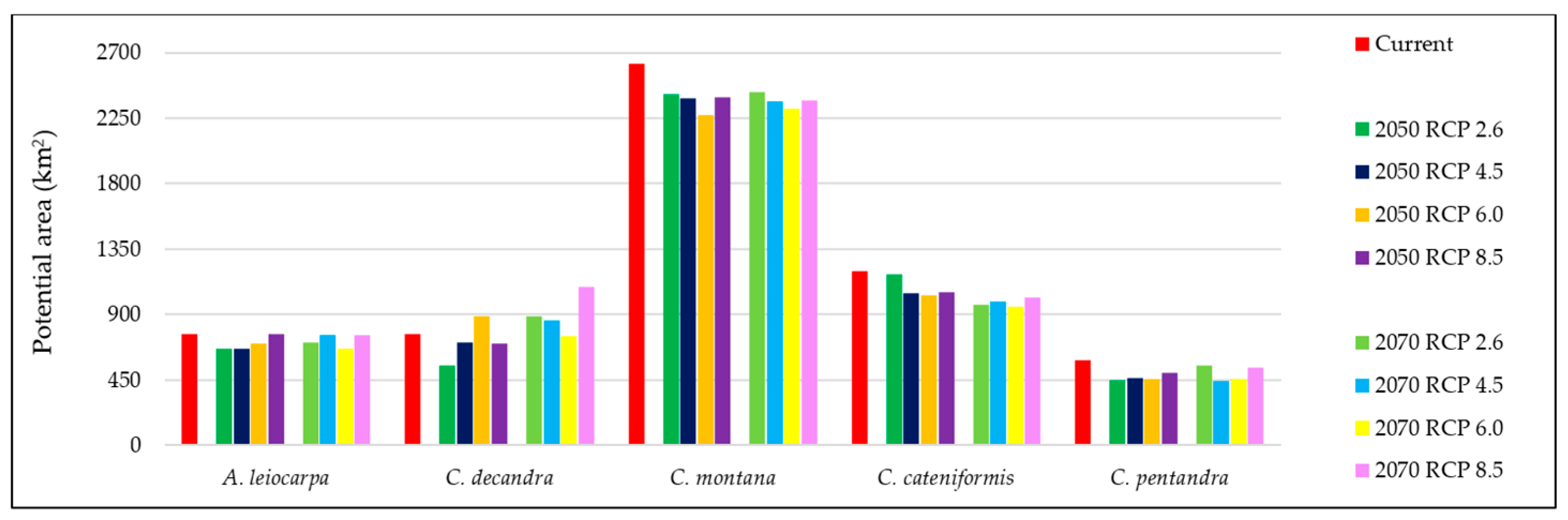
3.3. Species Distribution of Each Species
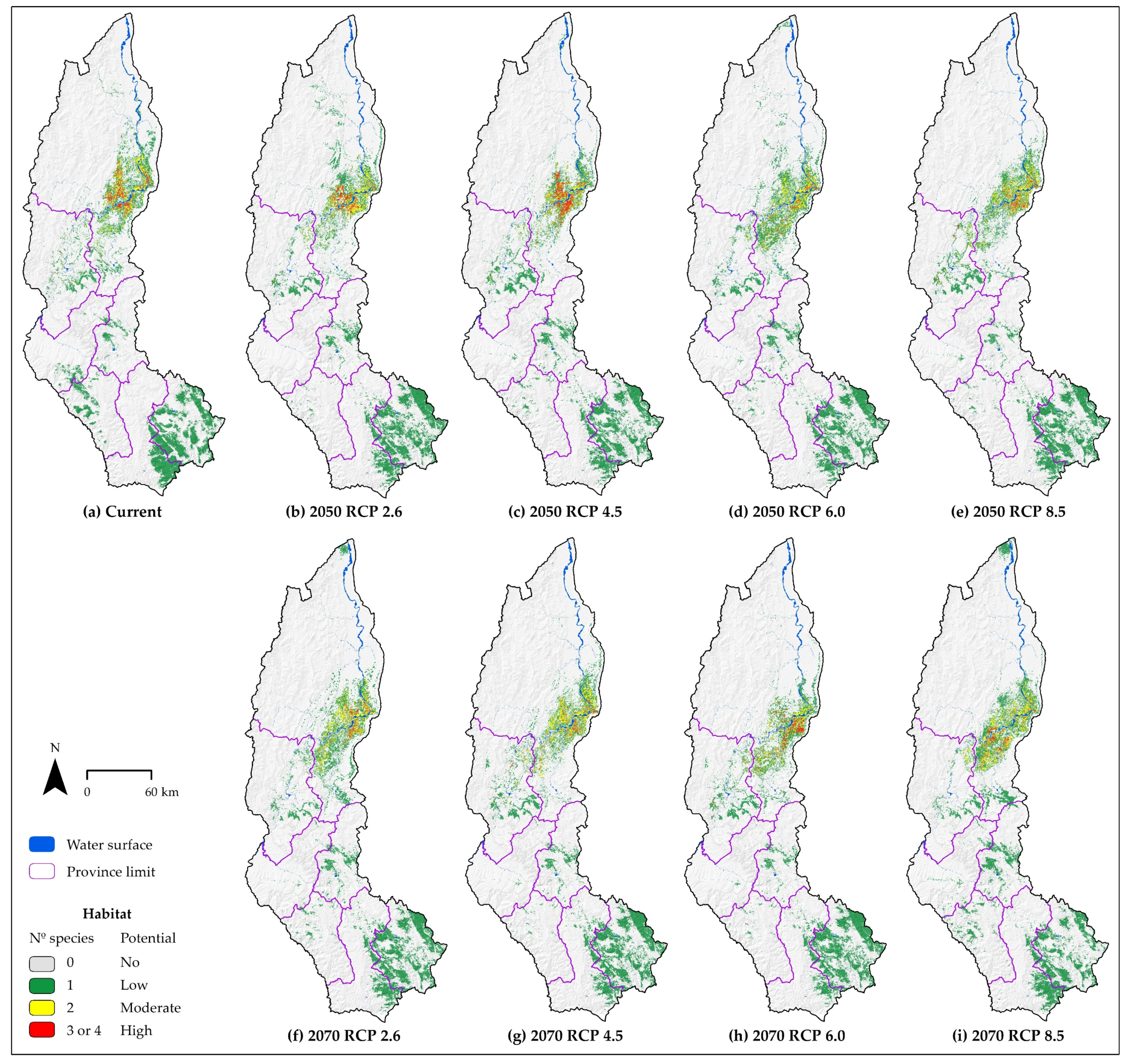
3.4. Combined Species Distribution of Multiple Species
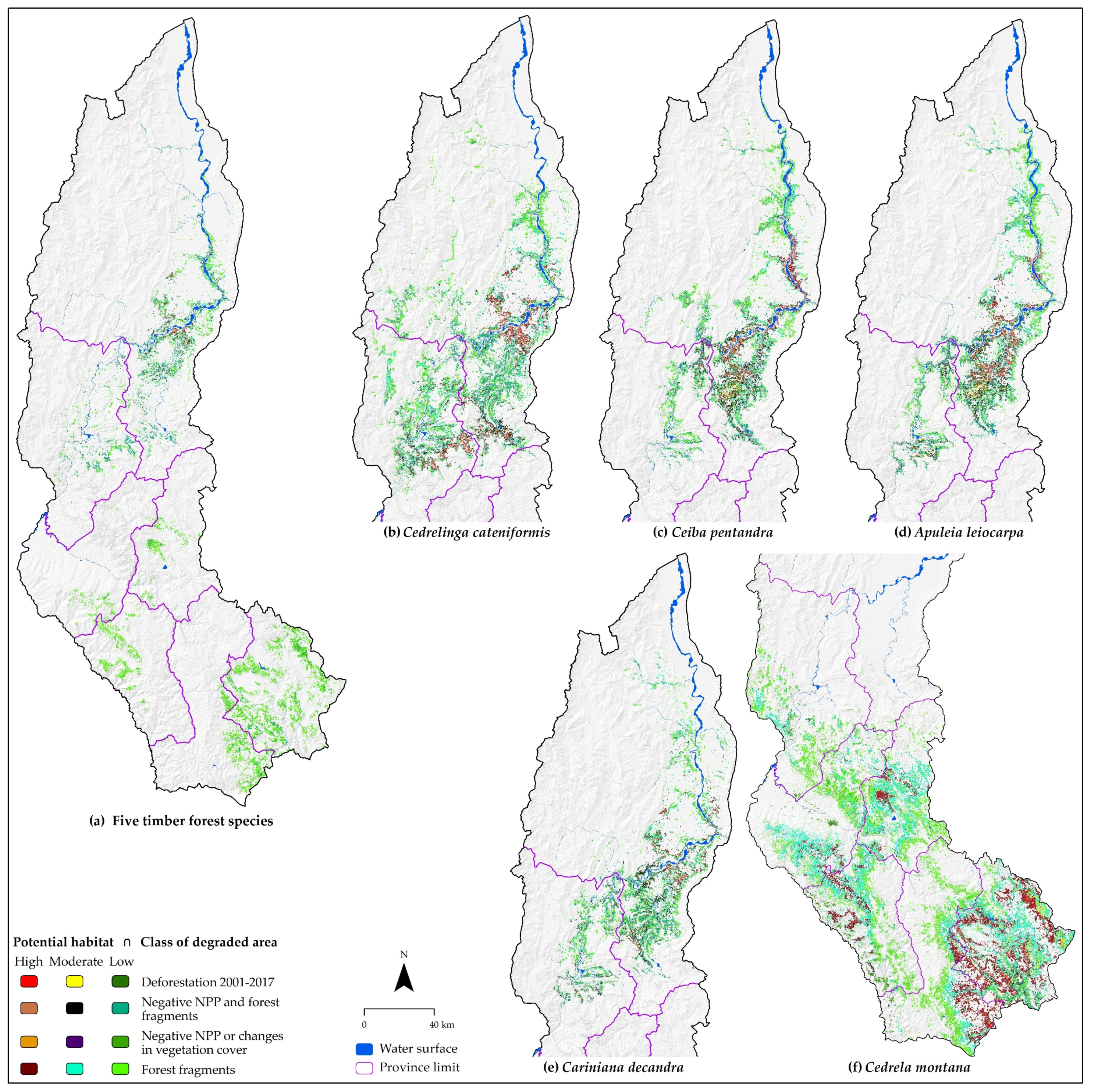
3.5. Degraded Lands with Restoration Potential
4. Discussion
4.1. About the Effects of Climate Change
4.2. About Forest Restoration in Peru
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sabogal, C.; Besacier, C.; McGuire, D. Forest and landscape restoration: Concepts, approaches and challenges for implementation. Unasylva 2015, 66, 3–10. [Google Scholar]
- FAO. Evaluación de los recursos forestales mundiales 2015. In ¿Cómo Están Cambiando Los Bosques Del Mundo? 2nd ed.; FAO: Roma, Italia, 2016; ISBN 9789253092833. [Google Scholar]
- MINAM. GEOBOSQUES: Bosque y Pérdida de Bosque. Available online: http://geobosques.minam.gob.pe/geobosque/view/perdida.php (accessed on 15 December 2019).
- De Sy, V.; Herold, M.; Achard, F.; Beuchle, R.; Clevers, J.G.P.W.; Lindquist, E.; Verchot, L. Land use patterns and related carbon losses following deforestation in South America. Environ. Res. Lett. 2015, 10, 124004. [Google Scholar] [CrossRef]
- Guariguata, R.M.; Arce, J.; Ammour, T.; Capella, J.L. Las Plantaciones Forestales en Perú: Reflexiones, Estatus Actual y Perspectivas a Futuro; CIFOR: Bogor, Indonesia, 2017; ISBN 9786023870530. [Google Scholar]
- Dourojeanni, R.M.J. Aprovechamiento del barbecho forestal en áreas de agricultura migratoria en la Amazonía peruana. Revi. For. Perú 1987, 14, 1–33. [Google Scholar] [CrossRef]
- MINAM. Estudio para la Identificación de Áreas Degradadas y Propuesta de Monitoreo; MINAM: Lima, Perú, 2017.
