Genome Insights into the Novel Species Jejubacter calystegiae, a Plant Growth-Promoting Bacterium in Saline Conditions

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain, Growth Conditions, and Genomic DNA Extraction
2.2. Whole-Genome Sequencing and Assembly
2.3. Genome Annotation and Property Analysis
2.4. Evaluation of Strain KSNA2 on Seed Germination in Saline Conditions
2.5. Nucleotide Sequence Accession Numbers
3. Results and Discussion
3.1. Genomic Features
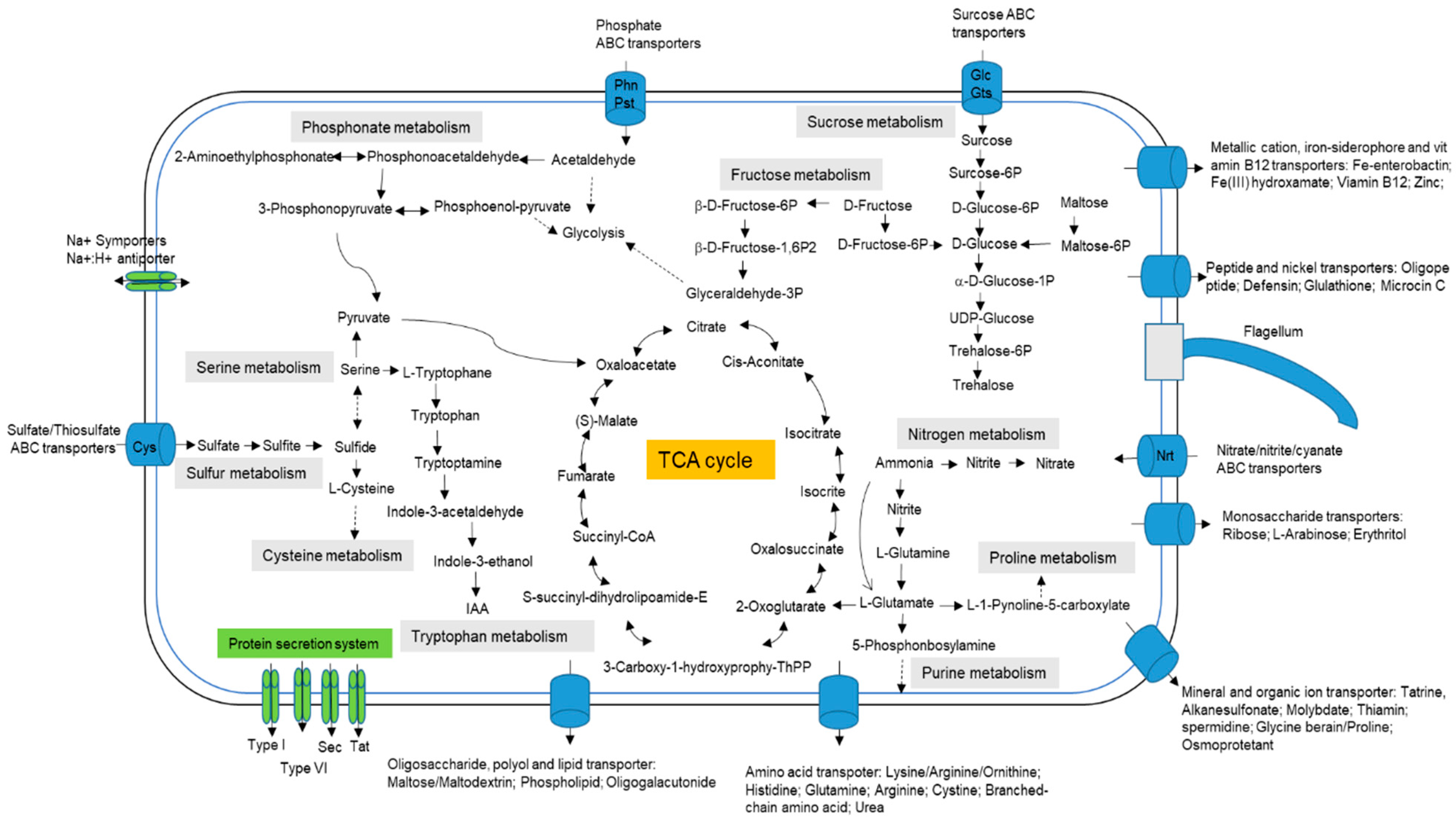
3.2. Genes of Central Metabolism and Cellular Processes
3.3. Genes Associated with Multi-Stress Response
3.4. Secondary Metabolites and Salt Tolerance Pathway
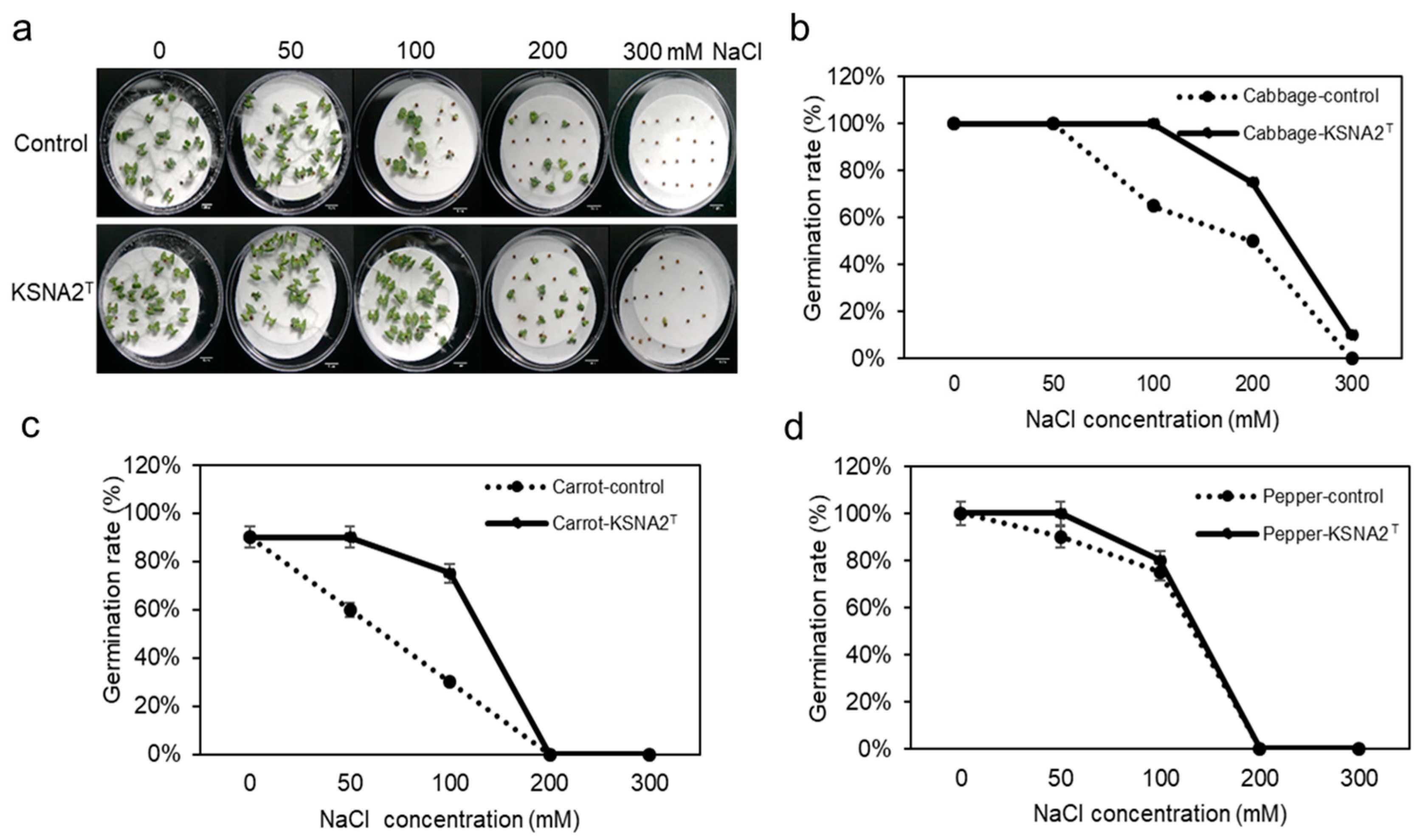
3.5. Strain KSNA2T Improves Seed Germination in Saline Conditions
4. Discussion
4.1. Potential Ecological Role of Strain KSNA2T
4.2. Potential Role of Strain KSNA2T as an Agricultural Reagent
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tool for its alleviation. Saudi J. Biol. Sci. 2015, 2, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.; Redman, R. More than 400 million years of evolution and some plants still can’t make it on their own: Plant stress tolerance via fungal symbiosis. J. Exp. Bot. 2008, 59, 1109–1114. [Google Scholar] [CrossRef]
- Rodriguez, R.J.; Henson, J.; Van Volkenburgh, E.; Hoy, M.; Wright, L.; Beckwith, F.; Kim, Y.O.; Redman, R.S. Stress tolerance in plants via habitat-adapted symbiosis. ISME J. 2008, 2, 404–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redman, R.S.; Kim, Y.O.; Woodward, C.J.; Greer, C.; Espino, L.; Doty, S.L.; Rodriguez, R.J. Increased fitness of rice plants to abiotic stress via habitat adapted symbiosis: A strategy for mitigating impacts of climate change. PLoS ONE 2011, 6, e14823. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Xin, K.; Liu, H.; Cheng, J.; Shen, X.; Wang, Y.; Zhang, L. Pantoea alhagi, a novel endophytic bacterium with ability to improve growth and drought tolerance in wheat. Sci. Rep. 2017, 7, 41564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodd, I.C.; Perez-Alfocea, F. Microbial amelioration of crop salinity stress. J. Exp. Bot. 2012, 63, 3415–3428. