Individual Variation in Predatory Behavior, Scavenging and Seasonal Prey Availability as Potential Drivers of Coexistence between Wolves and Bears

 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Area and Study Species
2.2. GPS and Predation Data from Bears and Wolves
2.3. Monitoring of Scavenging with Camera Traps
2.4. Data Analyses
2.4.1. Kill Rates
2.4.2. Scavenging Events
3. Results
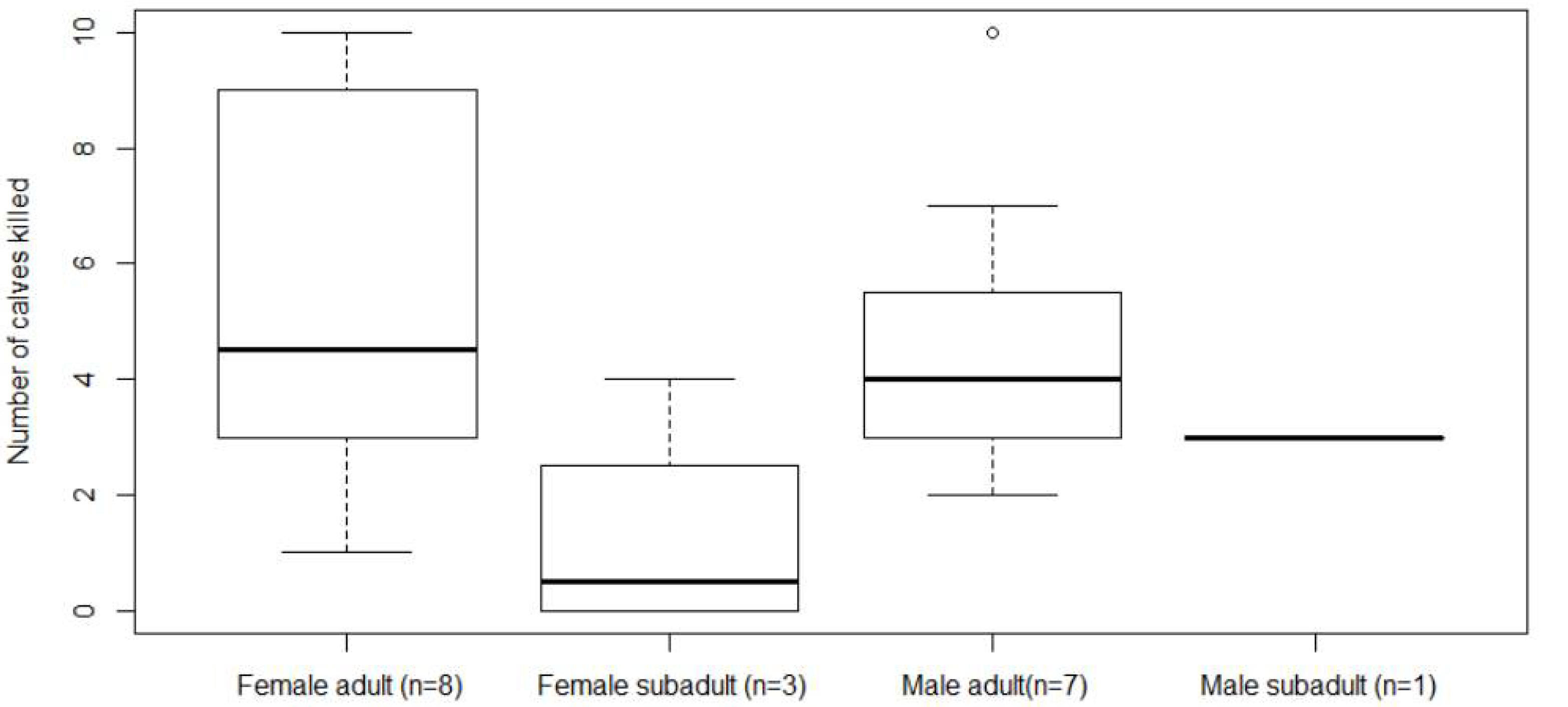
3.1. Direct Interactions Derived from GPS-Data and Field Visits
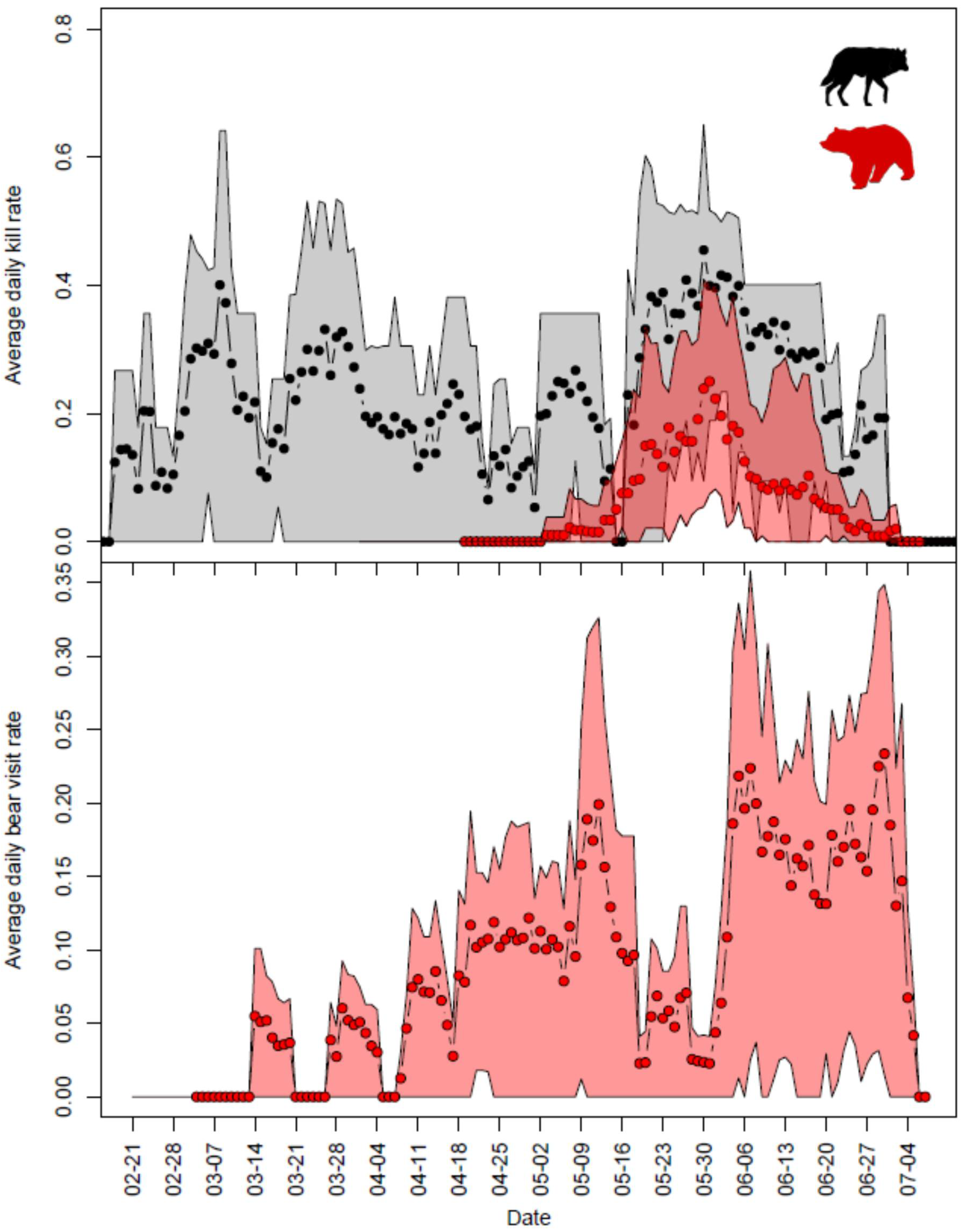
3.2. Temporal Patterns in Bear and Wolf Kill Rates
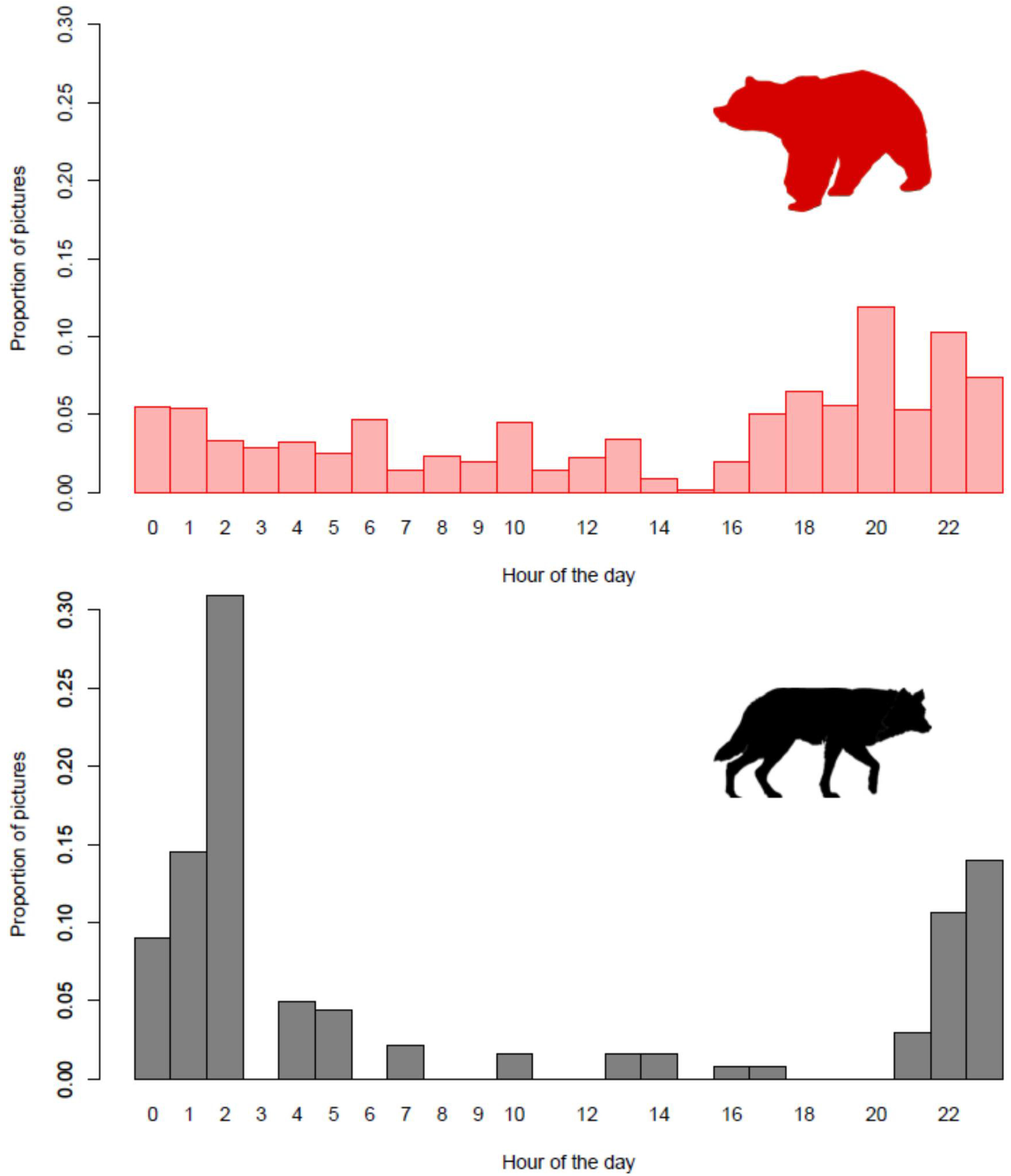
3.3. Scavenging Events
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Bear and Year of Study | Start Date | End Date |
|---|---|---|
| Lafmamack 2014 | 5/14/2014 | 5/14/2014 |
| Lafmamack 2014 | 5/22/2014 | 5/27/2014 |
| Lafmamack 2014 | 6/3/2014 | 6/9/2014 |
| Lafmamack 2014 | 6/12/2014 | 6/27/2014 |
| Galju 2014 | 4/28/2014 | 7/1/2014 |
| Klummy 2014 | 4/30/2014 | 5/9/2014 |
| Klummy 2014 | 5/21/2014 | 5/26/2014 |
| Klummy 2014 | 6/12/2014 | 6/19/2014 |
| Klummy 2014 | 6/23/2014 | 6/27/2014 |
| Klumpa 2014 | 4/30/2014 | 5/9/2014 |
| Klumpa 2014 | 5/21/2014 | 6/27/2014 |
| Risslo 2014 | 4/22/2014 | 6/23/2014 |
| Ruta 2014 | 4/29/2014 | 6/27/2014 |
| Jarpa 2014 | 5/6/2014 | 6/23/2014 |
| Kil-kalle 2014 | 4/30/2014 | 6/27/2014 |
| Spjuta 2014 | 5/11/2014 | 7/1/2014 |
| Strandas 2014 | 5/11/2014 | 6/27/2014 |
| Tappele 2014 | 5/11/2014 | 7/1/2014 |
| Abborrgina 2015 | 5/3/2015 | 7/1/2015 |
| Gymasa 2015 | 5/13/2015 | 7/3/2015 |
| Jarpa 2015 | 6/12/2015 | 7/3/2015 |
| Kil-kalle 2015 | 4/27/2015 | 5/3/2015 |
| Kil-kalle 2015 | 5/13/2015 | 5/16/2015 |
| Kil-kalle 2015 | 5/27/2015 | 5/30/2015 |
| Kil-kalle 2015 | 6/6/2015 | 6/15/2015 |
| Kil-kalle 2015 | 6/24/2015 | 7/3/2015 |
| Klummy 2015 | 4/27/2015 | 7/3/2015 |
| Klumpa 2015 | 4/27/2015 | 5/25/2015 |
| Klumpa 2015 | 6/6/2015 | 6/9/2015 |
| Klumpa 2015 | 6/21/2015 | 6/24/2015 |
| Lutane 2015 | 4/27/2015 | 5/31/2015 |
| Lutane 2015 | 6/6/2015 | 7/3/2015 |
| Risslo 2015 | 4/27/2015 | 5/22/2015 |
| Risslo 2015 | 6/9/2015 | 6/15/2015 |
| Risslo 2015 | 6/21/2015 | 7/3/2015 |
| Tappele 2015 | 4/27/2015 | 6/24/2015 |

References
- Terborgh, J.; Estes, J.A. Trophic Cascades; Island Press: Washington, DC, USA, 2010. [Google Scholar]