- Román, F.; Mamani, A.; Cruz, A.; Sandoval, C.; Cuesta, F. Orientaciones Para la restauracion de ecosistemas forestales y otros ecosistemas de vegetación Silvestre; SERFOR: Lima, Perú, 2018. [Google Scholar]
- Laestadius, L.; Maginnis, S.; Minnemeyer, S.; Potapov, P.; Saint-Laurent, C.; Sizer, N. Mapa de oportunidades de restauración del paisaje forestal. Unasylva 2011, 62, 47–48. [Google Scholar]
- Hillbrand, A.; Borelli, S.; Conigliaro, M.; Olivier, A. Agroforestry for Landscape Restoration: Exploring the Potential of Agroforestry to Enhance the Sustainability and Resilience of Degraded Landscapes; FAO: Rome, Italy, 2017. [Google Scholar]
- Cerrón, M.J.; del Castillo, R.J.D.; Thomas, E.; Mathez-Stiefel, S.-L.; Franco, C.M.; Mamani, C.A.; González, C.F.B.I. Experiencias de restauración en el Perú: Lecciones aprendidas; SERFOR: Bioversity, Perú; ICRAF: Lima, Perú, 2018; ISBN 9786124690839. [Google Scholar]
- Berrahmouni, N.; Regato, P.; Perfondry, M. Global Guidelines for the Restoration of Degraded Forests and Landscapes in Drylands: Building Resilience and Benefitting Livelihoods; FAO: Roma, Italia, 2015; Volume 25, ISBN 9789251089125. [Google Scholar]
- Sarmiento, F. Restoration of Equatorial Andes: The challenge for conservation of tropandean landscapes. In Biodiversity and Conservation of Neotropical Montane Forests; Churchill, S., Balslev, H., Forero, E., Luteyn, J., Eds.; The New York Botanical Garden: Nueva York, NY, USA, 1995; pp. 637–651. [Google Scholar]
- Dourojeanni, M.J. Esbozo de una nueva política forestal peruana. Rev. For. Perú 2019, 34, 4. [Google Scholar] [CrossRef]
- Sarmiento, F. Arrested succession in pastures hinders regeneration of Tropandean forests and shreds mountain landscapes. Environ. Conserv. 1997, 24, 14–23. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, H.; Yue, J.; Xie, T.; Xu, Y.; Tian, Y. Predicting shifts in the suitable climatic distribution of walnut (Juglans regia L.) in China: Maximum entropy model paves the way to forest management. Forests 2018, 9, 103. [Google Scholar] [CrossRef] [Green Version]
- Mateo, R.G.; Felicísimo, Á.M.; Muñoz, J. Modelos de distribución de especies: Una revisión sintética. Rev. Chil. Hist. Nat. 2011, 84, 217–240. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–252. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling method. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Aguirre-Gutiérrez, J.; Carvalheiro, L.G.; Polce, C.; van Loon, E.E.; Raes, N.; Reemer, M.; Biesmeijer, J.C. Fit-for-Purpose: Species Distribution Model Performance Depends on Evaluation Criteria-Dutch Hoverflies as a Case Study. PLoS ONE 2013, 8, e0063708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Bai, Y.; Wei, X.; Li, X. Distributional dynamics of a vulnerable species in response to past and future climate change: A window for conservation prospects. Peer-Rev. J. 2018, 2018, 1–25. [Google Scholar] [CrossRef]
- Cavalcante, R.A.; Nascimento, F.A.; Pereira, M.A.; Souza, D.P.; Fontana, A.P.; Cotarelli, V.M.; Oliveira, M.A.; Moura Júnior, E.G. Ampliação do conhecimento biogeográfico de Pleurophora pulchra (Lythraceae) com enfoque da conservação. Rodriguésia 2019, 70, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Vilchez, D.; Sotomayor, D.A.; Zorrilla, C. Ex situ conservation priorities for the peruvian wild tomato species (Solanum, L. SECT. Lycopersicum (MILL.) WETTST.). Ecol. Apl. 2019, 18, 171. [Google Scholar] [CrossRef]
- Alfonso-Corrado, C.; Naranjo-Luna, F.; Clark-Tapia, R.; Campos, J.E.; Rojas-Soto, O.R.; Luna-Krauletz, M.D.; Bodenhorn, B.; Gorgonio-Ramírez, M.; Pacheco-Cruz, N. Effects of environmental changes on the occurrence of Oreomunnea mexicana (Juglandaceae) in a biodiversity hotspot cloud forest. Forests 2017, 8, 261. [Google Scholar] [CrossRef]
- Qin, A.; Liu, B.; Guo, Q.; Bussmann, R.W.; Ma, F.; Jian, Z.; Xu, G.; Pei, S. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Romero-Sanchez, M.E.; Perez-Miranda, R.; Gonzalez-Hernandez, A.; Velasco-Garcia, M.V.; Velasco-Bautista, E.; Flores, A. Current and potential spatial distribution of six endangered pine species of Mexico: Towards a conservation strategy. Forests 2018, 9, 767. [Google Scholar] [CrossRef] [Green Version]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using MaxEnt modeling to predict the potential distribution of the endemic plant Rosa arabica Crép. In Egypt. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- Otieno, B.A.; Nahrung, H.F.; Steinbauer, M.J. Where did you come from? Where did you go? Investigating the origin of invasive Leptocybe species using distribution modelling. Forests 2019, 10, 115. [Google Scholar] [CrossRef] [Green Version]
- Kariyawasam, C.S.; Kumar, L.; Ratnayake, S.S. Invasive plant species establishment and range dynamics in Sri Lanka under climate change. Entropy 2019, 21, 571. [Google Scholar] [CrossRef] [Green Version]
- Antúnez, P.; Suárez-Mota, M.E.; Valenzuela-Encinas, C.; Ruiz-Aquino, F. The potential distribution of tree species in three periods of time under a climate change scenario. Forests 2018, 9, 628. [Google Scholar] [CrossRef] [Green Version]
- Frey, G.P.; West, T.A.P.; Hickler, T.; Rausch, L.; Gibbs, H.K.; Börner, J. Simulated impacts of soy and infrastructure expansion in the Brazilian Amazon: A maximum entropy approach. Forests 2018, 9, 600. [Google Scholar] [CrossRef] [Green Version]
