Diversity and Origin of the Central Mexican Alpine Flora


Abstract
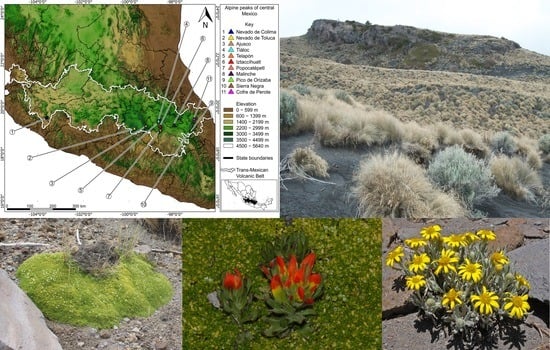
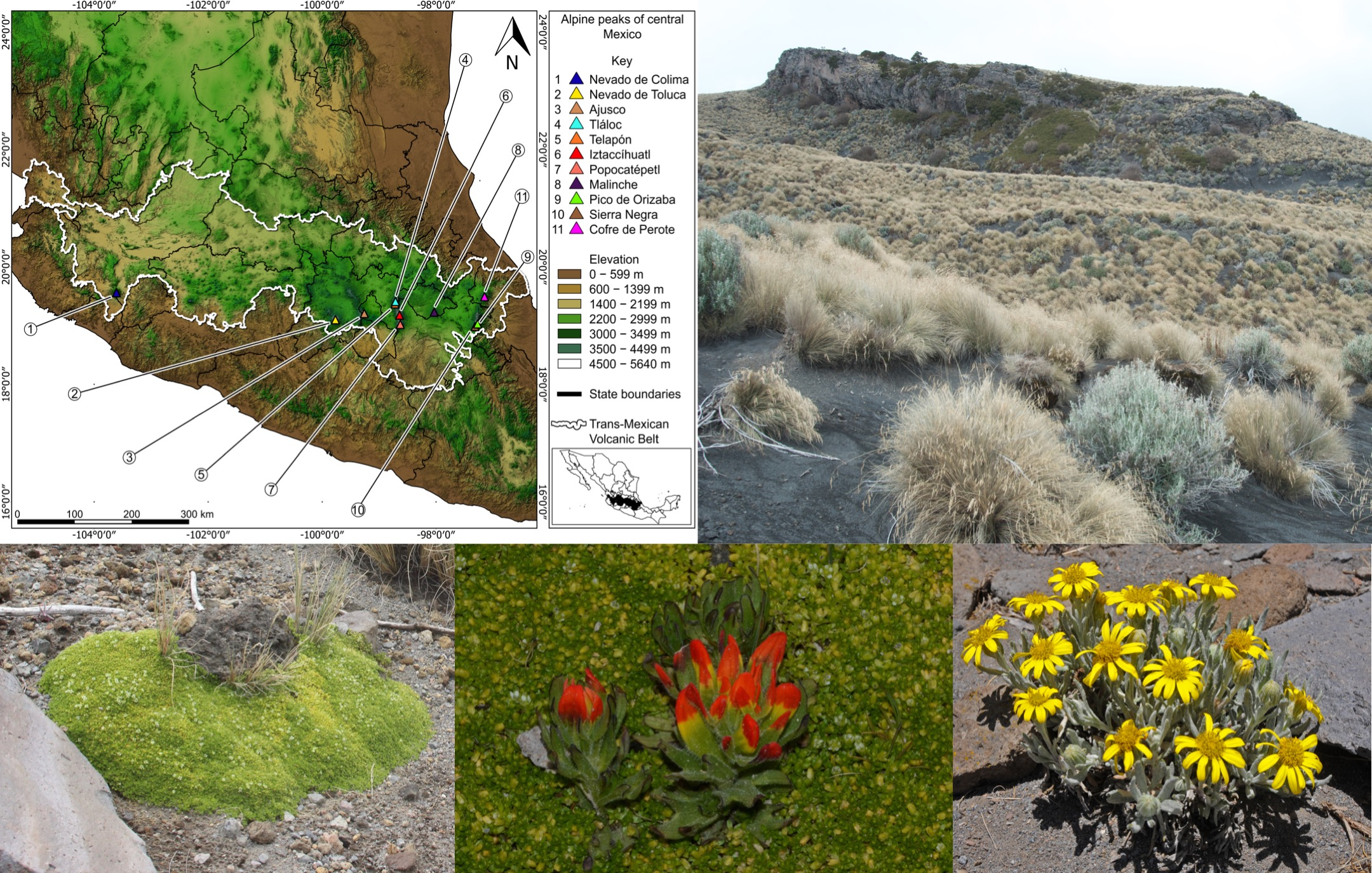
:
1. Introduction
2. Materials and Methods
3. Results
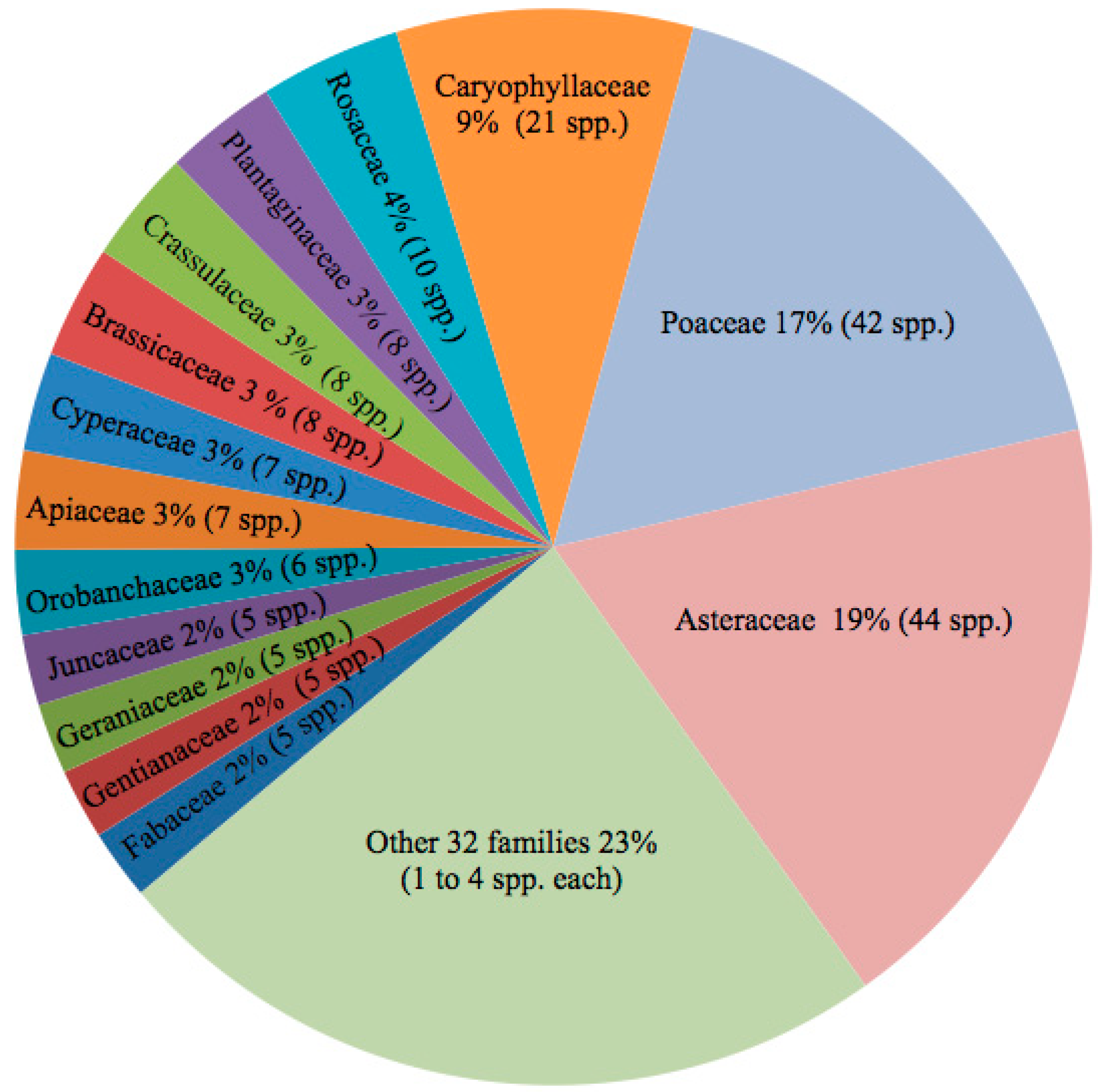
3.1. Diversity and Community Structure
3.2. Endemism
3.3. Phytogeography
3.4. Growth Forms
3.5. Conservation Status
4. Discussion
4.1. Diversity
4.2. Endemism
4.3. Introduced Taxa
4.4. Growth Forms
4.5. Phytogeography
4.6. Conservation
4.7. Upward Vegetational Shifts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | NC | NT | AJ | TL | TE | IZ | PP | ML | PO | SN | CP | Habit | Overall Distribution |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PTERIDOPHYTES | |||||||||||||
| ASPLENIACEAE | |||||||||||||
| Asplenium castaneum Schltdl. & Cham. | x | x | x | x | x | x | x | x | x | x | ph | Mexico to Bolivia and Peru | |
| Asplenium fibrillosum Pringle & Davenp. | x | ph | endemic to central Mexico | ||||||||||
| DRYOPTERIDACEAE | |||||||||||||
| Elaphoglossum mathewsii (Fée) T. Moore | x | x | ph | Mexico to Bolivia and Chile | |||||||||
| Polystichum speciosissimum (A. Braun ex Kunze) Copel. | x | x | x | x | x | x | x | x | ph | central Mexico to Panama | |||
| POLYPODIACEAE | |||||||||||||
| Alansmia spathulata (A.R. Sm.) Moguel & M. Kessler | x | x | ph | Mexico to Guatemala | |||||||||
| Melpomene peruviana (Desv.) A.R. Sm. & R.C. Moran | x | x | x | ph | disjunct between Mexico and South America | ||||||||
| Pleopeltis polylepis (Roem. ex Kunze) T. Moore var. polylepis | x | ph | USA to Guatemala | ||||||||||
| Polypodium calirhiza S.A. Whitmore & A.R. Sm. | x | ph | western USA to central Mexico | ||||||||||
| PTERIDACEAE | |||||||||||||
| Gaga marginata (H.B.K.) F.W. Li & Windham | x | ph | Mexico to Bolivia and Argentina | ||||||||||
| Myriopteris lendigera (Cav.) J. Sm. | x | ph | southwestern USA to Ecuador and the Antilles | ||||||||||
| Pellaea ternifolia (Cav.) Link | x | ph | USA to Argentina, Chile, and the Antilles | ||||||||||
| WOODSIACEAE | |||||||||||||
| Cystopteris fragilis (L.) Bernh. | x | x | x | x | x | x | x | x | x | x | ph | Canada to Chile and Argentina, also native to the Old World | |
| GYMNOSPERMS | |||||||||||||
| CUPRESSACEAE | |||||||||||||
| Juniperus monticola Martínez fo. compacta Martínez | x | x | x | x | x | x | x | x | x | x | x | sh | endemic to Mexico |
| PINACEAE | |||||||||||||
| Abies religiosa (H.B.K.) Schltdl. & Cham. | x | tr | Mexico to Guatemala | ||||||||||
| Pinus hartwegii Lindl. | x | x | x | x | x | x | x | x | x | x | x | tr | Mexico to Honduras |
| ANGIOSPERMS | |||||||||||||
| APIACEAE | |||||||||||||
| Chaerophyllum orizabae (I.M. Johnst.) K.F. Chung | x | x | x | x | ph | endemic to the TMVB | |||||||
| Chaerophyllum tolucanum (I.M. Johnst.) K.F. Chung | x | x | x | x | x | ph | endemic to the TMVB | ||||||
| Eryngium carlinae F. Delaroche | x | ph | Mexico to Panama | ||||||||||
| Eryngium proteiflorum F. Delaroche | x | x | x | x | x | x | x | x | x | ph | endemic to the TMVB | ||
| Lilaeopsis schaffneriana (Schltdl.) J.M. Coult. & Rose | x | ph | Mexico to Ecuador | ||||||||||
| Ottoa oenanthoides H.B.K. | x | ph | Mexico to Ecuador | ||||||||||
| Tauschia alpina (J.M. Coult. & Rose) Mathias | x | x | x | x | ph | endemic to the TMVB | |||||||
| ASTERACEAE | |||||||||||||
| Achillea millefolium L. | x | x | x | ph | nearly cosmopolitan in temperate regions | ||||||||
| Ageratina prunellifolia (H.B.K.) R.M. King & H. Rob. | x | x | x | x | ph | Mexico to Guatemala | |||||||
| Ageratina vernicosa (Sch. Bip. ex Greenm.) R.M. King & H. Rob. | x | sh | endemic to Mexico | ||||||||||
| Baccharis conferta H.B.K. | x | x | x | sh | endemic to Mexico | ||||||||
| Barkleyanthus salicifolius (H.B.K.) H. Rob & Brettell | x | x | sh | southwestern USA to Honduras | |||||||||
| Bidens anthemoides (DC.) Sherff | x | x | x | ph | endemic to Mexico | ||||||||
| Bidens triplinervia H.B.K. | x | x | x | x | x | x | x | x | x | sh | Mexico to Argentina | ||
| Chionolaena lavandulifolia (H.B.K.) Benth. & Hook. f. ex B.D. Jacks. | x | x | x | x | x | x | x | x | x | sub | Mexico to Central America | ||
| Chionolaena salicifolia (Bertol.) G.L. Nesom | x | sub | Mexico to Guatemala | ||||||||||
| Cirsium ehrenbergii Sch. Bip. | x | x | x | x | x | ph | endemic to Mexico | ||||||
| Cirsium jorullense (H.B.K.) Spreng. | x | ph | endemic to Mexico | ||||||||||
| Cirsium nivale (H.B.K.) Sch. Bip. | x | x | x | x | x | x | x | x | x | ph | endemic to the TMVB | ||
| Conyza coronopifolia H.B.K. | x | an | Mexico to South America | ||||||||||
| Cotula mexicana (DC.) Cabrera | x | ph | Mexico to South America | ||||||||||
| Dahlia merckii Lehm. | x | ph | endemic to Mexico | ||||||||||
| Hymenoxys integrifolia (H.B.K.) Bierner | x | ph | Mexico to Guatemala | ||||||||||
| Erigeron galeottii (A. Gray) Greene | x | x | x | x | x | x | ph | endemic to Mexico | |||||
| Gamochaeta americana (Mill.) Wedd. | x | x | x | ph | Canada to South America and also the Antilles | ||||||||
| Gamochaeta purpurea (L.) Cabrera | x | ph | Canada to South America | ||||||||||
| Gamochaeta standleyi (Steyerm.) G.L. Nesom | x | x | x | x | ph | Mexico to Guatemala | |||||||
| Hieracium mexicanum Less. | x | x | x | x | x | x | ph | Mexico to Guatemala | |||||