- Ripple, W.J.; Smith, D.W.; Estes, J.A.; Beschta, R.L.; Wilmers, C.C.; Ritchie, E.G.; Hebblewhite, M.; Berger, J.; Elmhagen, B.; Letnic, M.; et al. Status and ecological effects of the world’s largest carnivores. Science 2014, 343, 1241484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebblewhite, M.; Hurd, T.E.; White, C.A.; Fryxell, J.M.; Nietvelt, C.G.; McKenzie, J.A.; Bayley, S.E.; Paquet, P.C. Human Activity Mediates a Trophic Cascade Caused By Wolves. Ecology 2005, 86, 2135–2144. [Google Scholar] [CrossRef]
- Ordiz, A.; Bischof, R.; Swenson, J.E. Saving large carnivores, but losing the apex predator? Biol. Conserv. 2013, 168, 128–133. [Google Scholar] [CrossRef] [Green Version]
- Bruskotter, J.T.; Shelby, L.B. Human Dimensions of Large Carnivore Conservation and Management. Hum. Dimens. Wildl. 2010, 15, 311–314. [Google Scholar] [CrossRef]
- Chapron, G.; Kaczensky, P.; Linnell, J.D.C.; von Arx, M.; Huber, H.; Andrén, H. Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science 2014, 346, 1517–1519. [Google Scholar] [CrossRef] [Green Version]
- May, R.; Van Dijk, J.; Wabakken, P.; Swenson, J.E.; Linnell, J.D.C.; Odden, J.; Pedersen, H.C.; Andersen, R.; Landa, A. Habitat differentiation within the large-carnivore community of Norway’s multiple-use landscapes. J. Appl. Ecol. 2008, 45, 1382–1391. [Google Scholar] [CrossRef] [Green Version]
- Wikenros, C.; Liberg, O.; Sand, H.; Andrén, H. Competition between recolonizing wolves and resident lynx in Sweden. Can. J. Zool. 2010, 88, 271–279. [Google Scholar] [CrossRef]
- Mattisson, J.; Andrén, H.; Persson, J.; Segerström, P. Influence of intraguild interactions on resource use by wolverines and Eurasian lynx. J. Mammal. 2011, 92, 1321–1330. [Google Scholar] [CrossRef]
- Elmhagen, B.; Ludwig, G.; Rushton, S.P.; Helle, P.; Lindén, H. Top predators, mesopredators and their prey: Interference ecosystems along bioclimatic productivity gradients. J. Anim. Ecol. 2010, 79, 785–794. [Google Scholar] [CrossRef]
- Gervasi, V.; Nilsen, E.B.; Sand, H.; Panzacchi, M.; Rauset, G.R.; Pedersen, H.C.; Kindberg, J.; Wabakken, P.; Zimmermann, B.; Odden, J.; et al. Predicting the potential demographic impact of predators on their prey: A comparative analysis of two carnivore-ungulate systems in Scandinavia. J. Anim. Ecol. 2012, 81, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Ballard, W.; Carbyn, L.N.; Smith, D.W. Wolf interactions with non-prey. In Wolves: Behavior, Ecology and Conservation; Mech, L.D., Boitani, L., Eds.; University of Chicago Press: Chicago, IL, USA, 2003; pp. 259–271. [Google Scholar]
- Gunther, K.A.; Smith, D.W. Interactions between wolves and female grizzly bears with cubs in Yellowstone National Park. Ursus 2004, 15, 232–238. [Google Scholar] [CrossRef]
