Morphometric Analyses Distinguish Wabash Pigtoe (Fusconaia flava) and Round Pigtoe (Pleurobema sintoxia) Mussels

Abstract
:1. Introduction
2. Methods
2.1. Specimen Collections
2.2. DNA Barcoding
2.3. Traditional Morphometrics
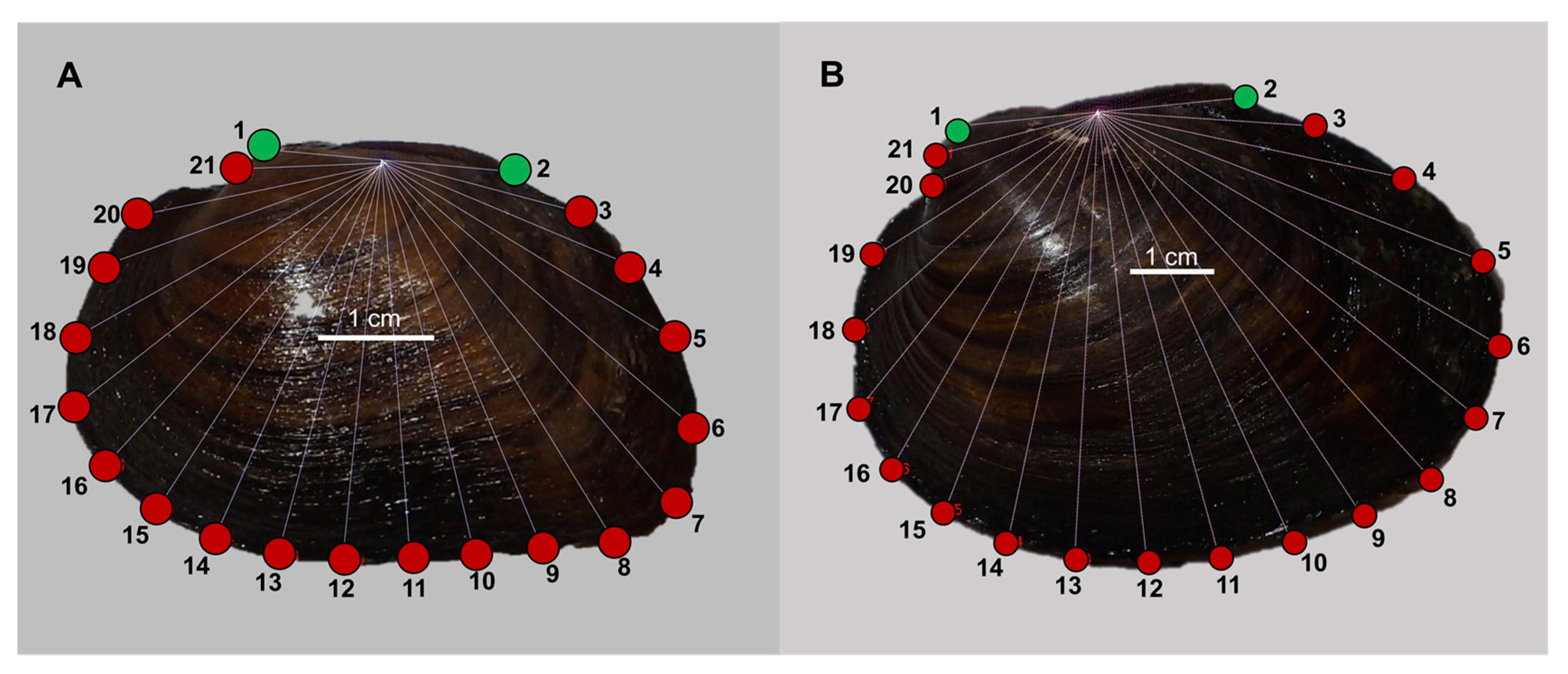
2.4. Geometric Morphometrics
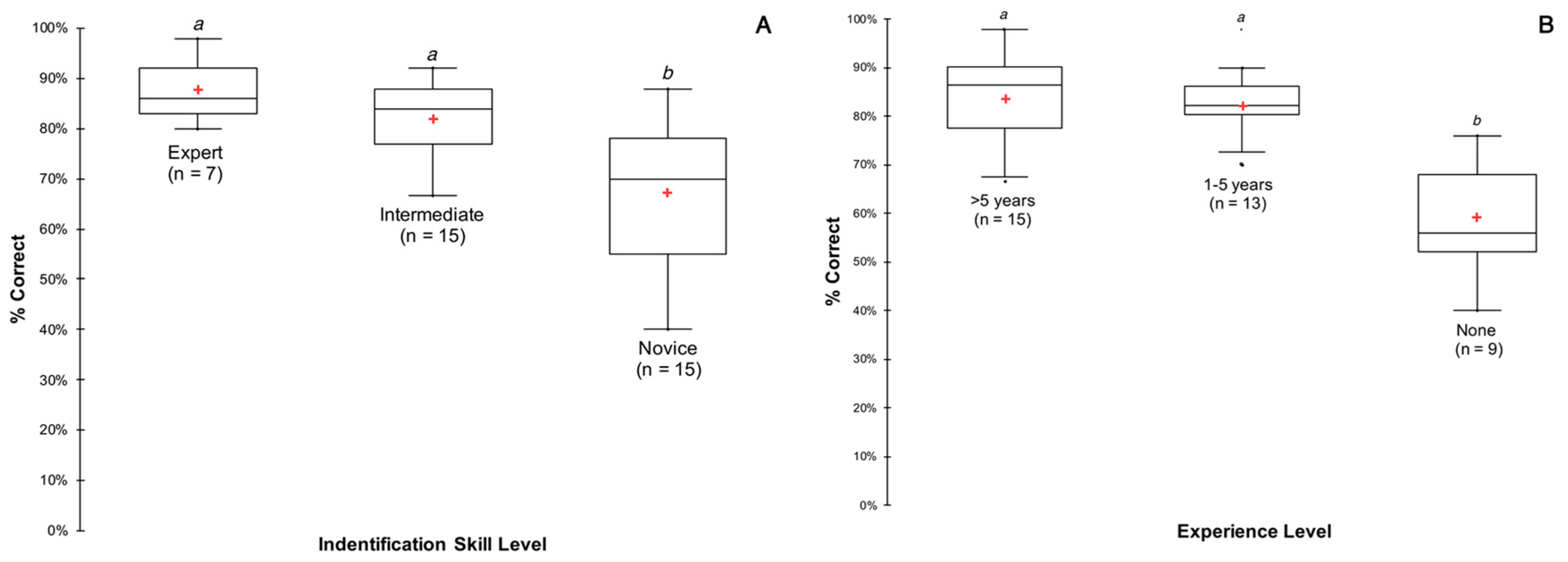
2.5. Identification Quiz
3. Results and Discussion
3.1. DNA Barcoding
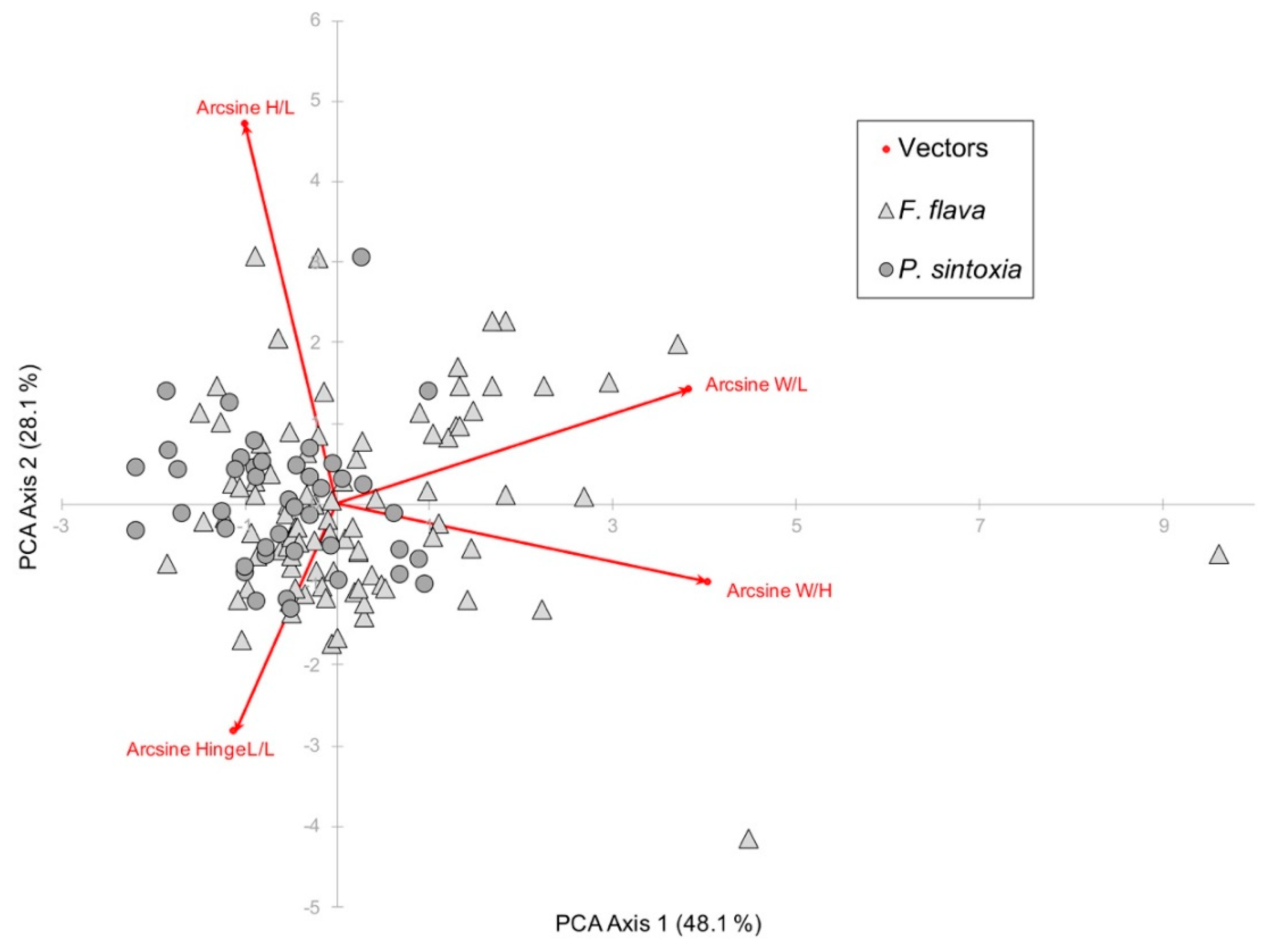
3.2. Traditional Morphometrics
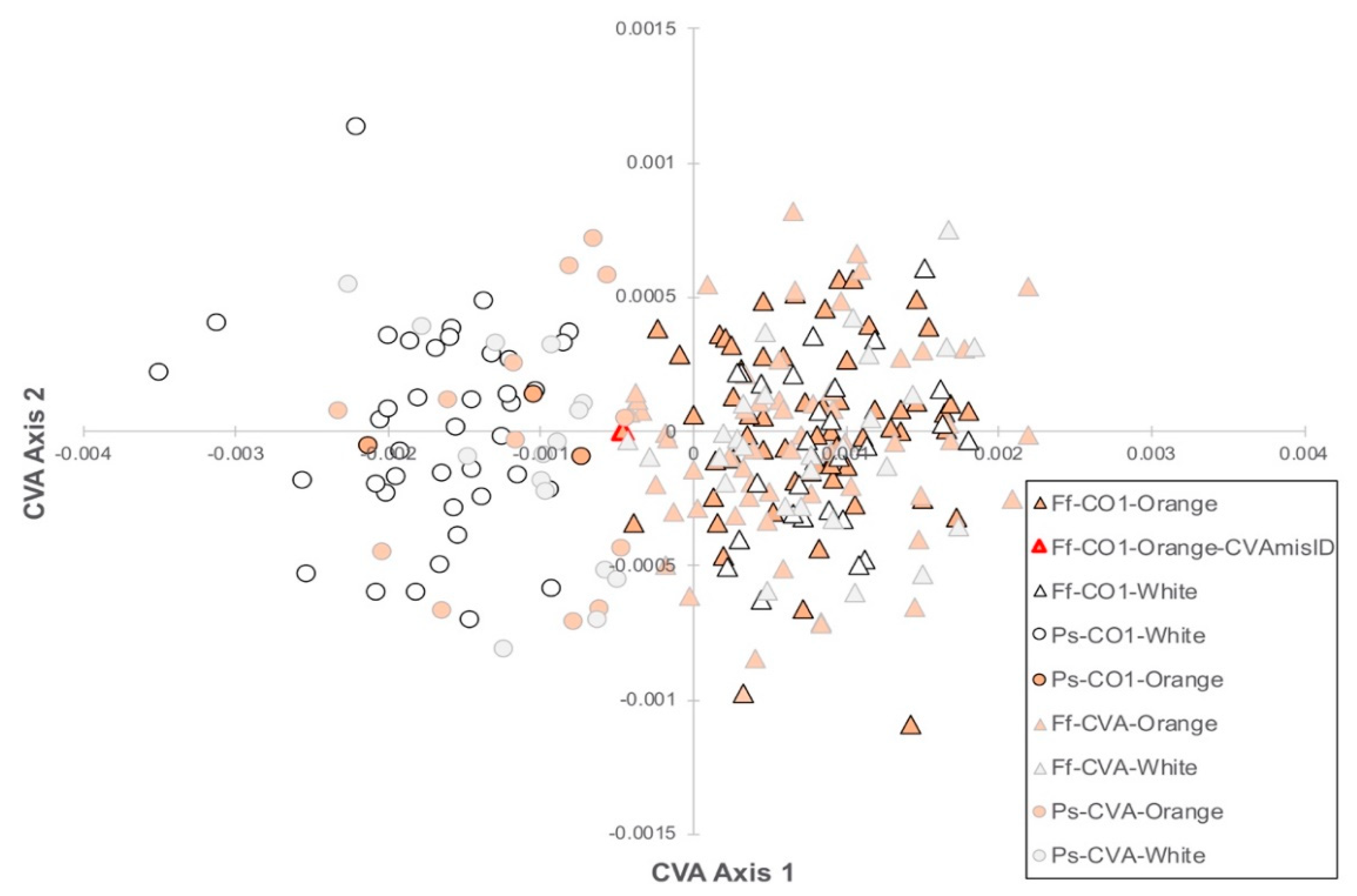