- Vargas, R.; Clima, J. Estudios temáticos para la Zonificación Ecológica Económica del departamento de Amazonas; Instituto de Investigaciones de la Amazonía Peruana (IIAP) & Programa de Investigaciones en Cambio Climático, Desarrollo Territorial y Ambiente (PROTERRA): Chachapoyas, Perú, 2010; Volume 1, pp. 1–27. [Google Scholar]
- GRA; IIAP. Zonificación Ecológica y Económica (ZEE) del departamento de Amazonas; GRA and IIAP: Iquitos, Perú, 2010. [Google Scholar]
- Oliva, C.S.M.; Maicelo, Q.J.L.; Torres, G.C.; Bardales, E.W. Propiedades fisicoquímicas del suelo en diferentes estadios de la agricultura migratoria en el Área de Conservación Privada “Palmeras de Ocol”, distrito de Molinopampa, provincia de Chachapoyas (departamento de Amazonas). Rev. Investig. Agroproducción sustentable 2017, 1, 9–21. [Google Scholar]
- Oliva, C.S.M.; Collazos, S.R.; Espárraga, E.T.A. Efecto de las plantaciones de Pinus patula sobre las características fisicoquímicas de los suelos en áreas altoandinas de la región Amazonas. Rev. INDES 2014, 2, 28–36. [Google Scholar]
- Oliva, C.M.; Collazos, S.R.; Goñas, M.M.; Bacalla, E.; Vigo, M.C.; Vásquez, P.H.; Espinosa, L.S.T.; Maicelo, Q.J.L. Efecto de los sistemas de producción sobre las características físico-químicas de los suelos del distrito de Molinopampa, provincia de Chachapoyas, región Amazonas. Rev. INDES 2014, 2, 44–52. [Google Scholar]
- Shanee, N.; Shanee, S. Land Trafficking, Migration, and Conservation in the “No-Man’s Land” of Northeastern Peru. Trop. Conserv. Sci. 2016, 9. [Google Scholar] [CrossRef] [Green Version]
- Rojas, B.N.B.; Barboza, C.E.; Maicelo, Q.J.L.; Oliva, C.S.M.; Salas, L.R. Deforestación en la Amazonía peruana: Índices de cambios de cobertura y uso del suelo basado en SIG. Boletín De La Asociación De Geógrafos Españoles 2019, 81, 1–34. [Google Scholar] [CrossRef]
- Mendoza, C.M.E.; Salas, L.R.; Barboza, C.E. Análisis multitemporal de la deforestación usando la clasificación basada en objetos, distrito de Leymebamba (Perú). Rev. INDES 2015, 3, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Salas, L.R.; Barboza, C.E.; Oliva, C.M. Dinámica multitemporal de índices de deforestación en el distrito de Florida, departamento de Amazonas, Perú. Rev. INDES 2014, 2, 18–27. [Google Scholar]
- OSINFOR. Reportes Estadísticos: Principales Especies Forestales Maderables Aprobadas. Available online: https://observatorio.osinfor.gob.pe/Estadisticas/Home/Reportes/1 (accessed on 15 December 2019).
- Stevens, G.C. The latitudinal gradient in geographic range: How so many species coexist in the tropics. Am. Nat. 1989, 133, 240–256. [Google Scholar] [CrossRef]
- OSINFOR. Modelamiento de la distribución potencial de 18 especies forestales en el departamento de Loreto; OSINFOR: Lima, Peru, 2016; ISBN 9786124706028.
- OSINFOR. Modelamieto espacial de nichos ecológicos para la evaluación de presencia de especies forestales maderables en la Amazonía Peruana; OSINFOR: Lima, Perú, 2013.
- Figueiredo, S.M.D.M.; Venticinque, E.M.; Figueiredo, E.O.; Ferreira, E.J.L. Predição da distribuição de espécies florestais usando variáveis topográficas e de índice de vegetação no leste do Acre, Brasil. Acta Amaz. 2015, 45, 167–174. [Google Scholar] [CrossRef] [Green Version]
- OSINFOR. Distribución de las Especies Forestales del Perú; OSINFOR: Lima, Perú, 2013; ISBN 9788578110796.
- Pennington, T.; Muellner, A.N.; Wise, R. A Monograph of Cedrela (Meliaceae); Dh Books: Milborne Port, UK, 2010; ISBN 9780953813476. [Google Scholar]
- Reynel, C.; Pennington, T.D.; Pennington, R.T. Árboles del Perú; Imprenta Bellido: Lima, Perú, 2016; ISBN 9786120022320. [Google Scholar]
- Reynel, C.; Pennington, R.T.; Pennington, T.D.; Flores, C.; Daza, A. Arboles útiles de la Amazonía peruana; Tarea Grafica Educativa: Lima, Perú, 2003. [Google Scholar]
- Brako, L.; Zaruchi, J. Catalogue of the Flowering Plants and Gymnosperms of Peru; Missouri Botanical Garden: St. Louis, MO, USA, 1993. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P.; et al. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014; ISBN 9789291691432. [Google Scholar]
- Gent, P.R.; Danabasoglu, G.; Donner, L.J.; Holland, M.M.; Hunke, E.C.; Jayne, S.R.; Lawrence, D.M.; Neale, R.B.; Rasch, P.J.; Vertenstein, M.; et al. The community climate system model version 4. J. Clim. 2011, 24, 4973–4991. [Google Scholar] [CrossRef]
- van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Lamarque, J.F.; et al. The representative concentration pathways: An overview. Clim. Chang. 2011, 109, 5–31. [Google Scholar] [CrossRef]
- Rong, Z.; Zhao, C.; Liu, J.; Gao, Y.; Zang, F.; Guo, Z.; Mao, Y.; Wang, L. Modeling the effect of climate change on the potential distribution of Qinghai spruce (Picea crassifolia Kom.) in Qilian Mountains. Forests 2019, 10, 62. [Google Scholar] [CrossRef] [Green Version]
- Al-Qaddi, N.; Vessella, F.; Stephan, J.; Al-Eisawi, D.; Schirone, B. Current and future suitability areas of kermes oak (Quercus coccifera L.) in the Levant under climate change. Reg. Environ. Chang. 2017, 17, 143–156. [Google Scholar] [CrossRef]
- Mousaei, S.M.; Rundel, P.W. The impact of climate change on habitat suitability for Artemisia sieberi and Artemisia aucheri (Asteraceae)—A modeling approach. Pol. J. Ecol. 2017, 65, 97–109. [Google Scholar] [CrossRef]