- Lewis, T.M.; Lafferty, D.J.R. Brown bears and wolves scavenge humpback whale carcass in Alaska. Ursus 2014, 25, 8–13. [Google Scholar] [CrossRef]
- Ordiz, A.; Krofel, M.; Milleret, C.; Seryodkin, I.V.; Tallian, A.; Støen, O.-G.; Sivertsen, T.R.; Kindberg, J.; Wabakken, P.; Sand, H.; et al. Interspecific interactions between brown bears, ungulates, and other large carnivores. In Bears of the World; Penteriani, V., Melletti, M., Eds.; Cambridge University Press: Cambridge, UK, 2020; in press. [Google Scholar]
- Ordiz, A.; Milleret, C.; Kindberg, J.; Mansson, J.; Wabakken, P.; Swenson, J.E.; Sand, H. Wolves, people, and brown bears influence the expansion of the recolonizing Wolf population in Scandinavia. Ecosphere 2015, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Milleret, C.; Ordiz, A.; Chapron, G.; Andreassen, H.P.; Kindberg, J.; Månsson, J.; Tallian, A.; Wabakken, P.; Wikenros, C.; Zimmermann, B.; et al. Habitat segregation between brown bears and gray wolves in a human-dominated landscape. Ecol. Evol. 2018, 8, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Pérez, A.; Ordiz, A.; Sand, H.; Swenson, J.E.; Wabakken, P.; Wikenros, C.; Zimmermann, B.; Akesson, M.; Milleret, C. No place like home? A test of the natal habitat-biased dispersal hypothesis in Scandinavian wolves. R. Soc. Open Sci. 2018, 5, 181379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ordiz, A.; Uzal, A.; Milleret, C.; Sanz-pérez, A.; Zimmermann, B.; Wikenros, C.; Wabakken, P.; Kindberg, J.; Swenson, J.E.; Sand, H. Wolf habitat selection when sympatric or allopatric with brown bears in Scandinavia. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Tallian, A.; Ordiz, A.; Metz, M.C.; Milleret, C.; Wikenros, C.; Smith, D.W.; Stahler, D.R.; Kindberg, J.; Macnulty, D.R.; Wabakken, P.; et al. Competition between apex predators? Brown bears decrease wolf kill rate on two continents. Proc. R. Soc. B Biol. Sci. 2017, 284, 20162368. [Google Scholar] [CrossRef]
- Boertje, R.D.; Keech, M.A.; Paragi, T.F. Science and Values Influencing Predator Control for Alaska Moose Management. J. Wildl. Manag. 2010, 74, 917–928. [Google Scholar] [CrossRef]
- Jonzén, N.; Sand, H.; Wabakken, P.; Swenson, J.E.; Kindberg, J.; Liberg, O.; Chapron, G. Sharing the bounty—Adjusting harvest to predator return in the Scandinavian human–wolf–bear–moose system. Ecol. Model. 2013, 265, 140–148. [Google Scholar] [CrossRef]
- Rauset, G.R.; Mattisson, J.; Andrén, H.; Chapron, G.; Persson, J. When species’ ranges meet: Assessing differences in habitat selection between sympatric large carnivores. Oecologia 2013, 172, 701–711. [Google Scholar] [CrossRef]