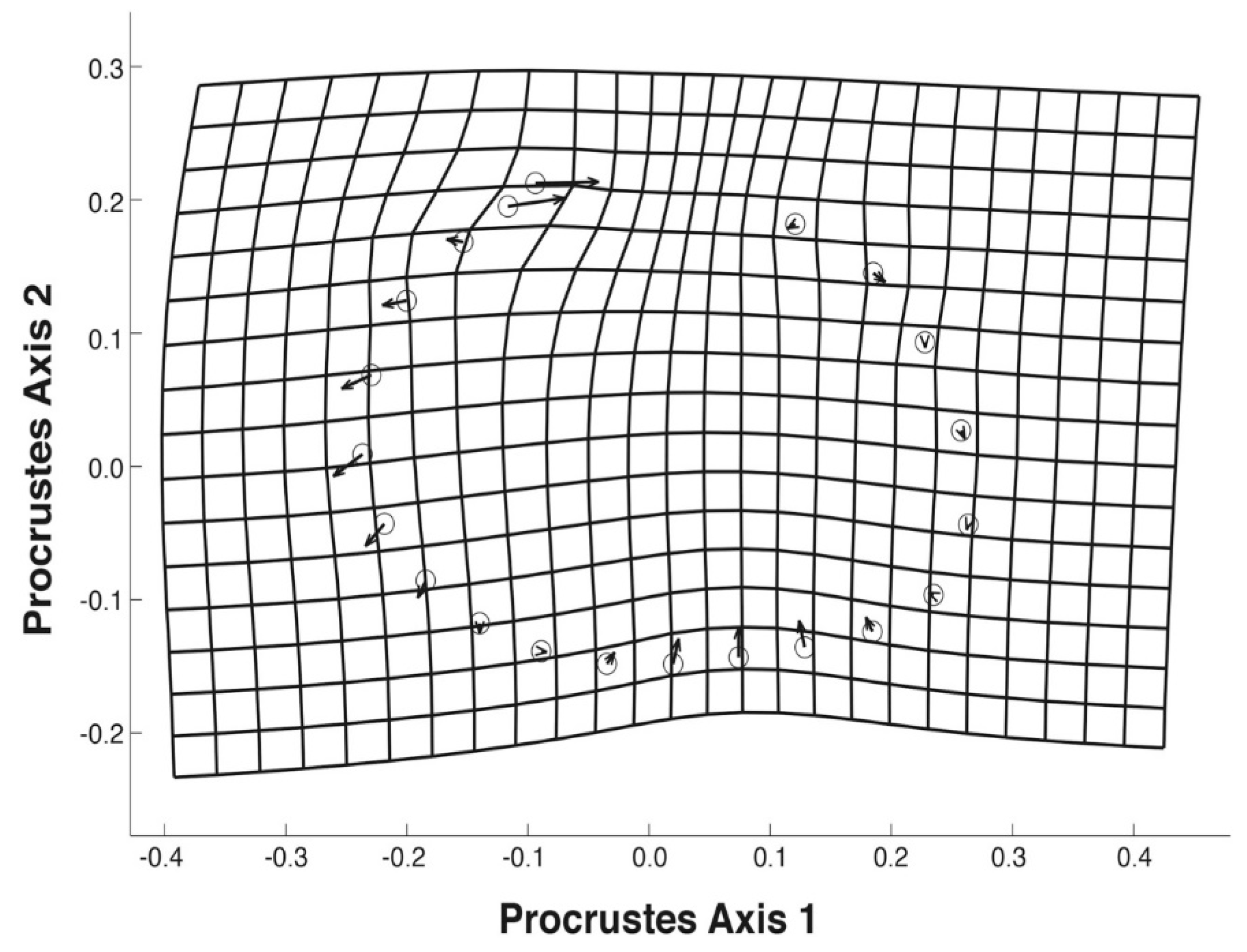
3.3. Geometric Morphometrics
3.4. Foot Color
3.5. Identification Quiz
3.6. Conservation Implications and Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Haag, W.R. North American Freshwater Mussels: Natural History, Ecology, and Conservation; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Ortmann, A.E. The geographical distribution of freshwater decapods and its bearing upon ancient geography. Proc. Am. Phil. Soc. 1920, 41, 267–400. [Google Scholar]