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Wang, D.; Lee, J.-S.; Kim, D.-H.; Jeong, J.C.; Kim, C.Y.; Kim, S.W.; Lee, J. Jejubacter calystegiae gen. nov., sp. nov., moderately halophilic, a new member of the family Enterobacteriaceae, isolated from beach morning glory. J. Microbiol. 2020, 58, 357–366. [Google Scholar] [CrossRef]
- Lin, L.; Li, Z.; Zhang, X.; Chang, S.; Yang, L.; Li, Y.; An, Q. Plant growth-promoting nitrogen-fixing enterobacteria are in association with sugarcane plants growing in Guanxi, China. Microbes Environ. 2012, 27, 391–398. [Google Scholar] [CrossRef] [Green Version]
- Vitorino, L.C.; Silva, F.O.; Cruvinel, B.G.; Bessa, L.A.; Rosa, M.; Souchie, E.L.; Silva, F.G. Biocontrol potential of Sclerotinia sclerotiorum and physiological changes in soybean in response to Butia archeri Palm Rhizobacteria. Plants 2020, 9, 64. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG tools for functional characterization of genome and metagenome sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, 81–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinheinz, K.A.; Joensen, K.G.; Larsen, M.V. Applying the ResFinder and VirulenceFinder web-services for easy identification of acquired antibiotic resistance and E. coli virulence genes in bacteriophage and prophage nucleotide sequences. Bacteriophage 2014, 4, e27943. [Google Scholar] [CrossRef] [Green Version]
- Teng, J.L.; Yeung, M.L.; Chan, E.; Jia, L.; Lin, C.H.; Huang, Y.; Tse, H.; Wong, S.S.; Sham, P.C.; Lau, S.K.; et al. PacBio But Not Illumina Technology can achieve fast, accurate and complete closure of the high GC, complex burkholderia pseudomallei Two-Chromosome genome. Front. Microbiol. 2017, 8, 1448. [Google Scholar] [CrossRef] [Green Version]
- Grant, J.R.; Stothard, P. The CGView server: A comparative genomics tool for circular genomes. Nucleic Acids Res. 2008, 36, 181–184. [Google Scholar] [CrossRef]
- Spaepen, S.; Vanderleyden, J.; Remans, M. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef] [Green Version]
