Altitudinal Vascular Plant Richness and Climate Change in the Alpine Zone of the Lefka Ori, Crete

 , ,
, ,  and
and
Abstract
:1. Introduction
- What are the changes in sub-surface temperature?
- What are the changes in species richness, species, and vegetation turnover?
- Are changes scale specific?
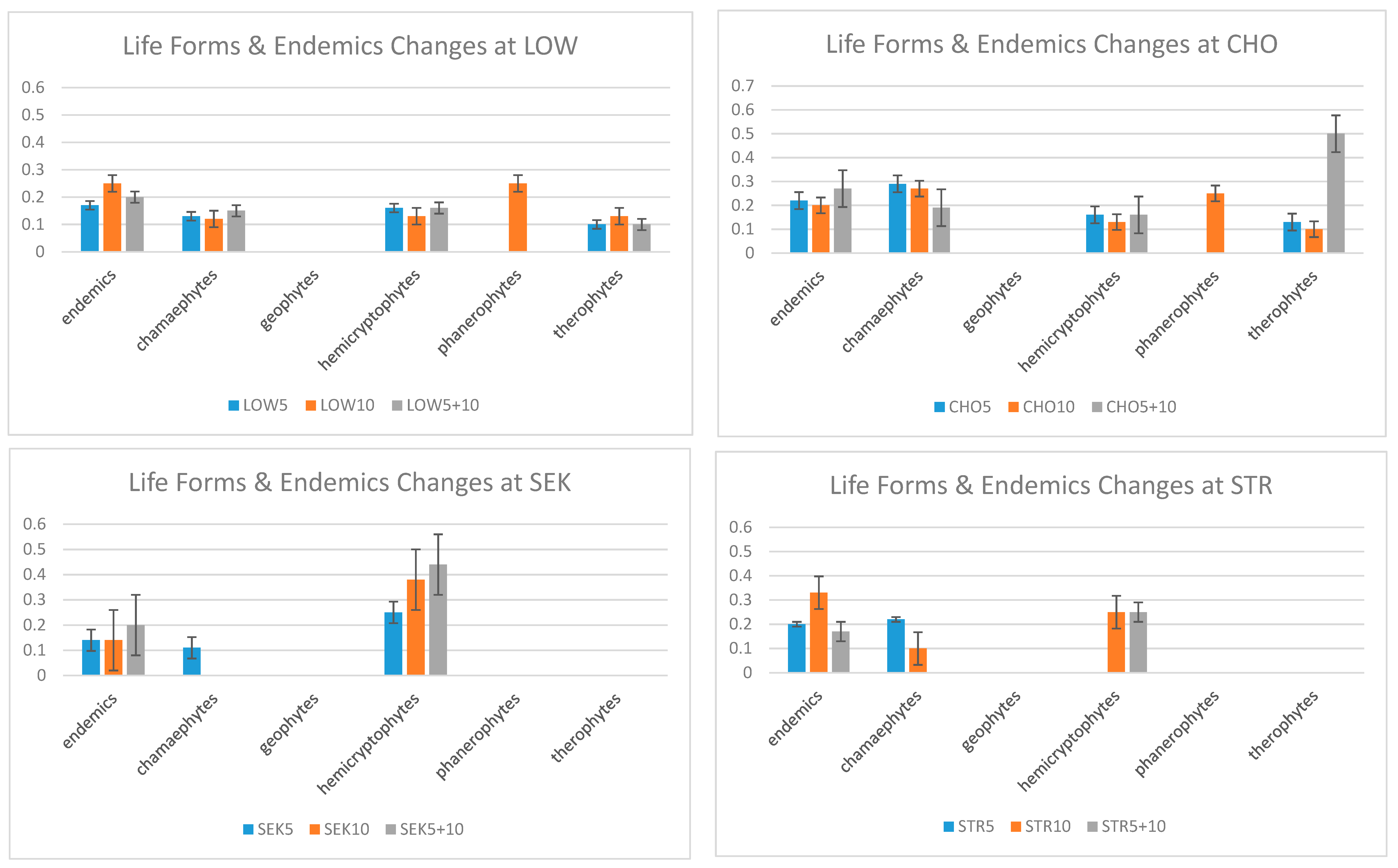
- Do different life forms and endemics show strong changes over time?
- What are the effects of temperature on species richness?
- Are observed changes fluctuations or signals of climate change?