- Mattisson, J.; Sand, H.; Wabakken, P.; Gervasi, V.; Liberg, O.; Linnell, J.D.C.; Rauset, G.R.; Pedersen, H.C. Home range size variation in a recovering wolf population: Evaluating the effect of environmental, demographic, and social factors. Oecologia 2013, 173, 813–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leclerc, M.; Vander, E.; Zedrosser, A.; Swenson, J.E.; Kindberg, J.; Pelletier, F. Quantifying consistent individual differences in habitat selection. Oecologia 2016, 180, 697–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauset, G.R.; Kindberg, J.; Swenson, J.E. Modeling female brown bear kill rates on moose calves using global positioning satellite data. J. Wildl. Manag. 2012, 76, 1597–1606. [Google Scholar] [CrossRef]
- Toscano, B.J.; Gownaris, N.J.; Heerhartz, S.M. Personality, foraging behavior and specialization: Integrating behavioral and food web ecology at the individual level. Oecologia 2016, 182, 55–69. [Google Scholar] [CrossRef]
- Hertel, A.G.; Swenson, J.E.; Bischof, R. A case for considering individual variation in diel activity patterns. Behav. Ecol. 2017, 28, 1524–1531. [Google Scholar] [CrossRef]
- Di Rienzo, N.; Montiglio, P. Four ways in which data-free papers on animal personality fail to be impactful. Front. Ecol. Evol. 2015, 3, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Davis, G.H.; Payne, E.; Sih, A. Commentary: Four ways in which data-free papers on animal personality fail to be impactful. Front. Ecol. Evol. 2015, 3, 102–104. [Google Scholar] [CrossRef] [Green Version]
- Bolnick, D.I.; Amarasekare, P.; Araújo, M.S.; Bürger, R.; Levine, J.M.; Novak, M.; Rudolf, V.H.W.; Schreiber, S.J.; Mark, C. Why intraspecific trait variation matters in community ecology. Trends Ecol. Evol. 2011, 26, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Valladares, F.; Bastias, C.; Godoy, O.; Granda, E.; Escudero, A. Species coexistence in a changing world. Front. Plant Sci. 2015, 6, 1–16. [Google Scholar] [CrossRef]
- Basille, M.; Fortin, D.; Dussault, C.; Ouellet, J.P.; Courtois, R. Ecologically based definition of seasons clarifies predator-prey interactions. Ecography 2013, 36, 220–229. [Google Scholar] [CrossRef]
- Araújo, M.B.; Rozenfeld, A. The geographic scaling of biotic interactions. Ecography 2014, 37, 406–415. [Google Scholar] [CrossRef] [Green Version]
- Ordiz, A.; Kindberg, J.; Sæbø, S.; Swenson, J.E.; Støen, O.G. Brown bear circadian behavior reveals human environmental encroachment. Biol. Conserv. 2014, 173, 1–9. [Google Scholar] [CrossRef]