- Geddes, B.A.; Ryu, M.H.; Mus, F.; Garcia Costas, A.; Peters, J.W.; Voigt, C.A.; Poole, P. Use of plant colonizing bacteria as chassis for transfer of N2-fixation to cereals. Curr. Opin. Biotechnol. 2015, 32, 216–222. [Google Scholar] [CrossRef]
- Shailendra, S.; Govind, G. Plant Growth promoting rhizobacteria (PGPR): Current and future prospects for development of sustainable agriculture. J. Microb. Biochem. Technol. 2015, 7, 96–102. [Google Scholar] [CrossRef]
- Kielak, A.M.; Cipriano, M.A.; Kuramae, E.E. Acidobacteria strains from subdivision 1 act as plant growth-promoting bacteria. Arch. Microbiol. 2016, 198, 987–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.I.; Kim, I.-H.; Nam, T.-J. Crude extract and solvent fractions of Calystegia soldanella induce G1 and S phase arrest of the cell cycle in HepG2 cells. Int. J. Oncol. 2017, 20, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Bae, K.H. The Medicinal Plants of Korea; Kyo-Hak Publishing: Seoul, Korea, 2000. [Google Scholar]
- Shabala, S.; Shabala, L. Ion transport and osmotic adjustment in plants and bacteria. Biomol. Concepts 2011, 2, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Vurukonda, S.; Giovanardi, D.; Stefani, E. Plant growth promoting and biocontrol activity of Streptomyces spp. as endophytes. Int. J. Mol. Sci. 2018, 19, 952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, F.; Yang, L.; Tai, F.; Hu, X.; Wang, W. “Omics” of maize stress response for sustainable food production: Opportunities and challenges. OMICS 2014, 18, 714–732. [Google Scholar] [CrossRef] [Green Version]
- Vardharajula, S.; Skz, A.; Vurukonda, S.S.K.P.; Shrivastava, M. Plant growth promoting endophytes and their interaction with plants to alleviate abiotic stress. Curr. Biotechnol. 2017, 6, 252–263. [Google Scholar] [CrossRef]
- Numan, M.; Bashir, S.; Khan, Y.; Mumtaz, R.; Shinwari, Z.K.; Khan, A.L.; Khan, A.; Harrasi, A. Plant growth promoting bacteria as an alternative strategy for salt tolerance in plants: A review. Microbiol. Res. 2018, 209, 21–32. [Google Scholar] [CrossRef]