| Laennecia schiedeana (Less.) G.L. Nesom | x | x | x | x | x | x | x | x | x | an/ph | USA to Panama | ||
| Mexerion sarmentosum (Klatt) G.L. Nesom | x | x | x | x | x | ph | endemic to the TMVB | ||||||
| Osbertia stolonifera (DC.) Greene | x | x | ph | Mexico to Guatemala | |||||||||
| Oxylobus arbutifolius (H.B.K.) A. Gray | x | x | x | x | x | x | x | x | x | ph/sub | Mexico to Guatemala | ||
| Packera bellidifolia (H.B.K.) W.A. Weber & Á. Löve | x | x | x | x | x | x | x | ph | endemic to Mexico | ||||
| Packera toluccana (DC.) W.A. Weber & Á. Löve | x | x | x | x | x | x | ph | endemic to Mexico | |||||
| Pseudognaphalium inornatum (DC.) Anderb. | x | ph | endemic to Mexico | ||||||||||
| Pseudognaphalium liebmannii (Sch. Bip. ex Klatt) Anderb. | x | x | x | x | x | x | x | x | x | x | x | an/ph | endemic to Mexico |
| Pseudognaphalium nubicola (I.M. Johnst.) Anderb. | x | x | ph | endemic to Mexico | |||||||||
| Pseudognaphalium oxyphyllum (DC.) Kirp. | x | x | x | ph | Mexico to Guatemala | ||||||||
| Robinsonecio gerberifolius (Sch. Bip. ex Hemsl.) T.M. Barkley & Janovec | x | x | x | x | x | x | ph | Mexico to Guatemala | |||||
| Roldana angulifolia (DC.) H. Rob. & Brettell | x | x | x | sh | endemic to Mexico | ||||||||
| Roldana barba-johannis (DC.) H. Rob. & Brettell | x | x | sh | Mexico to Guatemala | |||||||||
| Roldana candicans (Née) Villaseñor, S. Valencia & Coombes | x | x | ph | endemic to Mexico | |||||||||
| Roldana reticulata (DC.) H. Rob. & Brettell | x | ph | endemic to Mexico | ||||||||||
| Selloa plantaginea H.B.K. | x | x | ph | endemic to the TMVB | |||||||||
| Senecio callosus Sch. Bip. | x | x | x | ph | Mexico to Guatemala | ||||||||
| Senecio cinerarioides H.B.K. | x | x | x | x | sh | endemic to Mexico | |||||||
| Senecio mairetianus DC. | x | x | x | x | x | x | x | x | x | x | sh | Mexico to Guatemala | |
| Senecio procumbens H.B.K. | x | x | x | x | x | x | x | ph | endemic to the TMVB | ||||
| Senecio roseus Sch. Bip. | x | x | x | x | x | x | x | x | x | ph | endemic to the TMVB | ||
| +Senecio vulgaris L. | x | an | introduced from the Old World | ||||||||||
| +Taraxacum officinale F.H. Wigg. | x | x | x | x | x | ph | introduced from the Old World | ||||||
| BERBERIDACEAE | |||||||||||||
| Berberis alpina Zamudio | x | x | x | x | x | x | x | sh | endemic to central Mexico | ||||
| BORAGINACEAE | |||||||||||||
| Hackelia mexicana (Schltdl. & Cham.) I.M. Johnst. | x | ph | Mexico to Bolivia and Peru | ||||||||||
| Lithospermum distichum Ortega | x | x | ph | Mexico to Guatemala | |||||||||
| Phacelia heterophylla Pursh | x | x | ph | USA to central Mexico | |||||||||
| Phacelia platycarpa (Cav.) Spreng. | x | x | x | x | x | x | x | x | x | ph | Mexico to Guatemala | ||
| BRASSICACEAE | |||||||||||||
| Barbarea orthoceras Ledeb. | x | ph | Canada to Mexico | ||||||||||
| Cardamine obliqua Hochst. ex A. Rich. var. stylosa Rollins | x | ph | disjunct between Mexico and east Africa | ||||||||||
| Descurainia impatiens (Schltd. & Cham.) O.E. Schulz | x | x | x | x | x | x | x | ph | Mexico to Guatemala | ||||
| Draba jorullensis H.B.K. | x | x | x | x | x | x | x | x | x | x | x | ph | Mexico to Guatemala |
| *Draba nivicola Rose | x | x | x | x | x | x | x | x | ph | endemic to the alpine TMVB | |||
| Erysimum capitatum (Douglas ex Hook.) Greene | x | x | ph | USA to Mexico | |||||||||
| Lepidium schaffneri Thell. | x | ph | endemic to Mexico | ||||||||||
| Romanschulzia arabiformis (DC.) Rollins | x | ph | endemic to Mexico | ||||||||||
| CAMPANULACEAE | |||||||||||||
| Lobelia nana H.B.K. | x | ph | Mexico to Guatemala | ||||||||||
| CARYOPHYLLACEAE | |||||||||||||
| Arenaria bourgaei Hemsl. | x | ph | Mexico to Ecuador | ||||||||||
| Arenaria bryoides Willd. ex Schltdl. | x | x | x | x | x | x | x | x | x | ph | Mexico to Guatemala | ||
| Arenaria lanuginosa (Michx.) Rohrb. | x | x | ph | USA to South America and the Antilles | |||||||||
| Arenaria lycopodioides Willd. ex Schltdl. | x | ph | Mexico to Guatemala | ||||||||||
| Arenaria oresbia Greenm. | x | x | x | x | x | x | x | ph | endemic to Mexico | ||||
| Arenaria parvifolia Benth. | x | x | x | x | x | x | x | x | ph | disjunct between Mexico and South America | |||
| Arenaria reptans Hemsl. | x | x | x | x | x | x | x | x | x | ph | Mexico, Central America, and South America | ||
| Cerastium brachypodum (Engelm. ex A. Gray) B.L. Rob. | x | ph | Canada to central Mexico | ||||||||||
| +Cerastium glomeratum Thuill. | x | ph | introduced from the Old World | ||||||||||
| Cerastium nutans Raf. | x | x | ph | Canada to Guatemala | |||||||||
| Cerastium orithales Schltdl. | x | x | x | x | x | x | x | ph | endemic to the TMVB | ||||
| *Cerastium purpusii Greenm. | x | x | ph | endemic to the alpine TMVB | |||||||||
| Cerastium ramigerum Bartl. | x | x | x | x | x | x | x | x | ph | endemic to the TMVB | |||
| Cerastium tolucense D.A. Good | x | x | x | x | x | ph | endemic to the TMVB | ||||||
| Cerastium vulcanicum Schltdl. | x | x | x | x | x | x | x | ph | Mexico to Guatemala | ||||
| Colobanthus quitensis (H.B.K.) Bartl. | x | x | x | ph | disjunct between Mexico and South America, also Antarctica | ||||||||
| Drymaria effusa A. Gray var. depressa (Greene) J.A. Duke | x | x | x | x | x | an | USA to Mexico | ||||||