- Shea, C.P.; Peterson, J.T.; Wisniewski, J.M.; Johnson, N.A. Misidentification of freshwater mussel species (Bivalvia: Unionidae): Contributing factors, management implications, and potential solutions. J. N. Am. Benthol. Soc. 2011, 30, 446–458. [Google Scholar] [CrossRef]
- Inoue, K.; Hayes, D.M.; Harris, J.L.; Johnson, N.A.; Morrison, C.L.; Eackles, M.S.; King, T.L.; Jones, J.W.; Hallerman, E.M.; Christian, A.D.; et al. The Pleurobemini (Bivalvia: Unionida) revisited: Molecular species delineation using a mitochondrial DNA gene reveals multiple conspecifics and undescribed species. Invert. Syst. 2018, 32, 689–702. [Google Scholar] [CrossRef]
- Watters, G.T.; Hoggarth, M.A.; Stansbery, D.H. The Freshwater Mussels of Ohio; The Ohio State University: Columbus, OH, USA, 2009. [Google Scholar]
- Mulcrone, R.S.; Rathburn, J.E. Field Guide to the Freshwater Mussels of Michigan; Michigan Department of Natural Resources: Lansing, MI, USA, 2018. [Google Scholar]
- Pieri, A.M.; Inoue, K.; Johnson, N.A.; Smith, C.H.; Harris, J.L.; Robertson, C.; Randklev, C.R. Molecular and morphometric analyses reveal cryptic diversity within freshwater mussels (Bivalvia: Unionidae) of the western Gulf coastal drainages of the USA. Biol. J. Linn. Soc. 2018, 124, 261–277. [Google Scholar] [CrossRef]
- COSEWIC Assessment and Status Report on the Round Pigtoe Pleurobema sintoxia in Canada. Committee on the Status of Endangered Wildlife in Canada, Ottawa, ON Canada, 2004; vi + 33 pp. Available online: https://www.sararegistry.gc.ca/virtual_sara/files/cosewic/sr%5Fround%5Fpigtoe%5Fe%2Epdf (accessed on 10 January 2019).