- Farr, T.G.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. The Shuttle Radar Topography Mission. Rev. Geophys. 2007, 45, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Hengl, T.; De Jesus, J.M.; Heuvelink, G.B.M.; Gonzalez, M.R.; Kilibarda, M.; Blagotić, A.; Shangguan, W.; Wright, M.N.; Geng, X.; Bauer-Marschallinger, B.; et al. SoilGrids250m: Global gridded soil information based on machine learning. PLoS ONE 2017, 12, e0169748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; García, M.J.R.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 027–046. [Google Scholar] [CrossRef]
- De Marco, J.P.; Corrêa, N.C. Evaluating collinearity effects on species distribution models: An approach based on virtual species simulation. PLoS ONE 2018, 13, e02022403. [Google Scholar] [CrossRef] [PubMed]
- Laurente, M. Modeling the Effects of Climate Change on the Distribution of Cedrela odorata L. «Cedro» in the Peruvian Amazon. Biology 2015, 13, 213–224. [Google Scholar]
- Manel, S.; Ceri, W.H.; Ormerod, S.J. Evaluating presence–absence models in ecology: The need to account for prevalence. J. Appl. Ecol. 2001, 38, 921–931. [Google Scholar] [CrossRef]
- Hanley, J.A.; McNeil, B.J. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 1982, 143, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Araujo, M.; Pearson, R.; Thuiller, W.; Erhard, M. Validation of species-climate impact models under climate change. Glob. Chang. Biol. 2005, 11, 1504–1513. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Tao, J. Predicting the potential distribution of Paeonia veitchii (Paeoniaceae) in China by incorporating climate change into a maxent model. Forests 2019, 10, 190. [Google Scholar] [CrossRef] [Green Version]
- Leguía, E.; Soudre, M.; Rugnitz, M. Predicción y evaluación del impacto del cambio climático sobre los sistemas agroforestales en la amazonia peruana y andina ecuatoriana; IIAP, ICRAF: Pucallpa, Perú, 2010. [Google Scholar]
- Guzmán, M.A.N.; Chipana, C.A.J.; Apaza, J.M.I. Modeling ecological niches of threatened flora for climate change scenarios in Tacna department-Peru. Colomb. For. 2020, 23. [Google Scholar] [CrossRef]
- Estrada-Contreras, I.; Equihua, M.; Laborde, J.; Meyer, E.M.; Sanchez-Velasquez, L.R. Current and future distribution of the tropical tree Cedrela Odorata L. in Mexico under Climate Change Scenarios Using Maxlike. PLoS ONE 2016, 11, e0164178. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhao, C.; Feng, Z. A study of the impact of climate change on the potential distribution of Qinghai spruce (Picea crassifolia) in Qilian Mountains. Acta Ecol. Sin. 2009, 29, 278–285. [Google Scholar] [CrossRef]
- Ramos, J.H.; Santos, R.R.; Ramos, A.H.; Cuevas, X.G.; Hernández-Máximo, E.; Uicab, J.V.C.; López, D.S. Historical, current and future distribution of Cedrela odorata in Mexico. Acta Bot. Mex. 2018, 2018, 117–134. [Google Scholar] [CrossRef]
- Lamont, B.B.; Connell, S.W. Biogeography of Banksia in southwestern Australia. J. Biogeogr. 1996, 23, 295–309. [Google Scholar] [CrossRef]
- Sarmiento, F.O.; Kooperman, G.J. A Socio-Hydrological Perspective on Recent and Future Precipitation Changes Over Tropical Montane Cloud Forests in the Andes. Front. Earth Sci. 2019, 7, 1–6. [Google Scholar] [CrossRef]
- Sarmiento, F.O. Landscape Regeneration by Seeds and Successional Pathways to Restore Fragile Tropandean Slopelands. Mt. Res. Dev. 1997, 17, 239. [Google Scholar] [CrossRef] [Green Version]
- SERNANP. Servicio Nacional de Áreas Naturales Protegidas por el Estado. Servicios y Recursos. Available online: https://www.geoidep.gob.pe/servicio-nacional-de-areas-naturales-protegidas-por-el-estado (accessed on 15 April 2019).
- SPDA. Áreas de conservación privada en el Perú: Anances y propuestas a 20 años de su creación; Monteferri, B., Ed.; Sociedad Peruana de Derecho Ambiental: Lima, Perú, 2019; ISBN 9786124261442. [Google Scholar]
- Salinas-Rodríguez, M.M.; Sajama, M.J.; Gutiérrez-Ortega, J.S.; Ortega-Baes, P.; Estrada-Castillón, A.E. Identification of endemic vascular plant species hotspots and the effectiveness of the protected areas for their conservation in Sierra Madre Oriental, Mexico. J. Nat. Conserv. 2018, 46, 6–27. [Google Scholar] [CrossRef]
- Rong-Cai, Y.; Yeh, F.C. Genetic consequences of in situ and ex situ conservation of forest trees. For. Chron. 1992, 68, 720–729. [Google Scholar]
- Bonn Challenge. A Global Effort to Bring 150 Million Hectares of Deforested and Degraded Land into Restoration by 2020 and 350 Million Hectares by 2030. Available online: https://www.bonnchallenge (accessed on 24 March 2020).
- CBD. Decision Adopted By the Conference of the Parties to the Convention on Biological Diversity at Its Seventh Meeting: XI/16. Ecosystem restoration. In Proceedings of the Conference of the Parties to the Convention on Biological Diversity, Pyeongchang, Korea, 6–17 October 2014; Convention on Biological Diversity: Hyderabad, India, 2012. [Google Scholar]
- Initiative 20 × 20. Healthy Lands for Food, Water and Climate. A Country-Led Effort that Aims to Change the Dynamics of Land Degradation in Latin America and the Caribbean. Available online: https://initiative20x20 (accessed on 24 March 2020).