- Wabakken, P.; Sand, H.; Liberg, O.; Bjärvall, A. The recovery, distribution, and population dynamics of wolves on the Scandinavian peninsula, 1978–1998. Can. J. Zool. 2001, 725, 710–725. [Google Scholar] [CrossRef]
- Wabakken, P.; Svensson, L.; Maartmann, E.; Nordli, K.; Flagstad, Ø.; Åkesson, M. Bestandsovervåking av ulv vinteren 2019-2020 (In Norwegian); Rovdata and Viltskadecenter; SLU: St. Louis, MO, USA, 2020. [Google Scholar]
- Swenson, J.E.; Wabakken, P.; Sandegren, F.; Bjärvall, A.; Franzén, R.; Söderberg, A. The near extinction and recovery of brown bears in Scandinavia in relation to the bear management policies of Norway and Sweden. Wildl. Biol. 1995, 1, 11–25. [Google Scholar] [CrossRef]
- Solberg, K.H.; Bellemain, E.; Drageset, O.M.; Taberlet, P.; Swenson, J.E. An evaluation of field and non-invasive genetic methods to estimate brown bear (Ursus arctos) population size. Biol. Conserv. 2006, 128, 158–168. [Google Scholar] [CrossRef]
- Swenson, J.E.; Schneider, M.; Zedrosser, A.; Söderberg, A.; Franzén, R.; Kindberg, J. Challenges of managing a European brown bear population; lessons from Sweden, 1943–2013. Wildl. Biol. 2017, 1, wlb.00251. [Google Scholar] [CrossRef] [Green Version]
- Sand, H.; Eklund, A.; Zimmermann, B.; Wikenros, C.; Wabakken, P. Prey selection of Scandinavian wolves: Single large or several small? PLoS ONE 2016, 11, e0168062. [Google Scholar] [CrossRef]
- Arnemo, J.; Fahlman, Å.; Ahlqvist, P.; Andersen, R.; Andrén, H.; Brunberg, S.; Landa, A.; Liberg, O.; Odden, J.; Persson, J.; et al. Biomedical Protocols for Free-Ranging Brown Bears, Gray Wolves, Wolverines and Lynx; Arnemo, J., Fahlman, A., Eds.; Norwegian School of Veterinary Science: Tromsø, Norway, 2007. [Google Scholar]
- Ordiz, A.; Støen, O.-G.; Swenson, J.E.; Kojola, I.; Bischof, R. Distance-dependent effect of the nearest neighbor: Spatiotemporal patterns in brown bear reproduction. Ecology 2008, 89. [Google Scholar] [CrossRef]
- Sand, H.; Zimmermann, B.; Wabakken, P.; Andrèn, H.; Pedersen, H.C. Using GPS technology and GIS cluster analyses to estimate kill rates in wolf—ungulate ecosystems. Wildl. Soc. Bull. 2005, 33, 914–925. [Google Scholar] [CrossRef]
- Sand, H.; Wabakken, P.; Zimmermann, B.; Johansson, Ö.; Pedersen, H.C.; Liberg, O. Summer kill rates and predation pattern in a wolf-moose system: Can we rely on winter estimates? Oecologia 2008, 156, 53–64. [Google Scholar] [CrossRef]