2. Materials and Methods
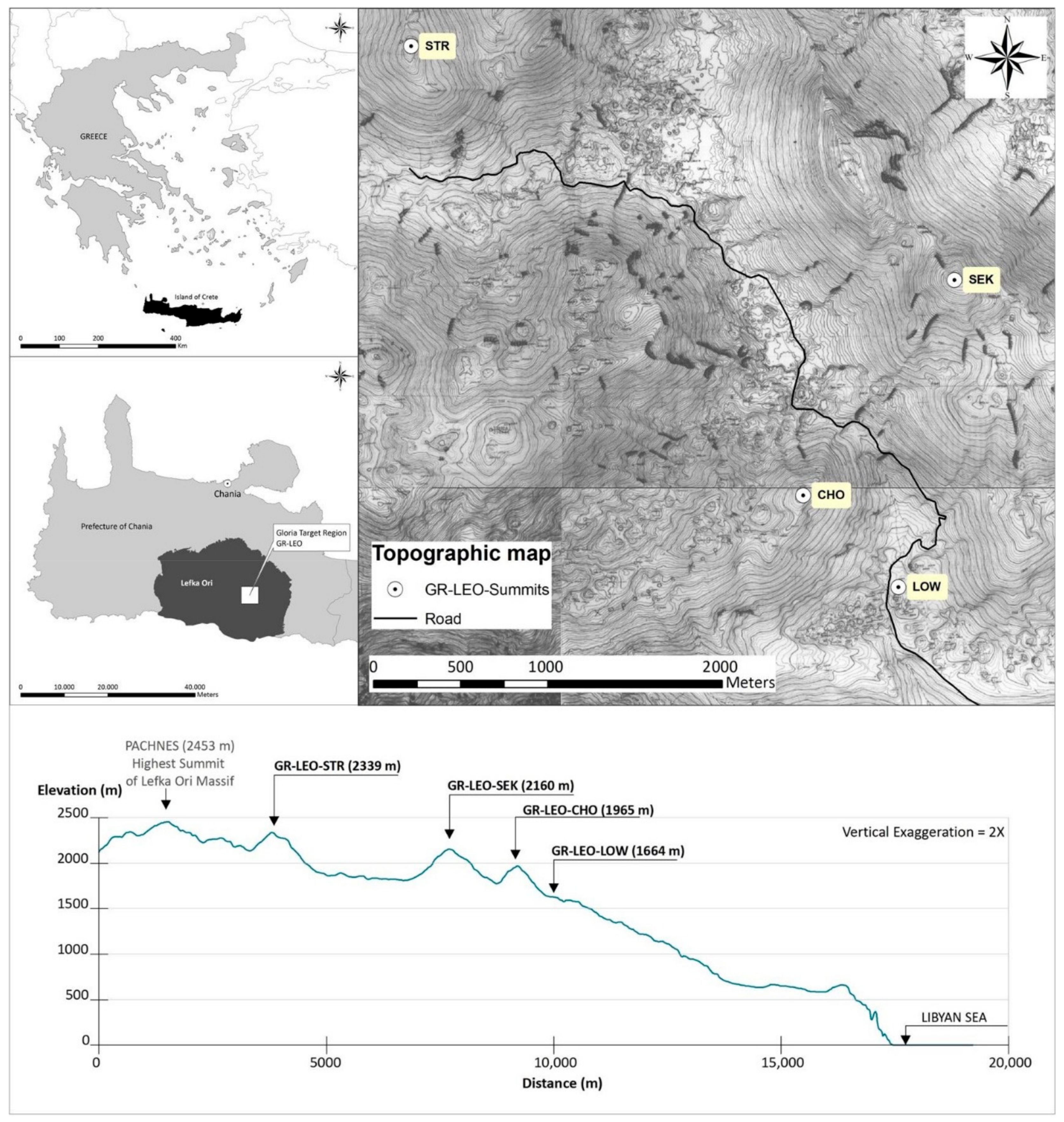
2.1. Study Area
2.2. Vegetation Sampling
2.3. Climatic Variation
2.4. Data Analysis
2.4.1. Climate
2.4.2. Species Richness
2.4.3. Species Diversity and Turnover
- Species turnover based on individual species frequency:where A is the frequency of quadrats where the species appeared in 2008; D is the frequency of quadrats where the species disappeared; U is the frequency of quadrats where the species’ frequency was unchanged.Tsp = (A + D)/(A + D + U),
- Vegetation turnover index at different spatial scales and life forms (at summit level):where A is the number of new species in 2008; D is the number of disappearing species; B is the number of species present in both years of comparison.Tveg = (A + D)/(A + D + B)
3. Results
3.1. Climate
3.2. General Patterns
3.3. Species and Vegetation Turnover
3.4. Species Richness and Climate
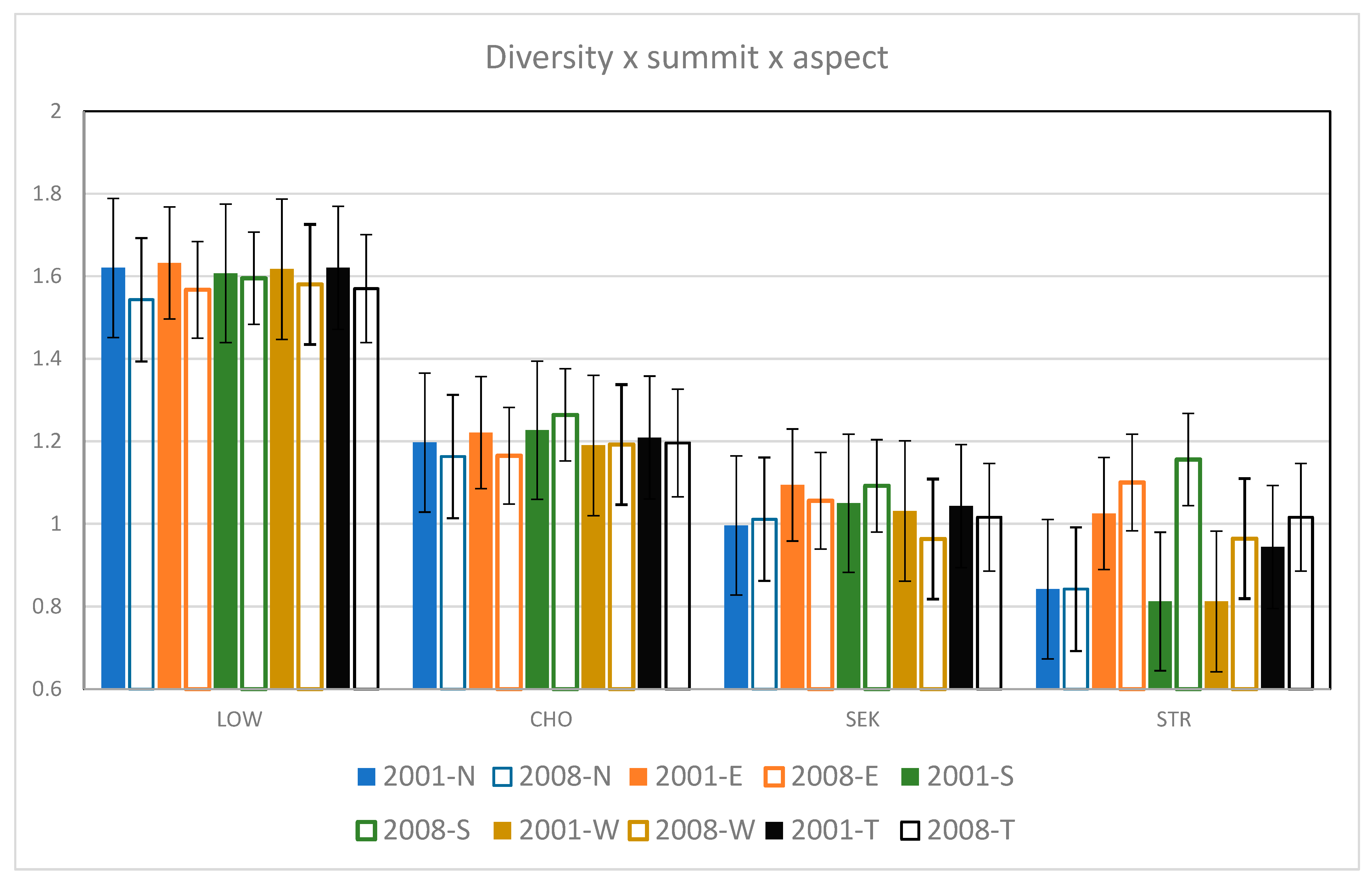
3.5. Species Diversity
4. Discussion
4.1. Species Richness and Climatic Parameters
4.2. Changes in Species Composition and Abundance
4.3. Changes in Endemic Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malcolm, J.R.; Liu, C.; Neilson, R.P.; Hansen, L.; Hannah, L. Global Warming and Extinctions of Endemic Species from Biodiversity Hotspots. Conserv. Biol. 2006, 20, 538–548. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction Risk from Climate Change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, J.J.; Intergovernmental Panel on Climate Change (Eds.) Climate Change 2001: Impacts, Adaptation, and Vulnerability: Contribution of Working Group II to the Third Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2001; ISBN 978-0-521-80768-5. [Google Scholar]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. (Eds.) IPCC, 2013: Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; p. 1535. [Google Scholar]
- Hock, R.; Rasul, G.; Adler, C.; Cáceres, B.; Gruber, S.; Hirabayashi, Y.; Jackson, M.; Kääb, A.; Kang, S.; Kutuzov, S.; et al. High Mountain Areas; IPCC—Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2019; pp. 131–202. [Google Scholar]
- Körner, C. Alpine Plant. Life: Functional Plant. Ecology of High. Mountain Ecosystems, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2003; ISBN 978-3-540-00347-2. [Google Scholar]
- Pauli, H.; Gottfried, M.; Reiter, K.; Klettner, C.; Grabherr, G. Signals of Range Expansions and Contractions of Vascular Plants in the High Alps: Observations (1994–2004) at the GLORIA Master Site Schrankogel, Tyrol, Austria. Glob. Chang. Biol. 2007, 13, 147–156. [Google Scholar] [CrossRef]
- Pauli, H.; European Commission; Directorate General for Research. The Gloria Field Manual: Multi-Summit Approach; Directorate-General for Research: Luxembourg, 2004; ISBN 978-92-894-4737-9. [Google Scholar]
- Holzinger, B.; Hülber, K.; Camenisch, M.; Grabherr, G. Changes in Plant Species Richness over the Last Century in the Eastern Swiss Alps: Elevational Gradient, Bedrock Effects and Migration Rates. Plant Ecol. 2008, 195, 179–196. [Google Scholar] [CrossRef]
- Vittoz, P.; Bodin, J.; Ungricht, S.; Burga, C.A.; Walther, G.-R. One Century of Vegetation Change on Isla Persa, a Nunatak in the Bernina Massif in the Swiss Alps. J. Veg. Sci. 2008, 19, 671–680. [Google Scholar] [CrossRef]