| Sagina saginoides (L.) H. Karst. | x | x | ph | circumboreal, in the Americas from Canada and Greenland to central Mexico | |||||||||
| +Scleranthus annuus L. | x | an | introduced from the Old World | ||||||||||
| +Stellaria media (L.) Vill. | x | an/ph | introduced from the Old World | ||||||||||
| Stellaria umbellata Turzc. | x | ph | disjunct between central Mexico and the western USA and Canada, also in the Old World | ||||||||||
| COMMELINACEAE | |||||||||||||
| Weldenia candida Schult. f. | x | ph | Mexico to Guatemala | ||||||||||
| CRASSULACEAE | |||||||||||||
| Echeveria mucronata Schltdl. | x | ph | endemic to Mexico | ||||||||||
| Echeveria secunda Booth ex Lindl. | x | x | x | x | x | x | x | ph | endemic to central Mexico | ||||
| Echeveria subalpina Rose & Purpus | x | x | ph | endemic to the TMVB | |||||||||
| Sedum clavifolium Rose | x | ph | endemic to the TMVB | ||||||||||
| Sedum goldmanii (Rose) Moran | x | x | x | x | x | x | x | ph | endemic to central Mexico | ||||
| Sedum minimum Rose | x | x | x | x | x | x | ph | endemic to the TMVB | |||||
| Sedum obcordatum R.T. Clausen | x | x | x | ph | endemic to the TMVB | ||||||||
| Tillaea saginoides Maxim. | x | x | x | an | Alaska to southern Mexico, also native to the Old World | ||||||||
| CYPERACEAE | |||||||||||||
| Carex curviculmis Reznicek | x | x | x | ph | endemic to Mexico | ||||||||
| Carex echinata Murray | x | ph | USA to Guatemala | ||||||||||
| Carex geophila Mack. | x | ph | USA to Guatemala | ||||||||||
| Carex hermannii Cochrane | x | ph | endemic to the TMVB | ||||||||||
| Carex orizabae Liebm. | x | x | x | ph | Mexico to Guatemala | ||||||||
| Carex peucophila Holm. | x | x | x | x | ph | Mexico to Guatemala | |||||||
| Eleocharis acicularis (L.) Roem. & Schult. | x | ph | Greenland and Canada to Ecuador, also native to the Old World | ||||||||||
| ELATINACEAE | |||||||||||||
| Elatine brachysperma A. Gray | x | an/ph | USA to Mexico | ||||||||||
| ERICACEAE | |||||||||||||
| Comarostaphylis discolor (Hook.) Diggs | x | sh | Mexico to Guatemala | ||||||||||
| Monotropa hypopitys L. | x | ph | Alaska to Guatemala, also native to the Old World | ||||||||||
| Gaultheria myrsinoides H.B.K. | x | x | x | x | x | x | x | sh/sub | Mexico to South America | ||||
| Vaccinium cespitosum Michx. | x | x | x | x | x | x | x | x | x | sh/sub | Alaska and Canada to Guatemala | ||
| FABACEAE | |||||||||||||
| Astragalus hintonii Barneby | x | ph | endemic to the TMVB | ||||||||||
| Astragalus tolucanus B.L. Rob. & Seaton | x | ph | Mexico to Guatemala | ||||||||||
| Lupinus aschenbornii S. Schauer | x | x | x | x | x | x | ph | endemic to the TMVB | |||||
| Lupinus montanus H.B.K. | x | x | x | x | x | x | x | x | x | x | ph | Mexico to Guatemala | |
| Trifolium amabile H.B.K. | x | ph | Mexico to Costa Rica | ||||||||||
| GENTIANACEAE | |||||||||||||
| Gentiana ovatiloba Kusn. | x | x | ph | Mexico to Guatemala | |||||||||
| Gentiana perpusilla Brandegee | x | x | an | endemic to the TMVB | |||||||||
| Halenia brevicornis (H.B.K.) G. Don | x | an | Mexico to South America | ||||||||||
| Halenia plantaginea (H.B.K.) G. Don | x | x | x | x | ph | endemic to Mexico | |||||||
| Halenia pringlei B.L. Rob. & Seaton | x | x | x | x | x | x | x | ph | endemic to the TMVB | ||||
| GERANIACEAE | |||||||||||||
| +Erodium cicutarium (L.) L’Hér. ex Aiton | x | ph | introduced from the Old World | ||||||||||
| Geranium cruceroense R. Knuth | x | ph | endemic to central Mexico | ||||||||||
| Geranium latum Small | x | x | ph | endemic to central Mexico | |||||||||
| Geranium potentillifolium DC. | x | x | ph | endemic to Mexico | |||||||||
| Geranium seemannii Peyr. | x | ph | Mexico to South America | ||||||||||
| GROSSULARIACEAE | |||||||||||||
| Ribes ciliatum Humb. & Bonpl. ex Roem. & Schult. | x | x | x | x | x | x | x | x | sh | Mexico to Costa Rica | |||
| IRIDACEAE | |||||||||||||
| Sisyrinchium quadrangulatum Klatt | x | x | x | x | x | x | x | ph | endemic to the TMVB | ||||
| Sisyrinchium tenuifolium Humb. & Bonpl. ex Willd. | x | x | ph | endemic to Mexico | |||||||||
| JUNCACEAE | |||||||||||||
| Juncus balticus Willd. var. mexicanus (Willd. ex Schult. & Schult. f.) Snogerup | x | ph | Canada to Argentina, also native to the Old World | ||||||||||
| Luzula caricina E. Mey. | x | ph | Mexico to Guatemala | ||||||||||
| Luzula denticulata Liebm. | x | ph | Mexico to Panama | ||||||||||
| Luzula racemosa Desv. | x | x | x | x | x | x | x | x | x | x | x | ph | Mexico to South America |
| Luzula vulcanica Liebm. | x | x | x | ph | Mexico, Central America, and South America | ||||||||
| LAMIACEAE | |||||||||||||
| +Prunella vulgaris L. | x | ph/bi | introduced from the Old World | ||||||||||
| Salvia prunelloides H.B.K. | x | ph | endemic to Mexico | ||||||||||
| Stachys eriantha Benth. | x | x | x | x | x | x | x | ph | disjunct between Mexico and South America | ||||
| MELANTHIACEAE | |||||||||||||
| Anticlea frigida (Schltdl. & Cham.) Zomlefer & Jodd. | x | x | ph | endemic to Mexico | |||||||||
| MONTIACEAE | |||||||||||||
| Calandrinia acaulis H.B.K. | x | x | x | x | x | x | x | x | x | ph | Mexico to South America | ||
| Claytonia perfoliata Donn ex Willd. | x | ph | USA to Guatemala | ||||||||||