- International Union for Conservation of Nature. Forest Landscape Restoration. Available online: https://www.iucn:theme/forests/our-work/forest-landscape-restoration (accessed on 24 March 2020).
- Holl, K.D. Restoring tropical forests from the bottom up: How can ambitious forest restoration targets be implemented on the ground? Science 2017, 355, 455–456. [Google Scholar] [CrossRef] [Green Version]
- MINAM. Plan Nacional De Acción Ambiental-Planaa Perú: 2011–2021, 2nd ed.; MINAM: Lima, Perú, 2011.
- SERFOR. Ley Forestal y de Fauna Silvestre N 29763 y sus reglamentos, 2nd ed.; SERFOR: Lima, Perú, 2015. [Google Scholar]
- Sears, R.R.; Cronkleton, P.; Polo Villanueva, F.; Miranda Ruiz, M.; Pérez-Ojeda del Arco, M. Farm-forestry in the Peruvian Amazon and the feasibility of its regulation through forest policy reform. For. Policy Econ. 2018, 87, 49–58. [Google Scholar] [CrossRef]
- Mackey, B.; Kormos, C.F.; Keith, H.; Moomaw, W.R.; Houghton, R.A.; Mittermeier, R.A.; Hole, D.; Hugh, S. Understanding the importance of primary tropical forest protection as a mitigation strategy. Mitig. Adapt. Strateg. Glob. Chang. 2020. [Google Scholar] [CrossRef] [Green Version]
- Mostacero-Leon, J.; Mejia-Coico, F.; Pelaez-Pelaez, F.; Charcape-Ravelo, M. Especies madereras nativas del norte del perú. Rebiol 1998, 16, 67–78. [Google Scholar]
- Baluarte, V.J.R.; Alvarez, G.J.G. Modelamiento del crecimiento de tornillo Cedrelinga catenaeformis Ducke en plantaciones en Jenaro Herrera, departamento de Loreto, Perú. Folia Amaz. 2015, 24, 21. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Variable | Description | Min | Max | Mean | SD | Species 1 |
|---|---|---|---|---|---|---|---|
| Climate | bio01 | Annual Mean Temperature (°C) | 7.26 | 26.95 | 20.85 | 4.53 | e |
| bio02 | Mean Diurnal Range (°C) | 8.41 | 14.47 | 11.18 | 0.79 | c; e | |
| bio03 | Isothermality | 77.33 | 94.50 | 88.15 | 2.13 | a; b; d; e | |
| bio04 | Temperature Seasonality (°C) | 23.07 | 97.64 | 40.36 | 9.54 | c | |
| bio05 | Max Temperature of Warmest Month (°C) | 14.20 | 32.70 | 27.07 | 4.30 | ||
| bio06 | Min Temperature of Coldest Month (°C) | 0.00 | 21.10 | 14.38 | 4.74 | c | |
| bio07 | Temperature Annual Range (°C) | 10.00 | 15.70 | 12.68 | 0.76 | ||
| bio08 | Mean Temperature of Wettest Quarter (°C) | 7.47 | 26.87 | 20.90 | 4.52 | ||
| bio09 | Mean Temperature of Driest Quarter (°C) | 6.48 | 26.87 | 20.51 | 4.69 | b; d | |
| bio10 | Mean Temperature of Warmest Quarter (°C) | 7.85 | 27.25 | 21.27 | 4.49 | a | |
| bio11 | Mean Temperature of Coldest Quarter (°C) | 6.48 | 26.50 | 20.28 | 4.54 | ||
| bio12 | Annual Precipitation (mm) | 382.00 | 2611.00 | 1568.98 | 543.17 | ||
| bio13 | Precipitation of Wettest Month (mm) | 51.00 | 280.00 | 182.23 | 49.95 | ||
| bio14 | Precipitation of Driest Month (mm) | 9.00 | 174.00 | 91.51 | 45.59 | ||
| bio15 | Precipitation Seasonality (mm) | 12.64 | 62.53 | 25.11 | 9.84 | c; d | |
| bio16 | Precipitation of Wettest Quarter (mm) | 134.00 | 812.00 | 504.18 | 157.96 | a; b; c; e | |
| bio17 | Precipitation of Driest Quarter (mm) | 36.00 | 556.00 | 294.58 | 141.97 | ||
| bio18 | Precipitation of Warmest Quarter (mm) | 125.00 | 608.00 | 402.07 | 117.49 | a; c; d; e | |
| bio19 | Precipitation of Coldest Quarter (mm) | 36.00 | 702.00 | 352.30 | 190.15 | c | |
| rad | Solar radiation (kJ m−2 day−1) | 12,011.58 | 15,285.75 | 13,742.68 | 526.64 | d; e | |
| Topography | dem | Elevation above mean sea level (m.a.s.l.) | 155.00 | 4919.00 | 1368.60 | 929.43 | a; b; c; d; e |
| slope | Terrain tilt (°) | 0.01 | 76.27 | 12.85 | 8.63 | a | |
| aspect | Cardinal slope direction (°) | 0.00 | 360.00 | 177.54 | 103.49 | a; b; d; e | |
| Soil | ph | pH × 10 un KCl at 0.30 m | 37.00 | 67.00 | 44.35 | 3.66 | a; b; c; d; e |
| cic | Cation exchange capacity at 0.30 m (cmolc kg−1) | 6.00 | 61.00 | 18.83 | 6.29 | a; b; d; e | |