- Cummings, K.S.; Mayer, K.S. Field Guide to Freshwater Mussels of the Midwest; Illinois Natural History Survey: Champaign, IL USA, 1992; Manual 5, 194p; Available online: https://wwv.inhs.illinois.edu/collections/mollusk/publications/guide/ (accessed on 10 January 2019).
- Newth, J.L.; Wood, K.A.; McDonald, R.A.; Nuno, A.; Semenov, I.; Chistyakov, A.; Mikhaylova, G.; Bearhop, S.; Belousova, A.; Glazov, P.; et al. Conservation implications of misidentification and killing of protected species. Conserv. Sci. Prac. 2019, 1, e24. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lon. Ser. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Campbell, D.C.; Johnson, P.D.; Williams, J.D.; Rindsberg, A.K.; Serb, J.M.; Small, K.K.; Lydeard, C. Identification of ‘extinct’ freshwater mussel species using DNA barcoding. Mol. Ecol. Res. 2008, 8, 711–724. [Google Scholar] [CrossRef]
- Boyer, S.L.; Howe, A.A.; Juergens, N.W.; Hove, M.C. A DNA-barcoding approach to identifying juvenile freshwater mussels (Bivalvia: Unionidae) recovered from naturally infested fishes. J. N. Am. Benthol. Soc. 2011, 30, 182–194. [Google Scholar] [CrossRef]
- Wu, R.-W.; Liu, Y.-T.; Sa, W.; Liu, X.-J.; Zanatta, D.T.; Roe, K.J.; Song, X.-L.; An, C.-T.; Wu, X.-P. Testing the utility of DNA barcodes and a preliminary phylogenetic framework for Chinese freshwater mussels (Bivalvia: Unionidae) from the middle and lower Yangtze River. PLoS ONE 2018, 13, e0200956. [Google Scholar] [CrossRef]
- Webster, M.; Sheets, H.D. A practical introduction to landmark-based geometric morphometrics. In Quantitative Methods in Paleontology; Alroy, J., Hunt, G., Eds.; Yale University Press: New Haven, CT, USA, 2010; Volume 16, pp. 163–188. [Google Scholar]
- Inoue, K.; Hayes, D.M.; Harris, J.L.; Christian, A.D. Phylogenetic and morphometric analyses reveal ecophenotypic plasticity in freshwater mussels Obovaria jacksoniana and Villosa arkansasensis (Bivalvia: Unionidae). Ecol. Evol. 2013, 3, 2670–2683. [Google Scholar] [CrossRef]
- Inoue, K.; McQueen, A.L.; Harris, J.L.; Berg, D.J. Molecular phylogenetics and morphological variation reveal recent speciation in freshwater mussels of the genera Arcidens and Arkansia (Bivalvia: Unionidae). Biol. J. Linn. Soc. 2014, 112, 535–545. [Google Scholar] [CrossRef] [Green Version]
- Beauchamp, K.A.; Beyett, T.W.; Scott, M.W.; Zanatta, D.T. Detection of hybrid Pyganodon grandis and P. lacustris (Bivalvia: Unionidae) using F- and M-lineage mtDNA sequences and geometric morphometrics. J. Mollusc. Stud. 2020, 86, 233–239. [Google Scholar] [CrossRef]
- Beyett, T.W.; McNichols-O’Rourke, K.; Morris, T.J.; Zanatta, D.T. Use of morphometric analyses and DNA barcoding to distinguish Truncilla donaciformis and Truncilla truncata (Bivalvia: Unionidae). Freshw. Moll. Biol. Cons. 2020, in press. [Google Scholar]
- Henley, W.F.; Grobler, J.P.; Neves, R.J. Non-invasive method to obtain DNA from freshwater mussels (Bivalvia: Unionidae). J. Shellfish Res. 2006, 25, 975–977. [Google Scholar]