| Montia chamissoi (Ledeb. ex Spreng.) Greene | x | an/ph | Alaska and Canada to Mexico | ||||||||||
| Montia fontana L | x | an | temperate zones of the world | ||||||||||
| ONAGRACEAE | |||||||||||||
| Epilobium ciliatum Raf. | x | x | ph | Canada to South America | |||||||||
| OROBANCHACEAE | |||||||||||||
| Castilleja moranensis H.B.K. | x | x | ph | endemic to the TMVB | |||||||||
| Castilleja pectinata M. Martens & Galeotti | x | x | ph | Mexico to Guatemala | |||||||||
| Castilleja scorzonerifolia H.B.K. | x | x | x | x | x | x | x | ph | endemic to Mexico | ||||
| Castilleja tenuifolia M. Martens & Galeotti | x | ph | endemic to Mexico | ||||||||||
| *Castilleja tolucensis H.B.K. | x | x | x | x | x | x | x | endemic to the alpine TMVB | |||||
| Pedicularis orizabae Schltdl. & Cham. | x | ph | central Mexico to Guatemala | ||||||||||
| OXALIDACEAE | |||||||||||||
| Oxalis alpina (Rose) Rose ex R. Knuth | x | x | x | ph | USA to Guatemala | ||||||||
| PHRYMACEAE | |||||||||||||
| Erythranthe glabrata (H.B.K.) G.L. Nesom | x | ph | Canada to Argentina and Chile | ||||||||||
| PIPERACEAE | |||||||||||||
| Peperomia basiradicans G. Mathieu | x | x | ph | endemic to the TMVB | |||||||||
| PLANTAGINACEAE | |||||||||||||
| Callitriche heterophylla Pursh | x | x | ph | Greenland and Canada to South America | |||||||||
| Penstemon gentianoides (H.B.K.) Poir. | x | x | x | x | x | x | x | x | x | x | ph | Mexico to Guatemala | |
| Penstemon roseus (Cerv. ex Sweet) G. Don | x | x | x | ph | endemic to Mexico | ||||||||
| Plantago australis Lam. | x | ph | USA to South America | ||||||||||
| Plantago nivea H.B.K. | x | ph | Mexico to Guatemala | ||||||||||
| *Plantago tolucensis Pilg. | x | x | x | x | x | x | ph | endemic to the alpine TMVB | |||||
| Plantago tubulosa Decne. | x | ph | Mexico, Guatemala, and South America | ||||||||||
| Veronica serpyllifolia L. | x | x | ph | widespread in the northern hemisphere and the Americas | |||||||||
| POACEAE | |||||||||||||
| Agrostis bourgaei E. Fourn. | x | x | x | x | ph | endemic to the TMVB | |||||||
| Agrostis calderoniae Acosta Cast. | x | ph | endemic to the TMVB | ||||||||||
| Agrostis perennans (Walter) Tuck. | x | ph | Canada to Guatemala | ||||||||||
| Agrostis subpatens Hitchc. | x | x | x | x | ph | Mexico to Costa Rica | |||||||
| Agrostis tolucensis H.B.K. | x | x | x | x | x | x | x | x | x | x | x | ph | Mexico to Argentina and Chile |
| Bromus carinatus Hook. & Arn. | x | x | ph | USA to Costa Rica | |||||||||
| +Bromus diandrus Roth. | x | ph | introduced from the Old World | ||||||||||
| Bromus exaltatus Bernh. | x | x | ph | central Mexico to Panama | |||||||||
| Calamagrostis eriantha (H.B.K.) Steud. | x | x | x | ph | endemic to Mexico | ||||||||
| Calamagrostis orizabae (Rupr. ex Fourn.) Beal | x | x | x | x | ph | endemic to Mexico | |||||||
| Calamagrostis rigescens (J. Presl) Scribn. | x | x | ph | disjunct between Mexico and South America | |||||||||
| Calamagrostis tolucensis (H.B.K.) Trin. ex Steud. | x | x | x | x | x | x | x | x | ph | central Mexico to Honduras | |||
| Cinna poiformis (H.B.K.) Scribn. & Merr. | x | x | x | x | ph | central Mexico to Bolivia and Peru | |||||||
| Deschampsia liebmanniana (E. Fourn.) Hitchc. | x | x | x | x | ph | endemic to the TMVB | |||||||
| Festuca hephaestophila Nees | x | x | x | x | x | x | x | x | x | ph | central Mexico to Guatemala | ||
| Festuca jaliscana E.B. Alexeev. | x | ph | endemic to the TMVB | ||||||||||
| Festuca livida (H.B.K.) Willd. ex Spreng. | x | x | x | x | x | x | x | ph | central Mexico and Guatemala | ||||
| Festuca orizabensis E.B. Alexeev. | x | ph | endemic to Mexico | ||||||||||
| Festuca tolucensis H.B.K. | x | x | x | x | x | x | x | x | x | x | ph | Mexico to Colombia and Venezuela | |
| Muhlenbergia macroura (H.B.K.) Hitch. | x | ph | northern Mexico to Costa Rica | ||||||||||
| Muhlenbergia nigra Hitchc. | x | x | x | x | x | ph | central Mexico and Guatemala | ||||||
| Muhlenbergia orophila Swallen | x | ph | central Mexico and Guatemala | ||||||||||
| Muhlenbergia peruviana (P. Beauv.) Steud. | x | x | x | ph | southwestern USA to Guatemala; South America | ||||||||
| Muhlenbergia quadridentata (H.B.K.) Trin. | x | x | x | x | ph | northern Mexico to Guatemala | |||||||
| Muhlenbergia ramulosa (H.B.K.) Swallen | x | x | ph | southwestern USA to Costa Rica; Argentina | |||||||||
| Muhlenbergia tricholepis (Torr.) Columbus | x | x | ph | western USA to central Mexico | |||||||||
| Muhlenbergia utilis (Torr.) Hitchc. | x | ph | southwestern USA to southern Mexico | ||||||||||
| Nassella mexicana (Hitchc.) R.W. Pohl | x | x | ph | central Mexico to Argentina | |||||||||
| Peyritschia koelerioides (Peyr.) E. Fourn. | x | x | x | ph | Mexico to Guatemala | ||||||||
| Phleum alpinum L. | x | x | ph | circumboreal, in the Americas from Canada and Greenland to Chile and Argentina | |||||||||
| +Poa annua L. | x | x | x | x | x | x | x | ph | introduced from the Old World | ||||
| Poa calycina (Ball) Refulio var. mathewsii (Ball) Refulio | x | x | x | x | x | x | ph | disjunct between Mexico and South America | |||||
| Poa chamaeclinos Pilg. | x | ph | disjunct between Mexico and South America | ||||||||||
| Poa gymnantha Pilg. | x | ph | disjunct between Mexico and South America | ||||||||||