- Erschbamer, B.; Kiebacher, T.; Mallaun, M.; Unterluggauer, P. Short-Term Signals of Climate Change along an Altitudinal Gradient in the South Alps. Plant Ecol. 2009, 202, 79–89. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Reiter, K.; Grabherr, G. High Mountain Summits as Sensitive Indicators of Climate Change Effects on Vegetation Patterns: The “Multi Summit-Approach” of GLORIA (Global Observation Research Initiative in Alpine Environments). In Global Change and Protected Areas; Visconti, G., Beniston, M., Iannorelli, E.D., Barba, D., Eds.; Advances in Global Change Research; Springer: Dordrecht, The Netherlands, 2001; Volume 9, pp. 45–51. ISBN 978-90-481-5686-3. [Google Scholar]
- Virtanen, R.; Dirnböck, T.; Dullinger, S.; Grabherr, G.; Pauli, H.; Staudinger, M.; Villar, L. Patterns in the Plant Species Richness of European High Mountain Vegetation. In Alpine Biodiversity in Europe; Nagy, L., Grabherr, G., Körner, C., Thompson, D.B.A., Eds.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2003; Volume 167, pp. 149–172. ISBN 978-3-642-62387-5. [Google Scholar]
- Parolo, G.; Rossi, G. Upward Migration of Vascular Plants Following a Climate Warming Trend in the Alps. Basic Appl. Ecol. 2008, 9, 100–107. [Google Scholar] [CrossRef]
- Michelsen, O.; Syverhuset, A.O.; Pedersen, B.; Holten, J.I. The Impact of Climate Change on Recent Vegetation Changes on Dovrefjell, Norway. Diversity 2011, 3, 91–111. [Google Scholar] [CrossRef] [Green Version]
- Dullinger, S.; Gattringer, A.; Thuiller, W.; Moser, D.E.; Zimmermann, N.; Guisan, A.; Willner, W.; Plutzar, C.; Leitner, M.; Mang, T.; et al. Extinction debt of high-mountain plants under twenty-first-century climate change. Nat. Clim. Chang. 2012, 2, 619–622. [Google Scholar] [CrossRef]
- Grabherr, G.; Gottfried, M.; Gruber, A.; Pauli, H. Patterns and Current Changes in Alpine Plant Diversity. Ecol. Stud. Anal. Synth. (USA) 1995, 113, 167–181. [Google Scholar]
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.-P.; Iglesias, A.; Lange, M.A.; Lionello, P.; Llasat, M.C.; Paz, S.; et al. Climate Change and Interconnected Risks to Sustainable Development in the Mediterranean. Nat. Clim. Chang. 2018, 8, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Fronzek, S.; Carter, T.R.; Jylhä, K. Representing Two Centuries of Past and Future Climate for Assessing Risks to Biodiversity in Europe. Glob. Ecol. Biogeogr. 2012, 21, 19–35. [Google Scholar] [CrossRef]
- Kirtman, B.; Power, S.B.; Adedoyin, A.J.; Boer, G.J.; Bojariu, R.; Doblas-Reyes, F.; Fiore, A.M.; Kimoto, M.; Meehl, G.; Prather, M.; et al. Near-Term Climate Change: Projections and Predictability. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; p. 76. [Google Scholar]
- Polade, S.D.; Pierce, D.W.; Cayan, D.R.; Gershunov, A.; Dettinger, M.D. The Key Role of Dry Days in Changing Regional Climate and Precipitation Regimes. Sci. Rep. 2015, 4, 4364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogués-Bravo, D.; Araújo, M.B.; Lasanta, T.; Moreno, J.I.L. Climate Change in Mediterranean Mountains during the 21st Century. Ambi 2008, 37, 280–285. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Araujo, M.B.; Sykes, M.T.; Prentice, I.C. Climate Change Threats to Plant Diversity in Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar] [CrossRef] [Green Version]
- Mooney, H.A.; Kalin Arroyo, M.T.; Bond, W.J.; Canadell, J.; Hobbs, R.J.; Lavorel, S.; Neilson, R.P. Mediterranean-Climate Ecosystems. In Global Biodiversity in a Changing Environment: Scenarios for the 21st Century; Chapin, F.S., Sala, O.E., Huber-Sannwald, E., Eds.; Ecological Studies; Springer: New York, NY, USA, 2001; pp. 157–199. ISBN 978-1-4613-0157-8. [Google Scholar]