| cot | Organic carbon content at 0.15 m (g kg−1) | 4.00 | 269.00 | 52.12 | 31.16 | d |
| Scenario | A. leiocarpa | C. decandra | C. montana | C. cateniformis | C. pentandra | |
|---|---|---|---|---|---|---|
| Current | 0.954 | 0.958 | 0.868 | 0.914 | 0.952 | |
| CCSM4 2050 | RCP 2.6 | 0.965 | 0.959 | 0.875 | 0.902 | 0.963 |
| RCP 4.5 | 0.963 | 0.956 | 0.858 | 0.899 | 0.962 | |
| RCP 6.0 | 0.964 | 0.952 | 0.860 | 0.903 | 0.960 | |
| RCP 8.5 | 0.961 | 0.965 | 0.861 | 0.908 | 0.960 | |
| CCSM4 2070 | RCP 2.6 | 0.965 | 0.955 | 0.858 | 0.909 | 0.958 |
| RCP 4.5 | 0.963 | 0.959 | 0.870 | 0.904 | 0.960 | |
| RCP 6.0 | 0.964 | 0.955 | 0.867 | 0.905 | 0.961 | |
| RCP 8.5 | 0.961 | 0.944 | 0.856 | 0.912 | 0.959 | |
| Mean | 0.962 | 0.956 | 0.864 | 0.906 | 0.959 | |
| Habitat Potential | Current (km2) | CCSM4 2050 (%) | CCSM4 2070 (%) 1 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| RCP 2.6 | RCP 4.5 | RCP 6.0 | RCP 8.5 | RCP 2.6 | RCP 4.5 | RCP 6.0 | RCP 8.5 | ||
| C. cateniformis | |||||||||
| High | 1194.76 | −1.4 | −12.6 | −13.7 | −11.7 | −19.1 (−17.9) | −17.4 (−5.5) | −20.3 (−7.6) | −14.6 (−3.3) |
| Moderate | 2666.85 | 56.4 | 88.8 | 67.5 | 50.2 | 43.2 (−8.5) | 66.9 (−11.6) | 64.7 (−1.6) | 34.9 (−10.2) |
| Low | 4757.6 | 25.8 | 24.8 | 4.9 | 33.6 | 1.4 (−19.4) | 16.7 (−6.5) | 13.5 (8.2) | 10.1 (−17.6) |
| Total | 8619.21 | 31.5 | 39.4 | 21.7 | 32.5 | 11.5 (−15.2) | 27.5 (−8.5) | 24.7 (2.5) | 14.4 (−13.7) |
| C. pentandra | |||||||||
| High | 584.39 | −23.7 | −21.5 | −21.6 | −15.1 | −6.0 (23.3) | −24.2 (−3.5) | −22.2 (−0.8) | −8.9 (7.3) |
| Moderate | 1966.59 | −24.8 | −24.6 | −16.9 | −18.0 | −16.1 (11.5) | −13.3 (15.0) | −18.2 (−1.6) | −23.1 (−6.2) |
| Low | 3128.21 | −44.8 | −40.9 | −44.4 | −43.7 | −42.7 (3.9) | −43.0 (−3.5) | −46.5 (−3.7) | −43.9 (−0.5) |
| Total | 5679.19 | −35.7 | −33.3 | −32.5 | −31.9 | −29.7 (9.3) | −30.8 (3.7) | −34.2 (−2.5) | −33.1 (−1.8) |
| A. leiocarpa | |||||||||
| High | 761.55 | −12.8 | −12.8 | −7.8 | 0.2 | −7.5 (6.1) | −0.2 (14.4) | −13.3 (−6.0) | −0.9 (−1.1) |
| Moderate | 2449.94 | −22.6 | −21.4 | −13.3 | −11.2 | −21.4 (1.6) | −18.2 (4.0) | −15.8 (−2.9) | −19.3 (−9.1) |
| Low | 2822.38 | −34.0 | −36.0 | −34.6 | −37 | −37.0 (−4.5) | −37.2 (−1.9) | −40.7 (−9.4) | −33.0 (6.4) |
| Total | 6033.88 | −26.7 | −27.1 | −22.6 | −21.8 | −27.0 (−0.3) | −24.8 (3.2) | −27.1 (−5.9) | −23.4 (−2.0) |
| C. decandra | |||||||||
| High | 761.67 | −27.9 | −7.0 | 16.1 | −8.2 | 16.1 (60.9) | 12.7 (21.1) | −1.7 (−15.4) | 42.5 (55.3) |
| Moderate | 1885.82 | −17.6 | −8.7 | 7.0 | −26.1 | −7.4 (12.4) | −10.5 (−2.0) | −23.1 (−28.2) | 12.9 (52.7) |
| Low | 2903.42 | −15.9 | −9.4 | 15.9 | −30.0 | −30.5 (−17.3) | −32.9 (−26.0) | −27.3 (−37.3) | 16.0 (65.8) |
| Total | 5550.91 | −18.1 | −8.8 | 12.9 | −25.7 | −16.3 (2.3) | −19.1 (−11.2) | −22.4 (−31.3) | 18.6 (59.6) |
| C. montana | |||||||||
| High | 2625.42 | −7.9 | −9.1 | −13.4 | −8.7 | −7.5 (0.5) | −10.1 (−1.1) | −12.0 (1.6) | −9.7 (−1.0) |
| Moderate | 5832.68 | −7.4 | 2.6 | 4.3 | −4.3 | 3.3 (11.5) | −5.6 (−8.0) | −3.7 (−7.7) | 14.3 (19.5) |
| Low | 8649.03 | −17.4 | −15.7 | −15.1 | −13.8 | −15.7 (2.0) | −16.2 (−0.6) | −13.6 (1.8) | −17.8 (−4.7) |
| Total | 17107.1 | −12.6 | −8.4 | −8.2 | −9.8 | −8.0 (5.2) | −11.6 (−3.5) | −10.0 (−1.9) | −5.6 (4.6) |
| Habitat Potential | Nº Species | Current (km2) | CCSM4 2050 (%) | CCSM4 2070 (%) 1 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| RCP 2.6 | RCP 4.5 | RCP 6.0 | RCP 8.5 | RCP 2.6 | RCP 4.5 | RCP 6.0 | RCP 8.5 | |||
| High | 3 or 4 | 182.26 | −20.3 | 30.8 | −56.2 | −38.3 | −41.5 (-26.6) | −42.6 (-56.2) | −14.6 (95.0) | −30.7 (12.3) |
| Moderate | 2 | 547.58 | −16.7 | −27.3 | −11.8 | −5.2 | −0.6 (19.3) | 8.3 (49.0) | −25.0 (−14.9) | 12.4 (18.5) |
| Low | 1 | 4275.76 | −8.8 | −13.0 | −3.1 | −5.9 | −3.7 (5.6) | −8.7 (5.0) | −10.9 (−8.0) | −3.1 (2.9) |