- Stenset, N.E.; Lutnæs, P.N.; Bjarnadóttir, V.; Dahle, B.; Fossum, K.H.; Jigsved, P.; Johansen, T.; Neumann, W.; Opseth, O.; Rønning, O.; et al. Seasonal and annual variation in the diet of brown bears Ursus arctos in the boreal forest of southcentral Sweden. Wildl. Biol. 2016, 22, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Ordiz, A.; Rodríguez, C.; Naves, J.; Fernández, A.; Huber, D.; Kaczensky, P.; Mertens, A.; Mertzanis, Y.; Mustoni, A.; Palazón, S.; et al. Distance-based criteria to identify minimum number of brown bear females with cubs in Europe. Ursus 2007, 18, 158–167. [Google Scholar] [CrossRef]
- Swenson, J.; Dahle, B.; Busk, H.; Opseth, O.; Johansen, T.; Soderberg, A.; Wallin, K.; Cederlund, G. Predation on Moose Calves by European Brown Bears. J. Wildl. Manag. 2007, 71, 1993–1997. [Google Scholar] [CrossRef]
- Krofel, M.; Kos, I.; Jerina, K. The noble cats and the big bad scavengers: Effects of dominant scavengers on solitary predators. Behav. Ecol. Sociobiol. 2012, 66, 1297–1304. [Google Scholar] [CrossRef]
- Steyaert, S.M.J.G.; Kindberg, J.; Swenson, J.E.; Zedrosser, A. Male reproductive strategy explains spatiotemporal segregation in brown bears. J. Anim. Ecol. 2013, 82, 836–845. [Google Scholar] [CrossRef] [Green Version]
- López-Bao, J.V.; Mattisson, J.; Persson, J.; Aronsson, M.; Andrén, H. Tracking neighbours promotes the coexistence of large carnivores. Sci. Rep. 2016, 6, 23198. [Google Scholar] [CrossRef] [Green Version]
- Réale, D.; Reader, S.; Sol, D.; McDougall, P.; Dingemanse, N. Integrating animal temperament within ecology and evolution. Biol. Rev. Camb. Philos. Soc. 2007, 82, 291–318. [Google Scholar] [CrossRef] [Green Version]
- Bergman, J.N.; Bennett, J.R.; Binley, A.D.; Cooke, S.J.; Fyson, V.; Hlina, B.L.; Reid, C.H.; Vala, M.A.; Madliger, C.L. Scaling from individual physiological measures to population-level demographic change: Case studies and future directions for conservation management. Biol. Conserv. 2019, 238, 108242. [Google Scholar] [CrossRef]
- Cozzi, G.; Chynoweth, M.; Kusak, J.; Coban, E.; Coban, A.; Ozgul, A.; Sekerciouglu, C.H. Anthropogenic food resources foster the coexistence of distinct life history strategies: Year-round sedentary and migratory brown bears. J. Zool. 2016, 300, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Estes, J.; Riedman, M.; Staedler, M.; Tinker, M.; Lyon, B. Individual variation in prey selection by sea otters: Patterns, causes and implications. J. Anim. Ecol. 2003, 72, 144–155. [Google Scholar] [CrossRef] [Green Version]