- Ruiz-Labourdette, D.; Nogués-Bravo, D.; Ollero, H.S.; Schmitz, M.F.; Pineda, F.D. Forest Composition in Mediterranean Mountains Is Projected to Shift along the Entire Elevational Gradient under Climate Change: Forest Dynamics under Climate Change. J. Biogeogr. 2012, 39, 162–176. [Google Scholar] [CrossRef]
- Kapralov, D.S.; Shiyatov, S.G.; Moiseev, P.A.; Fomin, V.V. Changes in the Composition, Structure, and Altitudinal Distribution of Low Forests at the Upper Limit of Their Growth in the Northern Ural Mountains. Russ. J. Ecol. 2006, 37, 367–372. [Google Scholar] [CrossRef]
- Kullman, L. Tree Line Population Monitoring of Pinus Sylvestris in the Swedish Scandes, 1973?2005: Implications for Tree Line Theory and Climate Change Ecology. J. Ecol. 2007, 95, 41–52. [Google Scholar] [CrossRef]
- Lenoir, J.; Gegout, J.C.; Marquet, P.A.; de Ruffray, P.; Brisse, H. A Significant Upward Shift in Plant Species Optimum Elevation During the 20th Century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef]
- Engler, R.; Randin, C.F.; Thuiller, W.; Dullinger, S.; Zimmermann, N.E.; Araújo, M.B.; Pearman, P.B.; Lay, G.L.; Piedallu, C.; Albert, C.H.; et al. 21st Century Climate Change Threatens Mountain Flora Unequally across Europe. Glob. Chang. Biol. 2011, 17, 2330–2341. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Grytnes, J.-A.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Rixen, C.; Winkler, M.; Bardy-Durchhalter, M.; Barni, E.; et al. Accelerated Increase in Plant Species Richness on Mountain Summits Is Linked to Warming. Nature 2018, 556, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Rumpf, S.B.; Hülber, K.; Klonner, G.; Moser, D.; Schütz, M.; Wessely, J.; Willner, W.; Zimmermann, N.E.; Dullinger, S. Range Dynamics of Mountain Plants Decrease with Elevation. Proc. Natl. Acad. Sci. USA 2018, 115, 1848–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, R.F.; et al. Recent Plant Diversity Changes on Europe’s Mountain Summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peñuelas, J.; Boada, M. A Global Change-Induced Biome Shift in the Montseny Mountains (NE Spain). Glob. Chang. Biol. 2003, 9, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Stanisci, A.; Pelino, G.; Blasi, C. Vascular Plant Diversity and Climate Change in the Alpine Belt of the Central Apennines (Italy). Biodivers. Conserv. 2005, 14, 1301–1318. [Google Scholar] [CrossRef]
- Kazakis, G.; Ghosn, D.; Vogiatzakis, I.N.; Papanastasis, V.P. Vascular Plant Diversity and Climate Change in the Alpine Zone of the Lefka Ori, Crete. Biodivers. Conserv. 2007, 16, 1603–1615. [Google Scholar] [CrossRef]
- Stanisci, A.; Frate, L.; Morra Di Cella, U.; Pelino, G.; Petey, M.; Siniscalco, C.; Carranza, M.L. Short-Term Signals of Climate Change in Italian Summit Vegetation: Observations at Two GLORIA Sites. Plant Biosyst. An. Int. J. Deal. All Asp. Plant Biol. 2016, 150, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Calzado, M.R.F.; Mesa, J.M.; Merzouki, A.; Porcel, M.C. Vascular Plant Diversity and Climate Change in the Upper Zone of Sierra Nevada, Spain. Plant Biosyst. An. Int. J. Deal. All Asp. Plant Biol. 2012, 146, 1044–1053. [Google Scholar] [CrossRef]
- Medail, F.; Quezel, P. Biodiversity Hotspots in the Mediterranean Basin: Setting Global Conservation Priorities. Conserv. Biol. 1999, 13, 1510–1513. [Google Scholar] [CrossRef]
- Davis, S.D.; Heywood, V.H.; Hamilton, A.C.; World Wide Fund for Nature; International Union for Conservation of Nature and Natural Resources (Eds.) Centres of Plant Diversity: A Guide and Strategy for Their Conservation; World Wide Fund for Nature (WWF) and ICUN-World Conservation Union: Cambridge, UK, 1994; ISBN 978-2-8317-0197-4. [Google Scholar]