| Total | 5005.60 | −10.1 | −13.0 | −6.0 | −7.0 | −4.8 (5.9) | −8.1 (5.7) | -12.5 (-6.9) | −2.4 (4.9) | |
| Specie | Habitat Potential | Area (%) by Category and Class of Degraded Area | ||||
|---|---|---|---|---|---|---|
| High | Medium | Low | Total (%) | |||
| Deforestation 2001–2017 | Negative NPP and Forest Fragments | Negative NPP or Changes in Vegetation Cover | Forest Fragments | |||
| C. cateniformis | High | 3.2 (4) | 14.3 (11.05) | 0.0 (0.0) | 20 (2.76) | 37.5 (3.89) |
| Moderate | 2.9 (8.1) | 10.9 (18.72) | 0.0 (0.01) | 20.6 (6.34) | 34.4 (7.94) | |
| Low | 3.6 (17.71) | 11.2 (34.41) | 0.0 (0.01) | 19.3 (10.61) | 34.1 (14.05) | |
| Total | 3.3 (29.81) | 11.6 (64.18) | 0.0 (0.02) | 19.8 (19.72) | 34.6 (25.88) | |
| C. pentandra | High | 6.6 (4.01) | 19.9 (7.51) | 0.0 (0.0) | 23.9 (1.61) | 50.4 (2.55) |
| Moderate | 6.5 (13.28) | 18.1 (22.89) | 0.0 (0.0) | 20.1 (4.57) | 44.6 (7.61) | |
| Low | 3.1 (10.19) | 9.5 (19.13) | 0.0 (0.0) | 18.8 (6.82) | 31.4 (8.53) | |
| Total | 4.6 (27.48) | 13.5 (49.53) | 0.0 (0.0) | 19.8 (13) | 38 (18.69) | |
| A. leiocarpa | High | 5.1 (4.01) | 16.1 (7.93) | 0.0 (0.0) | 18.3 (1.61) | 39.5 (2.61) |
| Moderate | 5.2 (13.24) | 14.3 (22.58) | 0.0 (0.0) | 18.4 (5.2) | 37.8 (8.04) | |
| Low | 3.3 (9.6) | 9.7 (17.65) | 0.0 (0.0) | 19.1 (6.24) | 32.1 (7.85) | |
| Total | 4.3 (26.85) | 12.4 (48.16) | 0.0 (0.0) | 18.7 (13.05) | 35.3 (18.49) | |
| C. decandra | High | 1.9 (1.53) | 7.6 (3.74) | 0.0 (0.0) | 16 (1.41) | 25.5 (1.69) |
| Moderate | 3.7 (7.21) | 12.5 (15.25) | 0.0 (0.0) | 17.1 (3.73) | 33.3 (5.45) | |
| Low | 4.3 (13.11) | 12.5 (23.49) | 0.0 (0.0) | 18.4 (6.2) | 35.3 (8.89) | |
| Total | 3.8 (21.84) | 11.9 (42.48) | 0.0 (0.0) | 17.6 (11.34) | 33.3 (16.03) | |
| C. montana | High | 5 (13.67) | 1.8 (3.07) | 0.0 (0.3) | 39.7 (12.05) | 46.5 (10.59) |
| Moderate | 4.7 (28.77) | 1.7 (6.39) | 0.3 (4.35) | 36.9 (24.93) | 43.6 (22.07) | |
| Low | 1.6 (14.3) | 0.9 (5.14) | 1.3 (29.68) | 23.4 (23.47) | 27.3 (20.44) | |
| Total | 3.2 (56.74) | 1.3 (14.6) | 0.8 (34.33) | 30.5 (60.45) | 35.8 (53.11) | |
| Combined TFS | High | 2.8 (0.54) | 10.2 (1.2) | 0.0 (0.0) | 18.4 (0.39) | 31.4 (0.5) |
| Moderate | 4.2 (2.41) | 14.6 (5.17) | 0.0 (0.0) | 19.4 (1.23) | 38.3 (1.82) | |
| Low | 4.7 (20.73) | 7 (19.26) | 0.0 (0.3) | 31.9 (15.78) | 43.6 (16.15) | |
| Total | 4.5 (23.68) | 7.9 (25.62) | 0.0 (0.3) | 30 (17.4) | 42.5 (18.46) | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rojas Briceño, N.B.; Cotrina Sánchez, D.A.; Barboza Castillo, E.; Barrena Gurbillón, M.Á.; Sarmiento, F.O.; Sotomayor, D.A.; Oliva, M.; Salas López, R. Current and Future Distribution of Five Timber Forest Species in Amazonas, Northeast Peru: Contributions towards a Restoration Strategy. Diversity 2020, 12, 305. https://doi.org/10.3390/d12080305
Rojas Briceño NB, Cotrina Sánchez DA, Barboza Castillo E, Barrena Gurbillón MÁ, Sarmiento FO, Sotomayor DA, Oliva M, Salas López R. Current and Future Distribution of Five Timber Forest Species in Amazonas, Northeast Peru: Contributions towards a Restoration Strategy. Diversity. 2020; 12(8):305. https://doi.org/10.3390/d12080305
Chicago/Turabian StyleRojas Briceño, Nilton B., Dany A. Cotrina Sánchez, Elgar Barboza Castillo, Miguel Ángel Barrena Gurbillón, Fausto O. Sarmiento, Diego A. Sotomayor, Manuel Oliva, and Rolando Salas López. 2020. "Current and Future Distribution of Five Timber Forest Species in Amazonas, Northeast Peru: Contributions towards a Restoration Strategy" Diversity 12, no. 8: 305. https://doi.org/10.3390/d12080305
APA StyleRojas Briceño, N. B., Cotrina Sánchez, D. A., Barboza Castillo, E., Barrena Gurbillón, M. Á., Sarmiento, F. O., Sotomayor, D. A., Oliva, M., & Salas López, R. (2020). Current and Future Distribution of Five Timber Forest Species in Amazonas, Northeast Peru: Contributions towards a Restoration Strategy. Diversity, 12(8), 305. https://doi.org/10.3390/d12080305