- Etesami, H.; Beattie, G.A. Mining halophytes for plant growth-promoting halotolerant bacteria to enhance the salinity tolerance of non-halophytic crops. Front. Microbiol. 2018, 9, 148. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Singh, S.; Gaurav, A.K.; Srivastava, S.; Verma, J.P. Plant growth-promoting bacteria: Biological tools for the mitigation of salinity stress in plants. Front. Microbiol. 2020, 11, 1216. [Google Scholar] [CrossRef]
- Wu, T.Y.; Wu, X.-Q.; Xu, X.; Kong, W.; Wu, F. Salt tolerance mechanism and species identification of the plant rhizosphere bacterium JYZ-SD2. Curr. Microbiol. 2020, 77, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, P.; Kashyap, P.L.; Bhardwaj, A.K.; Kuppusamy, P.; Srivastava, A.K.; Tiwari, R.K. Bacterial endophyte mediated plant tolerance to salinity: Growth responses and mechanisms of action. World J. Microbiol. Biotechnol. 2020, 36, 26. [Google Scholar] [CrossRef] [PubMed]
- Vijaranakul, U.; Xiong, A.; Lockwood, K.; Jayaswal, R.K. Cloning and nucleotide sequencing of a Staphylococcus aureus gene encoding a branched-chain-amino-acid transporter. Appl. Environ. Microbiol. 1998, 64, 763–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scybert, S.; Pechous, R.; Sitthisak, S.; Nadakavukaren, M.J.; Wilkinson, B.J.; Jayaswal, R.K. NaCl-sensitive mutant of Staphylococcus aureus has a Tn917-lacZ insertion in its ars operon. FEMS Microbiol. Lett. 2003, 222, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Albdaiwi, R.N.; Khyami-Horani, H.; Ayad, J.Y.; Alananbeh, K.M.; Al-Sayaydeh, R. Isolation and Characterization of Halotolerant Plant Growth Promoting Rhizobacteria From Durum Wheat (Triticum turgidum subsp. durum) Cultivated in Saline Areas of the Dead Sea Region. Front. Microbiol. 2019, 10, 1639. [Google Scholar] [CrossRef] [Green Version]
- Amin, U.S.; Lash, T.D.; Wilkinson, B.J. Proline betaine is a highly effective osmoprotectant for Staphylococcus aureus. Arch. Microbiol. 1995, 163, 138–142. [Google Scholar] [CrossRef]
- Peddie, B.A.; Lever, M.; Randall, K.; Chambers, S.T. Osmoprotective activity, urea protection, and accumulation of hydrophilic betaines in Escherichia coli and Staphylococcus aureus. Antonie van Leeuwenhoek 1999, 75, 183–189. [Google Scholar] [CrossRef]