- Vogiatzakis, I.N.; Mannion, A.M.; Sarris, D. Mediterranean Island Biodiversity and Climate Change: The Last 10,000 Years and the Future. Biodivers. Conserv. 2016, 25, 2597–2627. [Google Scholar] [CrossRef]
- Médail, F. The Specific Vulnerability of Plant Biodiversity and Vegetation on Mediterranean Islands in the Face of Global Change. Reg. Environ. Chang. 2017, 17, 1775–1790. [Google Scholar] [CrossRef] [Green Version]
- Strid, A. Phytogeographical Aspects of the Greek Mountain Flora. Fragm. Florist. Et Geobot. Suppl. 1993, 2, 411–433. [Google Scholar]
- Phitos, D. (Ed.) The Red Data Book of Rare and Threatened Plants of Greece; 1. Publ.; World Wide Fund for Nature: Athen, Greece, 1995; ISBN 978-960-7506-04-7. [Google Scholar]
- Quézel, P. The study of groupings in the countries surrounding the Mediterranean: Some methodological aspects. In Mediterranean-Type Shrublands; Di Castri, F., Goodall, D.W., Sprecht, R.L., Eds.; Elsevier: Amsterdam, The Netherlands, 1981; pp. 87–93. [Google Scholar]
- Quézel, P. Floristic composition and phytosociological structure of sclerophyllous mattoral around the Mediterranean. In Mediterranean-Type Shrublands; Di Castri, F., Goodall, D.W., Sprecht, R.L., Eds.; Elsevier: Amsterdam, The Netherlands, 1981; pp. 107–121. [Google Scholar]
- Zafran, J. Contributions à La Flore et à Vegétation de La Crète; Université de Provence: Aix en Provence, France, 1990. [Google Scholar]
- Coldea, G.; Pop, A. Floristic Diversity in Relation to Geomorphological and Climatic Factors in the Subalpinealpine Belt of the Rodna Mountains (the Romanian Carpathians). Pirineos 2004, 158–159, 61–72. [Google Scholar] [CrossRef] [Green Version]
- Mark, A.F.; Dickinson, K.J.M.; Maegli, T.; Halloy, S.R.P. Two GLORIA Long-term Alpine Monitoring Sites Established in New Zealand as Part of a Global Network. J. R. Soc. N. Z. 2006, 36, 111–128. [Google Scholar] [CrossRef]
- Pickering, C.; Hill, W.; Green, K. Vascular Plant Diversity and Climate Change in the Alpine Zone of the Snowy Mountains, Australia. Biodivers. Conserv. 2008, 17, 1627–1644. [Google Scholar] [CrossRef] [Green Version]
- Rackham, O.; Moody, J.A. The Making of the Cretan Landscape; Manchester University Press, Distributed Exclusively in the USA and Canada by St. Martin’s Press: Manchester, UK; New York, NY, USA, 1996; ISBN 978-0-7190-3646-0. [Google Scholar]
- Vogiatzakis, I.N.; Griffiths, G.H.; Mannion, A.M. Environmental Factors and Vegetation Composition, Lefka Ori Massif, Crete, S. Aegean. Environ. Factors Veg. Compos. Crete. Glob Ecol. Biogeogr. 2003, 12, 131–146. [Google Scholar] [CrossRef]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea Volumes 1–5; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1964. [Google Scholar]
- Strid, A.; Tan, K. (Eds.) Mountain Flora of Greece; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1986; ISBN 978-0-521-25737-4. [Google Scholar]
- Turland, N.J.; Chilton, J.R.; Press, J.R. Flora of the Cretan Area: Annotated Checklist and Atlas; HMSO Publications: London, UK, 1993. [Google Scholar]
- Turland, N.; Chilton, L. Flora of Crete: Supplement II, Additions 1997–2008; Marengo Publications: Retford, UK, 2008. [Google Scholar]
- Jahn, R.; Schönfelder, P. Exkursionsflora Für Kreta; Ulmer: Stuttgart, Germany, 1995. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria; Available online: https://www.r-project.org/ (accessed on 21 December 2020).
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Venn, S.; Pickering, C.; Green, K. Short-Term Variation in Species Richness across an Altitudinal Gradient of Alpine Summits. Biodivers. Conserv. 2012, 21, 3157–3186. [Google Scholar] [CrossRef]