- Berg, D.J.; Haag, W.R.; Guttman, S.I.; Sickel, J.B. Mantle biopsy: A technique for nondestructive tissue-sampling of freshwater mussels. J. N. Am. Benth. Soc. 1995, 14, 577–581. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Plainview, NY, USA, 1989. [Google Scholar]
- Campbell, D.C.; Serb, J.M.; Buhay, J.E.; Roe, K.J.; Minton, R.L.; Lydeard, C. Phylogeny of north American amblemines (Bivalvia: Unionoida): Prodigious polyphyly proves pervasive across genera. Invert. Biol. 2005, 124, 131–164. [Google Scholar] [CrossRef]
- XLSTAT. XLSTAT: Data Analysis and Statistical Solutions for Microsoft Excel; Addinsoft: Paris, France, 2018. [Google Scholar]
- Sheets, H.D. Integrated Morphometrics Package (IMP), Version 8, 2014. Available online: https://www.animal-behaviour.de/imp/ (accessed on 3 May 2020).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site (River) | Latitude | Longitude | F. flava | P. sintoxia | ||||
|---|---|---|---|---|---|---|---|---|
| N Initial Field ID | N COI Confirmed ID | N Morphometric ID | N Initial Field ID | N COI Confirmed ID | N Morphometric ID | |||
| Belle River, MI | 42.774682 | −82.551841 | 15 | 16 | 15 | 2 | 1 | 2 |
| Clinton River, MI | 42.628098 | −83.395628 | 4 | 0 | 0 | 0 | 4 | 4 |
| Kalamazoo River, MI-site 5 | 42.338800 | −85.234710 | 4 | 3 | 3 | 0 | 1 | 1 |
| Kalamazoo River, MI-site 9 | 42.291040 | −85.559610 | 18 | 18 | 18 | 0 | 0 | 0 |
| Pine River, MI-LJR | 43.398068 | −84.836731 | 3 | 2 | 2 | 1 | 2 | 2 |
| Pine River, MI-site 15 | 43.335373 | −84.712585 | 15 | 6 | 6 | 5 | 14 | 14 |
| Pine River, MI-site 8 | 43.425904 | −84.835667 | 11 | 11 | 11 | 9 | 9 | 9 |
| River Raisin, MI | 42.177829 | −84.092819 | 17 | 17 | 17 | 3 | 3 | 3 |
| St Joseph River (Maumee), MI | 41.781106 | −84.650714 | 6 | 6 | 6 | 0 | 0 | 0 |
| Sydenham River, ON | 42.589146 | −82.127622 | 15 | 11 | 11 | 5 | 9 | 9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Willsie, J.A.; Morris, T.J.; Zanatta, D.T. Morphometric Analyses Distinguish Wabash Pigtoe (Fusconaia flava) and Round Pigtoe (Pleurobema sintoxia) Mussels. Diversity 2020, 12, 337. https://doi.org/10.3390/d12090337
Willsie JA, Morris TJ, Zanatta DT. Morphometric Analyses Distinguish Wabash Pigtoe (Fusconaia flava) and Round Pigtoe (Pleurobema sintoxia) Mussels. Diversity. 2020; 12(9):337. https://doi.org/10.3390/d12090337
Chicago/Turabian StyleWillsie, Julia A., Todd J. Morris, and David T. Zanatta. 2020. "Morphometric Analyses Distinguish Wabash Pigtoe (Fusconaia flava) and Round Pigtoe (Pleurobema sintoxia) Mussels" Diversity 12, no. 9: 337. https://doi.org/10.3390/d12090337
APA StyleWillsie, J. A., Morris, T. J., & Zanatta, D. T. (2020). Morphometric Analyses Distinguish Wabash Pigtoe (Fusconaia flava) and Round Pigtoe (Pleurobema sintoxia) Mussels. Diversity, 12(9), 337. https://doi.org/10.3390/d12090337