| Poa orizabensis Hitchc. | x | x | x | ph | Mexico to Guatemala | ||||||||
| +Poa pratensis L. var. pratensis | x | ph | introduced | ||||||||||
| Poa scaberula Hook. f. | x | x | x | x | x | ph | Mexico to South America | ||||||
| Stipa ichu (Ruiz & Pav.) Kunth | x | x | x | ph | Mexico to South America | ||||||||
| Trisetum ligulatum Finot & Zuloaga | x | ph | endemic to Mexico | ||||||||||
| Trisetum rosei Scribn. & Merr. | x | x | x | ph | Mexico to Guatemala | ||||||||
| Trisetum spicatum (L.) K. Richt. | x | x | x | x | x | x | x | x | x | x | x | ph | Canada and Greenland to Chile and Argentina, also native to the Old World |
| +Vulpia myuros (L.) C.C. Gmel. | x | ph | introduced from the Old World | ||||||||||
| POLEMONIACEAE | |||||||||||||
| Polemonium grandiflorum Benth. | x | ph | endemic to Mexico | ||||||||||
| POLYGONACEAE | |||||||||||||
| +Rumex acetosella L. | x | x | ph | introduced from the Old World | |||||||||
| RANUNCULACEAE | |||||||||||||
| Ranunculus donianus Pritz. ex Walp. | x | x | x | x | x | x | x | x | ph | Mexico to Guatemala | |||
| Ranunculus multicaulis D. Don ex G. Don | x | x | x | x | x | ph | Mexico to Central America | ||||||
| Ranunculus peruvianus Pers. | x | ph | Mexico to South America | ||||||||||
| Ranunculus praemorsus H.B.K. ex DC. | x | ph | Mexico to South America | ||||||||||
| ROSACEAE | |||||||||||||
| Acaena elongata L. | x | x | x | sh | Mexico to South America | ||||||||
| Alchemilla aphanoides Mutis ex L. f. | x | x | ph | Mexico to Central America | |||||||||
| Alchemilla pinnata Ruiz & Pav. | x | x | ph | Mexico to Guatemala; disjunct in South America | |||||||||
| Alchemilla pringlei (Rydb.) Fedde | x | x | x | x | ph | Mexico to Guatemala | |||||||
| Alchemilla procumbens Rose | x | x | x | x | x | x | x | ph | Mexico to South America | ||||
| Alchemilla vulcanica Schltdl. & Cham. | x | x | x | x | x | x | x | x | x | x | x | ph | Mexico to South America |
| Potentilla candicans Humb. & Bonpl. ex Nestl. | x | ph | endemic to the TMVB | ||||||||||
| Potentilla ranunculoides H.B.K. | x | x | x | x | x | x | x | ph | endemic to Mexico | ||||
| Potentilla richardii Lehm. | x | x | x | x | x | x | x | x | x | ph | endemic to the TMVB | ||
| Sibbaldia procumbens L. | x | x | x | x | ph | circumboreal, in the Americas from Canada and Greenland to Mexico | |||||||
| RUBIACEAE | |||||||||||||
| Crusea longiflora (Roem. & Schult.) W.R. Anderson | x | an | Mexico to Central America | ||||||||||
| SALICACEAE | |||||||||||||
| Salix paradoxa H.B.K. | x | tr/sh | endemic to Mexico | ||||||||||
| SCROPHULARIACEAE | |||||||||||||
| Limosella aquatica L. | x | x | x | an | also native to the Old World | ||||||||
| SOLANACEAE | |||||||||||||
| Solanum demissum Lindl. | x | x | x | ph | central Mexico to Guatemala | ||||||||
| URTICACEAE | |||||||||||||
| Urtica orizabae Liebm. | x | ph | endemic to the TMVB | ||||||||||
| VERBENACEAE | |||||||||||||
| Glandularia teucriifolia (M. Martens & Galeotti) Umber. | x | x | ph | Mexico to Guatemala | |||||||||
| VIOLACEAE | |||||||||||||
| Viola beamanii Calderón | x | x | ph | endemic to the TMVB | |||||||||
| Viola hemsleyana Calderón | x | ph | endemic to Mexico | ||||||||||
Appendix B
References
- Rzedowski, J. Vegetación de México, 1st ed.; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: México City, Mexico, 2006; p. 504. [Google Scholar]
- Beaman, J.H. The timberlines of Iztaccihuatl and Popocatepetl. Ecology 1962, 43, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Körner, C.H. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems, 2nd ed.; Springer Science & Business Media: New York, NY, USA, 2003; p. 349. [Google Scholar]
- Billings, W.D. Adaptations and origins of alpine plants. Arct. Alp. Res. 1974, 6, 129–142. [Google Scholar] [CrossRef]
- Miranda, F. La Vegetación de Chiapas; Gobierno del estado de Chiapas: Tuxtla Gutiérrez, Mexico, 1952; p. 324. [Google Scholar]
- Beaman, J.H.; Andersen, J.W. The vegetation, floristics and phytogeography of the summit of Cerro Potosí, Mexico. Am. Midl. Naturalist. 1966, 75, 1–33. [Google Scholar] [CrossRef]
- McDonald, J. The alpine-subalpine flora of northeastern Mexico. SIDA 1990, 14, 21–28. [Google Scholar]
- García-Arévalo, A.; González-Elizondo, S. Flora y vegetación de la cima del Cerro Potosí, Nuevo León, México. Acta Bot. Mex. 1991, 13, 53–74. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Tuena, A.; Orozco-Esquivel, M.T.; Ferrari, L. Igneous petrogenesis of the Trans-Mexican volcanic belt. Geo. Soc. Amer. Spec. Pap. 2007, 422, 129–181. [Google Scholar]
- Almeida-Leñero, L.; Escamilla, M.; De Azcárate, J.G.; González-Trápaga, A.; Cleef, A.M. Vegetación alpina de los volcanes Popocatépetl, Iztaccíhuatl y Nevado de Toluca. In Biodiversidad de la Faja Volcánica Transmexicana; Luna, I., Morrone, J.J., Espinosa, D., Eds.; FES-Zaragoza-CONABIO: México City, Mexico, 2007; pp. 179–198. [Google Scholar]
- García, E. Modificaciones al Sistema de Clasificación de Köppen, 2nd ed.; Instituto de Geofísica, Universidad Nacional Autónoma de México: México City, Mexico, 1973; p. 246. [Google Scholar]
- Lauer, W.; Klaus, D. Geoecological investigations on the timberline of Pico de Orizaba, Mexico. Arct. Alp. Res. 1975, 7, 315–330. [Google Scholar] [CrossRef]