- Parmesan, C.; Hanley, M.E. Plants and Climate Change: Complexities and Surprises. Ann. Bot. 2015, 116, 849–864. [Google Scholar] [CrossRef]
- Rahbek, C. The Elevational Gradient of Species Richness: A Uniform Pattern? Ecography 1995, 18, 200–205. [Google Scholar] [CrossRef]
- Lomolino, M.V. Elevation Gradients of Species-Density: Historical and Prospective Views. Glob. Ecol. Biogeogr. 2001, 10, 3–13. [Google Scholar] [CrossRef]
- Korner, C.; Spehn, E.M.; Spehn, E.M. Mountain Biodiversity: A Global Assessment; Routledge: Abingdon, UK, 2019; ISBN 978-0-429-34258-5. [Google Scholar]
- Gottfried, M.; Pauli, H.; Futschik, A.; Akhalkatsi, M.; Barančok, P.; Benito Alonso, J.L.; Coldea, G.; Dick, J.; Erschbamer, B.; Fernández Calzado, M.R.; et al. Continent-Wide Response of Mountain Vegetation to Climate Change. Nat. Clim. Chang. 2012, 2, 111–115. [Google Scholar] [CrossRef]
- Cavieres, L.A.; Brooker, R.W.; Butterfield, B.J.; Cook, B.J.; Kikvidze, Z.; Lortie, C.J.; Michalet, R.; Pugnaire, F.I.; Schöb, C.; Xiao, S.; et al. Facilitative Plant Interactions and Climate Simultaneously Drive Alpine Plant Diversity. Ecol. Lett. 2014, 17, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Tsanis, I.K.; Koutroulis, A.G.; Daliakopoulos, I.N.; Jacob, D. Severe Climate-Induced Water Shortage and Extremes in Crete. Clim. Chang. 2011, 106, 667–677. [Google Scholar] [CrossRef]
- Malanson, G.P.; Resler, L.M.; Butler, D.R.; Fagre, D.B. Mountain Plant Communities: Uncertain Sentinels? Prog. Phys. Geogr. Earth Environ. 2019, 43, 521–543. [Google Scholar] [CrossRef]
- Essl, F.; Dullinger, S.; Rabitsch, W.; Hulme, P.E.; Pyšek, P.; Wilson, J.R.U.; Richardson, D.M. Delayed Biodiversity Change: No Time to Waste. Trends Ecol. Evol. 2015, 30, 375–378. [Google Scholar] [CrossRef]
- Petitpierre, B.; McDougall, K.; Seipel, T.; Broennimann, O.; Guisan, A.; Kueffer, C. Will Climate Change Increase the Risk of Plant Invasions into Mountains? Ecol. Appl. 2016, 26, 530–544. [Google Scholar] [CrossRef]
- Alexander, J.M.; Diez, J.M.; Levine, J.M. Novel Competitors Shape Species’ Responses to Climate Change. Nature 2015, 525, 515–518. [Google Scholar] [CrossRef]
- le Roux, P.C.; McGeoch, M.A. Spatial Variation in Plant Interactions across a Severity Gradient in the Sub-Antarctic. Oecologia 2008, 155, 831–844. [Google Scholar] [CrossRef]
- Winkler, M.; Lamprecht, A.; Steinbauer, K.; Hülber, K.; Theurillat, J.-P.; Breiner, F.; Choler, P.; Ertl, S.; Gutiérrez Girón, A.; Rossi, G.; et al. The Rich Sides of Mountain Summits—A Pan-European View on Aspect Preferences of Alpine Plants. J. Biogeogr. 2016, 43, 2261–2273. [Google Scholar] [CrossRef]
- Dirnböck, T.; Essl, F.; Rabitsch, W. Disproportional Risk for Habitat Loss of High-Altitude Endemic Species under Climate Change. Glob. Chang. Biol. 2011, 17, 990–996. [Google Scholar] [CrossRef]
- Grabherr, G.; Gottfried, M.; Pauli, H. GLORIA: A Global Observation Research Initiative in Alpine Environments. Mt. Res. Dev. 2000, 20, 190–191. [Google Scholar] [CrossRef]
- Greuter, W. The relict element of the flora of Crete and its evolutionary significance. In Taxonomy, Phytogeography and Evolution; Academic Press: London, UK, 1972. [Google Scholar]
- Theurillat, J. Climate change and the alpine flora: Some perspectives. In Potential Ecological Impacts of Climate Change in the Alps and Fennoscandian Mountains; Jardin Botanique: Geneve, Switzerland, 1995; pp. 121–127. [Google Scholar]