- Støen, O.-G.; Bellemain, E.; Solve, S.; Swenson, J. Kin-related spatial structure in brown bears Ursus arctos. Behav. Ecol. 2005, 59, 191–197. [Google Scholar] [CrossRef]
- Suraci, J.P.; Nickel, B.A.; Wilmers, C.C. Fine-scale movement decisions by a large carnivore inform conservation planning in human-dominated landscapes. Landsc. Ecol. 2020, 35, 1635–1649. [Google Scholar] [CrossRef]
- Palomares, F.; Caro, T.M. Interspecific Killing among Mammalian Carnivores. Am. Nat. 1999, 153, 492–508. [Google Scholar] [CrossRef] [PubMed]
- Caro, T.M.; Stoner, C.J. The potential for interspecific competition among African carnivores. Biol. Conserv. 2003, 110, 67–75. [Google Scholar] [CrossRef]
- Périquet, S.; Fritz, H.; Revilla, E. The lion king and the hyaena queen: Large carnivore interactions and coexistence. Biol. Rev. 2015, 90, 1197–1214. [Google Scholar] [CrossRef]
- Wilmers, C.; Post, E. Predicting the influence of wolf-provided carrion on scavenger community dynamics under climate change scenarios. Glob. Chang. Biol. 2006, 12, 403–409. [Google Scholar] [CrossRef]
- Dettki, H.; Ericsson, G.; Giles, T.; Norrsken-Ericsson, M. Wireless Remote Animal Monitoring (WRAM)—A new international database e-infrastructure for telemetry sensor data from fish and wildlife. In Proceedings Etc 2012: Convention for Telemetry, Test Instrumentation and Telecontrol; European Society of Telemetry; Books on Demand: Manila, Philippines, 2013; pp. 247–256. ISBN 978-3-7322-5646-4. [Google Scholar]


| Bear | Year | Predation Period | Predation Period (# Days) | # Killed Calves | Bear Sex and Age Class | |
|---|---|---|---|---|---|---|
| Galju | 2014 | 16.05.2014 | 17.06.2014 | 32 | 10 | Single adult female |
| Ruta | 2014 | 18.05.2014 | 18.06.2014 | 31 | 5 | Single adult female |
| Spjuta | 2014 | 21.05.2014 | 12.06.2014 | 22 | 10 | Single adult female |
| Strandas | 2014 | 03.06.2014 | 09.06.2014 | 6 | 2 | Single adult female |
| Klummy | 2014 | 15.06.2014 | 15.06.2014 | 1 | 1 | Subadult female |
| 2015 | 08.05.2015 | 18.06.2015 | 41 | 4 | Single adult female | |
| Klumpa | 2014 | 10.06.2014 | 25.06.2014 | 15 | 4 | Subadult female |
| 2015 | 06.05.2015 | 19.05.2015 | 13 | 1 | Single adult female | |
| Jarpa | 2014 | _ | _ | 0 | 0 | Subadult female |
| 2015 | _ | _ | 0 | 0 | Subadult female | |
| Lafmamack | 2014 | 23.05.2014 | 24.06.2014 | 32 | 3 | Adult male |
| Risslo | 2014 | 13.05.2014 | 06.06.2014 | 24 | 7 | Adult male |
| 2015 | 20.05.2015 | 21.06.2015 | 32 | 4 | Adult male | |
| Tappele | 2014 | 22.05.2014 | 01.06.2014 | 10 | 10 | Adult male |
| 2015 | 11.05.2015 | 17.06.2015 | 37 | 4 | Adult male | |