- Smith, A.P.; Young, T.P. Tropical alpine plant ecology. Annu. Rev. Ecol. Syst. 1987, 18, 137–158. [Google Scholar] [CrossRef]
- Hedberg, O. Features of afroalpine plant ecology. Acta Phytogeogr. Suecica 1964, 49, 1–44. [Google Scholar]
- Rundel, P.W. Tropical alpine climates. In Tropical Alpine Environments: Plant Form and Function; Rundel, P.W., Smith, A.P., Meinzer, F.C., Eds.; Cambridge University Press: Cambridge, UK, 1994; pp. 21–44. [Google Scholar]
- Emiliani, C. Paleotemperature analysis of Caribbean cores P6304-8 and P6304-9 and a generalized temperature curve for the past 425,000 years. J. Geol. 1996, 74, 109–124. [Google Scholar] [CrossRef]
- Mastretta-Yanes, A.; Moreno-Letelier, A.; Piñero, D.; Jorgensen, T.H.; Emerson, B.C. Biodiversity in the Mexican highlands and the interaction of geology, geography and climate within the Trans-Mexican Volcanic Belt. J. Biogeogr. 2015, 42, 1586–1600. [Google Scholar] [CrossRef]
- McDonald, J.A. Fitogeografía e historia de la flora alpina-subalpina del noreste de México: Orígenes y Distribución. In Diversidad Biológica de México; Ramamoorthy, T., Bye, R., Lot, A., Fa, A., Eds.; Instituto de Biología, Universidad Nacional de México: México City, Mexico, 1998; pp. 665–686. [Google Scholar]
- Buytaert, W.; Cuesta-Camacho, F.; Tobón, C. Potential impacts of climate change on the environmental services of humid tropical alpine regions. Glob. Ecol. Biogeogr. 2011, 20, 19–33. [Google Scholar] [CrossRef]
- Ernakovich, J.G.; Hopping, K.A.; Berdanier, A.B.; Simpson, R.T.; Kachergis, E.J.; Steltzer, H.; Wallenstein, M.D. Predicted responses of arctic and alpine ecosystems to altered seasonality under climate change. Glob. Chang. Biol. 2014, 20, 3256–3269. [Google Scholar] [CrossRef] [PubMed]
- Pauli, H.; Gottfried, M.; Reiter, K.; Klettner, C.; Grabherr, G. Signals of range expansions and contractions of vascular plants in the high Alps: Observations (1994–2004) at the GLORIA* master site Schrankogel, Tyrol, Austria. Glob. Chang. Biol. 2007, 13, 147–156. [Google Scholar] [CrossRef]
- Rehfeldt, G.; Crookston, N.; Sáenz, C.; Campbell, E. North American vegetation model for land-use planning in a changing climate: A solution to large classification problems. Ecol. Appl. 2012, 22, 119–141. [Google Scholar] [CrossRef]
- Seimon, T.A.; Seimon, A.; Daszak, P.; Halloy, S.R.; Schloegel, L.M.; Aguilar, C.A.; Simmons, E. Upward range extension of Andean anurans and chytridiomycosis to extreme elevations in response to tropical deglaciation. Glob. Change Biol. 2007, 13, 288–299. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Araújo, M.B.; Sykes, M.T.; Prentice, I.C. Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. USA. 2005, 102, 8245–8250. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Amezcua, Y.; Steinmann, V.W.; Ruiz-Sanchez, E.; Rojas-Soto, O.R. Mexican alpine plants in the face of global warming: Potential extinction within a specialized assemblage of narrow endemics. Biodivers. Conserv. 2016, 25, 865–888. [Google Scholar] [CrossRef]
- Maza-Villalobos, S.; Cotler, H.; Almeida-Leñero, L.; Hoth, J.; Steinmann, V.W.; Mastretta, A.; Hernández-Cárdenas, R.A. Conservando el pastizal alpino mexicano. Conocimientos, amenazas y esperanzas. Biodiversitas 2019, 142, 12–16. [Google Scholar]
- Scherrer, P.; Pickering, C.M. Effects of grazing, tourism and climate change on the alpine vegetation of Kosciuszko National Park. Vic. Naturalist. 2001, 118, 93. [Google Scholar]
- Fernández, F.J.A.; López-Domínguez, J.C. Biodiversidad del Parque Nacional La Malinche; Gobierno del Estado de Tlaxcala: Tlaxcala, Mexico, 2005; p. 224. [Google Scholar]
- Narave, H. La vegetación del cofre de Perote, Veracruz, Mexico. Biótica 1985, 10, 35–64. [Google Scholar]
- Hernández-Cárdenas, R.A.; Mendoza-Ruiz, A.; Arredondo-Amezcua, L.; Steinmann, V.W. The Alpine Ferns of the Trans-Mexican Volcanic Belt. Am. Fern J. 2019, 109, 11–25. [Google Scholar] [CrossRef]
- Neyra, J.A. Guía de Las Altas Montañas de México y Una de Guatemala, 1st ed.; CONABIO y Gobierno Federal: México City, Mexico, 2012; p. 413. [Google Scholar]
- McVaugh, J. Gramineae. In Flora Novo-Galiciana: A Descriptive Account of the Vascular Plants of Western Mexico; Anderson, J.R., Ed.; Ann Arbor University of Michigan Press: Ann Arbor, MI, USA, 1983; Volume 14, pp. 1–436. [Google Scholar]
- De Rzedowski, G.C.; Rzedowski y colaboradores, J. Flora Fanerogámica del Valle de México, 2nd ed.; 1a reimp.; @Instituto de Ecología, A.C. y Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Pátzcuaro, Mexico, 2005; p. 1406. [Google Scholar]
- Durán-Espinosa, C. Grossulariaceae. In Flora de Veracruz; Sosa, V., Ed.; Instituto de Ecología, A.C. y Centro de Investigaciones Tropicales: Xalapa, Mexico, 2001; Volume 122, pp. 1–15. [Google Scholar]
- Villareal, J.A. Gentianaceae. In Flora de Veracruz; Sosa, V., Ed.; Instituto de Ecología, A.C. y Centro de Investigaciones Tropicales: Xalapa, Mexico, 2001; Volume 121, pp. 1–67. [Google Scholar]
- Thiers, B. Index Herbariorum: A global directory of public herbaria and associated staff. New York Botanical Garden’s Virtual Herbarium. Available online: http://sweetgum.nybg.org/science/ih (accessed on 8 November 2020).