- Fernández Calzado, M.R.; Molero, J. Changes in the Summit Flora of a Mediterranean Mountain (Sierra Nevada, Spain) as a Possible Effect of Climate Change. LAZA 2013, 34, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Harrison, S.; Noss, R. Endemism Hotspots Are Linked to Stable Climatic Refugia. Ann. Bot. 2017, 119, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Vogiatzakis, I.N. (Ed.) Mediterranean Mountain Environments: Vogiatzakis/Mediterranean Mountain Environments; John Wiley & Sons, Ltd.: Chichester, UK, 2012; ISBN 978-1-119-94115-6. [Google Scholar]
- Médail, F.; Diadema, K. Glacial Refugia Influence Plant Diversity Patterns in the Mediterranean Basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Hughes, P.D.; Woodward, J.C. Quaternary Glaciation in the Mediterranean Mountains: A New Synthesis. Geol. Soc. Lond. Spec. Publ. 2017, 433, 1–23. [Google Scholar] [CrossRef]
- Nagy, L.; Grabherr, G. The Biology of Alpine Habitats; OUP Oxford: Oxford, UK, 2009; ISBN 978-0-19-856703-5. [Google Scholar]
- Scherrer, D.; Körner, C. Topographically Controlled Thermal-Habitat Differentiation Buffers Alpine Plant Diversity against Climate Warming. J. Biogeogr. 2011, 38, 406–416. [Google Scholar] [CrossRef]
- Kulonen, A.; Imboden, R.A.; Rixen, C.; Maier, S.B.; Wipf, S. Enough Space in a Warmer World? Microhabitat Diversity and Small-Scale Distribution of Alpine Plants on Mountain Summits. Divers. Distrib. 2018, 24, 252–261. [Google Scholar] [CrossRef] [Green Version]
- Munson, S.M.; Sher, A.A. Long-Term Shifts in the Phenology of Rare and Endemic Rocky Mountain Plants. Am. J. Bot. 2015, 102, 1268–1276. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.D. Climate, Climate Change and Range Boundaries. Divers. Distrib. 2010, 16, 488–495. [Google Scholar] [CrossRef]
- Gómez, J.M.; González-Megías, A.; Lorite, J.; Abdelaziz, M.; Perfectti, F. The Silent Extinction: Climate Change and the Potential Hybridization-Mediated Extinction of Endemic High-Mountain Plants. Biodivers. Conserv. 2015, 24, 1843–1857. [Google Scholar] [CrossRef]
- Lenoir, J.; Svenning, J.-C. Climate-Related Range Shifts—A Global Multidimensional Synthesis and New Research Directions. Ecography 2015, 38, 15–28. [Google Scholar] [CrossRef]
- Urban, M.C. Accelerating Extinction Risk from Climate Change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandel, B.; Monnet, A.-C.; Govaerts, R.; Vorontsova, M. Late Quaternary Climate Stability and the Origins and Future of Global Grass Endemism. Ann. Bot. 2017, 119, 279–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| LOW ** | CHO | SEK | STR | |||||
|---|---|---|---|---|---|---|---|---|
| Location | E24.0873 N35.2686 | E24.0813 N35.2734 | E24.0892 N35.2819 | E24.0549 N35.2940 | ||||
| Altitude (m a.s.l.) | 1664 | 1965 | 2160 | 2339 | ||||
| Mean species richness per area | 2001 | 2008 | 2001 | 2008 | 2001 | 2008 | 2001 | 2008 |
| 1 m2 quadrats mean ± 1 SE | 11.87 ± 0.87 | +0.06 | 3.75 ± 0.59 | −0.06 | 1.50 ± 0.22 | −0.12 | 1.44 ± 0.44 | +0.44 |
| 5 m SAS (mean of aspects) | 42.00 ± 1.47 | −2.25 | 16.50 ± 0.86 | –2.00 | 8.75 ± 0.75 | +0.50 | 6.50 ± 1.32 | +0.75 |
| 10 m SAS (mean of aspects) | 39.25 ± 0.62 | −4.00 | 17.00 ± 0.70 | +1.00 | 10.00 ± 1.29 | +0.25 | 8.50 ± 1.19 | +2.75 |
| 5 + 10 m SAS (mean of aspects) | 46.75 ± 0.47 | –2.25 | 20.50 ± 0.64 | –1.00 | 12.75 ± 0.47 | –0.75 | 9.50 ± 1.50 | +1.75 |
| Explanatory Variable | LOW * | CHO | SEK | STR |
|---|---|---|---|---|
| Time t | 0.00032 | 0.000096 | 0.00013 | 0.000112 |
| January | 10.51 | 8.222 | 7.705 | 6.266 |
| February | 10.51 | 7.995 | 7.684 | 6.553 |