| Kil-kalle | 2014 | 28.05.2014 | 09.06.2014 | 12 | 3 | Subadult male |
| 2015 | 28.05.2015 | 10.06.2015 | 13 | 3 | Adult male | |
| Lutane | 2015 | 18.05.2015 | 21.06.2015 | 34 | 2 | Adult male |
| Abborrgina | 2015 | 23.05.2015 | 19.06.2015 | 27 | 8 | Adult female + 2 year-old cubs |
| Gymasa | 2015 | 18.05.2015 | 20.06.2015 | 33 | 4 | Single adult female |
| Wolf Territory | Year | Start Date | End Date | Killed prey (# per Moose Age Class) |
|---|---|---|---|---|
| Tenskog | 2010 | 2/13/2010 | 4/11/2010 | 5 juveniles |
| 2011 | 3/14/2011 | 5/16/2011 | 8 juveniles | |
| Tandsjön | 2012 | 2/20/2012 | 5/14/2012 | 1 neonate calf, 5 juveniles, 7 adults, 1 unknown (juvenile or adult) |
| 2014a | 3/19/2014 | 4/28/2014 | 3 juveniles | |
| 2014b | 5/21/2014 | 6/21/2014 | 11 neonate calves, 3 juveniles | |
| Kukumäki | 2013 | 2/25/2013 | 4/28/2013 | 2 juveniles, 2 adults |
| 2014a | 3/3/2014 | 4/27/2014 | 3 juveniles | |
| 2014b | 5/19/2014 | 6/22/2014 | 7 neonate calves, 6 juveniles | |
| 2015a | 3/4/2015 | 4/24/2015 | 11 juveniles, 1 unknown (juvenile or adult) | |
| 2015b | 5/18/2015 | 6/29/2015 | 16 neonate calves, 2 juveniles, 1 adult |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ordiz, A.; Milleret, C.; Uzal, A.; Zimmermann, B.; Wabakken, P.; Wikenros, C.; Sand, H.; Swenson, J.E.; Kindberg, J. Individual Variation in Predatory Behavior, Scavenging and Seasonal Prey Availability as Potential Drivers of Coexistence between Wolves and Bears. Diversity 2020, 12, 356. https://doi.org/10.3390/d12090356
Ordiz A, Milleret C, Uzal A, Zimmermann B, Wabakken P, Wikenros C, Sand H, Swenson JE, Kindberg J. Individual Variation in Predatory Behavior, Scavenging and Seasonal Prey Availability as Potential Drivers of Coexistence between Wolves and Bears. Diversity. 2020; 12(9):356. https://doi.org/10.3390/d12090356
Chicago/Turabian StyleOrdiz, Andrés, Cyril Milleret, Antonio Uzal, Barbara Zimmermann, Petter Wabakken, Camilla Wikenros, Håkan Sand, Jon E Swenson, and Jonas Kindberg. 2020. "Individual Variation in Predatory Behavior, Scavenging and Seasonal Prey Availability as Potential Drivers of Coexistence between Wolves and Bears" Diversity 12, no. 9: 356. https://doi.org/10.3390/d12090356
APA StyleOrdiz, A., Milleret, C., Uzal, A., Zimmermann, B., Wabakken, P., Wikenros, C., Sand, H., Swenson, J. E., & Kindberg, J. (2020). Individual Variation in Predatory Behavior, Scavenging and Seasonal Prey Availability as Potential Drivers of Coexistence between Wolves and Bears. Diversity, 12(9), 356. https://doi.org/10.3390/d12090356