- Lot, A.; Chiang, F. Manual de Herbario: Administración y Manejo de Colecciones, Técnicas de Recolección y Preparación de Ejemplares Botánicos; Consejo Nacional de la Flora de México: Ciudad de México, Mexico, 1986. [Google Scholar]
- Mickel, J.T.; Smith, A.R. The Pteridophytes of Mexico (Memoirs of the New York Botanical Garden 88); The New York Botanical Garden: New York, NY, USA, 2004; p. 1054. [Google Scholar]
- Sigel, E.M.; Windham, M.D.; Haufler, C.H.; Pryer, K.M. Phylogeny, divergence time estimates, and phylogeography of the diploid species of the Polypodium vulgare complex (Polypodiaceae). Syst. Bot. 2014, 39, 1042–1055. [Google Scholar] [CrossRef]
- Ppg, I.; Shmakov, A. A community-based classification for extant ferns and lycophytes. J. Syst. Evol. 2016, 6, 563–603. [Google Scholar] [CrossRef]
- Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; Stevens, P.F. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. 2016, 181, 1–20. [Google Scholar]
- GBIF: The Global Biodiversity Information Facility. Available online: https://www.gbif.org (accessed on 15 June 2017).
- Tropicos.org. Missouri Botanical Garden. Available online: http://www.tropicos.org (accessed on 10 January 2015).
- iNaturalist Application. Available online: https://www.inaturalist.org/ (accessed on 1 November 2020).
- IUCN 2020. The IUCN Red List of Threatened Species. Version 2020-3. Available online: www.iucnredlist.org (accessed on 10 February 2020).
- Secretaría de Medio Ambiente y Recursos Naturales. Norma Oficial Mexicana NOM-059-SEMARNAT-2010. Protección Ambiental-Especies nativas de México de flora y fauna silvestres-Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio. Lista de especies en riesgo. D. Of. De La Fed. De México 2010, 30, 1–78. [Google Scholar]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environm. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Zamudio, S. Notas sobre el género Berberis (Berberidaceae) en México. Acta Bot. Mex. 2009, 87, 31–70. [Google Scholar] [CrossRef] [Green Version]
- Villaseñor, J.L. Checklist of the native vascular plants of Mexico. Rev. Mex. Biodivers. 2016, 87, 559–902. [Google Scholar] [CrossRef] [Green Version]
- Körner, C.H. Alpine plant diversity: A global survey and functional interpretations. In Arctic and Alpine Biodiversity: Patterns, Causes and Ecosystem Consequences; Chapin, F.S., Körner, C., Eds.; Springer: Berlin/Heidelberg, Germany, 1995. [Google Scholar]
- Luteyn, J.L. Páramos, A Checklist of Plant Diversity, Geographical Distribution, and Botanical Literature; New York Botanical Garden Press: New York, NY, USA, 1999; p. 278. [Google Scholar]
- Rai, I.D.; Adhikari, B.S.; Rawa, G.S. Floral diversity along subalpine and alpine ecosystems in Tungnath area of Kedarnath wildlife sanctuary, Uttarakhand. Indian Forester. 2012, 138, 927–940. [Google Scholar]
- Hoxey, P. Notes on the cacti of the Sandia valley, Puno, Peru and a description of a new species of Echinopsis (Cactaceae). Bradleya 2016, 34, 187–199. [Google Scholar] [CrossRef]
- Delgadillo, C. Musgos alpinos del Nevado de Colima, México. Bot. Sci. 1987, 47, 7–11. [Google Scholar] [CrossRef]
- Sklenář, P.; Balslev, H. Geographic flora elements in the Ecuadorian superpáramo. Flora-Morphol. Distrib. Funct. Ecol. Plants 2007, 202, 50–61. [Google Scholar] [CrossRef]
- Klimes, L. Life-forms and clonality of vascular plants along an altitudinal gradient in E Ladakh (NW Himalayas). Basic Appl. Ecol. 2003, 4, 317–328. [Google Scholar] [CrossRef] [Green Version]
- Ángel, R.; Conrad, R.; Dvorsky, M.; Kopecky, M.; Kotilínek, M.; Hilesalu, I.; Schweingruber, F.; Dolezal, J. The root-associated microbial community of the world’s highest growing vascular plants. Microb. Ecol. 2016, 72, 394–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuesta, F.; Muriel, P.; Beck, S.; Meneses, R.I.; Halloy, S.; Salgado, S.; Ortiz, E.; Becerra, M.T. (Eds.) Biodiversidad y Cambio Climático en los Andes Tropicales—Conformación de Una Red de Investigación Para Monitorear sus Impactos y Delinear Acciones de Adaptación; Red Gloria-Andes: Lima, Ecuador, 2012; p. 180. [Google Scholar]
- Vargas, G.; Sánchez, J.J. Plantas con flores de los páramos de Costa Rica y Panamá: El páramo ístmico. In Páramos de Costa Rica; Kappelle, M., Horn, S.P., Eds.; INBio: San José, Costa Rica, 2005; pp. 397–435. [Google Scholar]
- Ferrari, L.; Orozco-Esquivel, T.; Manea, V.; Manea, M. The dynamic history of the Trans-Mexican Volcanic Belt and the Mexico subduction zone. Tectonophysics 2012, 522, 122–149. [Google Scholar] [CrossRef]
- Soreng, R.J.; Peterson, P.M. Revision of Poa, L. (Poaceae, Pooideae, Poeae, Poinae) in Mexico: New records, re-evaluation of P. ruprechtii, and two new species, P. palmeri and P. wendtii. PhytoKeys 2012, 15, 1–104. [Google Scholar] [CrossRef] [Green Version]
- Dehshiri, M.M.; Safikhani, K.; Mostafavi, H. Alpine flora of some part of Alvand Mountain, Hamadan province. J. Pl. Biol. 2016, 30, 89–104. [Google Scholar]
- Mark, A.F.; Dickinson, K.J.M.; Hofstede, R.G.M. Alpine vegetation, plant distribution, life forms, and environments in a perhumid New Zealand Region: Oceanic and tropical high mountain affinities. Arct. Antarc. Alp. Res. 2000, 32, 240–254. [Google Scholar] [CrossRef]