| March | 11.39 | 8.585 | 6.972 | 5.964 |
| April | 11.25 | 8.300 | 5.720 | 5.422 |
| May | 12.11 | 9.598 | 7.802 | 7.351 |
| June | 12.05 | 9.537 | 8.514 | 7.504 |
| July | 12.56 | 10.03 | 8.716 | 8.117 |
| August | 12.69 | 10.05 | 8.833 | 7.863 |
| September | 11.41 | 8.769 | 7.481 | 6.205 |
| October | 10.82 | 7.940 | 6.758 | 5.488 |
| November | 9.309 | 6.531 | 5.329 | 4.193 |
| December | 9.588 | 7.006 | 6.350 | 5.427 |
| sin((2/365)*π*t) | −4.976 | −4.030 | −4.099 | −3.848 |
| cos((2/365)*π*t) | 8.371 | 7.475 | 7.370 | 7.339 |
| GR-LEO | 2001 | 2008 | Trend * | Lost | New |
|---|---|---|---|---|---|
| Total species | 70 | 66 |  | 8 | 4 |
| Endemics | 20 | 18 | | 3 | 1 |
| Summits | |||||
| LOW ** | 59 | 55 | | 6 | 2 |
| CHO | 32 | 28 | | 7 | 3 |
| SEK | 18 | 18 | -- | 2 | 2 |
| STR | 14 | 15 |  | 0 | 1 |
| Site/Sampling Scale | Model | t-Value | p-Value |
| LOW 5 m SAS * | Pair t-test | 1.0944 | 0.133 |
| LOW 10 m SAS | Pair t-test | 3.7033 | 0.003 * |
| CHO 5 m SAS | Pair t-test | 2.8284 | 0.0633 |
| CHO 10 m SAS | Pair t-test | 0.5624 | 0.6134 |
| SEK 10 m SAS | Pair t-test | 1.7321 | 0.1817 |
| SEK 10 m SAS | Pair t-test | 0.2641 | 0.8089 |
| STR 5 m SAS | Pair t-test | 1.5667 | 0.2151 |
| STR 10 m SAS | Pair t-test | 1.9612 | 0.1447 |
| Site/Sampling Scale | Model | R2 | sig.level |
| SAS 5–All sites | Linear regression against temp. means | 0.78 | 0.001 |
| SAS 5–All sites | Linear regression against min temp. means | 0.85 | 0.001 |
| SAS 5–All sites | Linear regression against max temp. means | 0.63 | 0.001 |
| SAS 10–All sites | Linear regression against temp. means | 0.86 | 0.001 |
| SAS 10–All sites | Linear regression against min temp. means | 0.9 | 0.001 |
| SAS 10–All sites | Linear regression against max temp. means | 0.76 | 0.001 |
| SAS–5 SR changes–all sites | Linear regression against temp. means | 0.34 | 0.05 |
| SAS–5 SR changes–all sites | Linear regression against min temp. means | 0.36 | 0.05 |
| SAS–5 SR changes–all sites | Linear regression against max temp. means | 0.27 | 0.05 |
| SAS–5 SR changes–all sites | Linear regression against June temp. means | 0.46 | 0.05 |
| SAS–10 SR changes–all sites | Linear regression against temp. means | 0.11 | n.s |
| SAS–10 SR changes–all sites | Linear regression against min temp. means | 0.16 | n.s |
| SAS–10 SR changes–all sites | Linear regression against max temp. means | 0.13 | n.s |
| SAS–10 SR changes–all sites | Linear regression against June temp. means | 0.14 | n.s |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kazakis, G.; Ghosn, D.; Remoundou, I.; Nyktas, P.; Talias, M.A.; Vogiatzakis, I.N. Altitudinal Vascular Plant Richness and Climate Change in the Alpine Zone of the Lefka Ori, Crete. Diversity 2021, 13, 22. https://doi.org/10.3390/d13010022
Kazakis G, Ghosn D, Remoundou I, Nyktas P, Talias MA, Vogiatzakis IN. Altitudinal Vascular Plant Richness and Climate Change in the Alpine Zone of the Lefka Ori, Crete. Diversity. 2021; 13(1):22. https://doi.org/10.3390/d13010022
Chicago/Turabian StyleKazakis, George, Dany Ghosn, Ilektra Remoundou, Panagiotis Nyktas, Michael A. Talias, and Ioannis N. Vogiatzakis. 2021. "Altitudinal Vascular Plant Richness and Climate Change in the Alpine Zone of the Lefka Ori, Crete" Diversity 13, no. 1: 22. https://doi.org/10.3390/d13010022
APA StyleKazakis, G., Ghosn, D., Remoundou, I., Nyktas, P., Talias, M. A., & Vogiatzakis, I. N. (2021). Altitudinal Vascular Plant Richness and Climate Change in the Alpine Zone of the Lefka Ori, Crete. Diversity, 13(1), 22. https://doi.org/10.3390/d13010022