- Smith, A.P. Introduction to tropical vegetation. In Tropical Alpine Environments: Plant Form and Function; Rundel, P.W., Smith, A.P., Meinzer, F.C., Eds.; Cambridge University Press: Cambridge, UK, 1994; pp. 1–19. [Google Scholar]
- Aubert, S.; Boucher, F.; Lavergne, F.; Renaud, J.; Choler, P. 1914–2014: A revised worldwide catalogue of cushion plants 100 years after Hauri and Schröter. Alp. Bot. 2014, 124, 59–70. [Google Scholar] [CrossRef]
- Morrone, J.J. Fundamental biogeographic patterns across the Mexican Transition Zone: An evolutionary approach. Ecography 2010, 33, 335–361. [Google Scholar] [CrossRef]
- Gámez, N.; Escalante, T.; Rodríguez, G.; Linaje, M.; Morrone, J.J. Caracterización biogeográfica de la Faja Volcánica Transmexicana y análisis de los patrones de distribución de su mastofauna. Rev. Mex. Biodivers. 2012, 81, 258–272. [Google Scholar] [CrossRef] [Green Version]
- Sarmiento, G. Ecological Features of Climate. In High Tropical Mountains. High Altitude Tropical Biogeography; Vuilleumer, F., Monasterio, M., Eds.; Oxford University Press: New York, NY, USA, 1986; pp. 11–45. [Google Scholar]
- Rzedowski, J. An Ecological and Phytogeographical Analysis of the Grasslands of Mexico. Taxon 1975, 24, 67–80. [Google Scholar] [CrossRef]
- Ramírez-Villegas, J.; Cuesta, F.; Devenish, C.; Peralvo, M.; Jarvis, A.; Arnillas, C.A. Using species distributions models for designing conservation strategies of Tropical Andean biodiversity under climate change. J. Nat. Conserv. (Jena) 2014, 22, 391–404. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.-C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Ghosn, D. Recent plant diversity changes on Europe’s mountain summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef] [Green Version]
- Heilprin, A. The Temperate and Alpine Floras of the Giant Volcanoes of Mexico. Proc. Am. Philos. Soc. 1892, 30, 4–22. [Google Scholar]




| Volcano | Location | Elevation (m) | Summit Coordinates | Alpine Area (km2) | Number of Species | Last Eruption (ya) |
|---|---|---|---|---|---|---|
| El Ajusco | Ciudad de México | 3940 | 19°12′25″ N | 2 | 63 | 1620 * |
| 99°15′35″ W | ||||||
| Telapón | Estado de México and Puebla | 4080 | 19°22′15.42″ N | 0.03 | 57 | 30,000 n |
| 98°43′12.42″ W | ||||||
| Tláloc | Estado de México and Puebla | 4140 | 19°24′41.7″ N | 0.97 | 72 | 30–31,000 n |
| 98°42′44.8″ W | ||||||
| Cofre de Perote | Veracruz | 4230 | 19°29′39.24″ N | 1.57 | 93 | 900 * |
| 97°08′51.12″ W | ||||||
| Nevado de Colima | Jalisco | 4280 | 19°33′46.92″ N | 0.91 | 31 | 30–35,000 n |
| 103°36′30.66″ W | ||||||
| Malinche | Puebla and Tlaxcala | 4440 | 19°13′51.54″ N | 3.55 | 62 | 3190 * |
| 98°01′55.26″ W | ||||||
| Sierra Negra | Puebla | 4580 | 18°59′07.68″ N | 5.23 | 46 | unknown |
| 97°18′51.24″ W | ||||||
| Nevado de Toluca | Estado de México | 4690 | 19°06′06.72″ N | 15.27 | 102 | 3370 * |
| 99°46′03.36″ W | ||||||
| Iztaccíhuatl | Puebla and Estado de México | 5220 | 19°10′48.4″ N | 38.58 | 170 | 20,000 (fissurelike eruption at 9000) n |
| 98°38′29.3″ W | ||||||
| Popocatépetl | Estado de México, Puebla and Morelos | 5460 | 19°01′18.4″ N | 32.79 | 81 | currently active * |
| 98°37′39.6″ W | ||||||
| Pico de Orizaba | Veracruz y Puebla | 5640 | 19°01′48″ N | 48.83 | 78 | 174 * |
| 97°16′10″ W |
| Distribution | Number of Species (Percentage of Flora) |
|---|---|
| Endemic to the study area | 4 (1.7%) |
| Endemic to the study area and adjacent subalpine forest | 8 (3.4%) |
| Endemic to the Trans-Mexican Volcanic Belt | 36 (14.8%) |
| Endemic to Mexico | 76 (32.1%) |
| Mexico to Guatemala | 41 (17.3%) |
| Mexico to Central America (beyond Guatemala) | 15 (6.3%) |
| Mexico, Central America, and South America | 29 (12.2%) |
| Disjunct between Mexico and South America | 9 (3.8%) |
| Canada and/or USA to Mexico | 9 (3.8%) |
| Canada and/or USA to Central/South America | 20 (8.4%) |
| Also native to the Old World | 15 (6.3%) |
| Introduced from the Old World | 12 (5.1%) |
| Region | Genera |
|---|---|
| Endemic to Mexico | Mexerion |
| USA/Canada to Mexico | Hymenoxys, Tauschia |
| USA/Canada to Central America | Comarostaphylis, Laennecia, Roldana |
| Mexico to Central America | Dahlia, Osbertia, Oxylobus, Romanschulzia, Weldenia |
| USA/Canada to South America | Barkleyanthus, Crusea, Gaga, Glandularia, Nassela, Phacelia |
| Mexico to South America | Alansmia, Chionolaena, Echeveria, Melpomene, Ottoa, Peyritschia, Selloa |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steinmann, V.W.; Arredondo-Amezcua, L.; Hernández-Cárdenas, R.A.; Ramírez-Amezcua, Y. Diversity and Origin of the Central Mexican Alpine Flora. Diversity 2021, 13, 31. https://doi.org/10.3390/d13010031
Steinmann VW, Arredondo-Amezcua L, Hernández-Cárdenas RA, Ramírez-Amezcua Y. Diversity and Origin of the Central Mexican Alpine Flora. Diversity. 2021; 13(1):31. https://doi.org/10.3390/d13010031
Chicago/Turabian StyleSteinmann, Victor W., Libertad Arredondo-Amezcua, Rodrigo Alejandro Hernández-Cárdenas, and Yocupitzia Ramírez-Amezcua. 2021. "Diversity and Origin of the Central Mexican Alpine Flora" Diversity 13, no. 1: 31. https://doi.org/10.3390/d13010031
APA StyleSteinmann, V. W., Arredondo-Amezcua, L., Hernández-Cárdenas, R. A., & Ramírez-Amezcua, Y. (2021). Diversity and Origin of the Central Mexican Alpine Flora. Diversity, 13(1), 31. https://doi.org/10.3390/d13010031