- Schwan, W.R.; Shibata, S.; Aizawa, S.; Wolfe, A.J. The two-component response regulator RcsB regulates type 1 piliation in Escherichia coli. J. Bacteriol. 2007, 189, 7159–7163. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Categories | Count | Ratio (%) | |
|---|---|---|---|
| Information storage and processing | |||
| J | Translation, ribosomal structure, and biogenesis | 184 | 3.9561 |
| A | RNA processing and modification | 1 | 0.0215 |
| K | Transcription | 344 | 7.3963 |
| L | Replication, recombination, and repair | 185 | 3.9776 |
| Cellular process and signaling | |||
| B | Chromatin structure and dynamics | 0 | 0.0000 |
| D | Cell cycle control, cell division, chromosome partitioning | 41 | 0.8815 |
| Y | Nuclear structure | 0 | 0.0000 |
| V | Defense mechanisms | 58 | 1.2470 |
| T | Signal transduction mechanisms | 149 | 3.2036 |
| M | Cell wall/membrane/envelope biogenesis | 265 | 5.6977 |
| N | Cell motility | 48 | 1.0320 |
| Z | Cytoskeleton | 0 | 0.0000 |
| W | Extracellular structures | 0 | 0.0000 |
| U | Intracellular trafficking, secretion, and vesicular transport | 80 | 1.7201 |
| O | Posttranslational modification, protein turnover, chaperones | 140 | 3.0101 |
| Metabolism | |||
| C | Energy production and conversion | 291 | 6.2567 |
| G | Carbohydrate transport and metabolism | 357 | 7.6758 |
| E | Amino acid transport and metabolism | 379 | 8.1488 |
| F | Nucleotide transport and metabolism | 100 | 2.1501 |
| H | Coenzyme transport and metabolism | 142 | 3.0531 |
| I | Lipid transport and metabolism | 102 | 2.1931 |
| P | Inorganic ion transport and metabolism | 266 | 5.7192 |
| Q | Secondary metabolites biosynthesis, transport, and catabolism | 75 | 1.6126 |
| Poorly characterized | |||
| R | General function prediction only | 139 | 2.9886 |
| S | Function unknown | 1305 | 28.0585 |
| Totals | 4651 | 100 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, L.M.; Lee, Y.J.; Han, H.L.; Lee, M.H.; Jeong, J.C.; Kim, C.Y.; Kim, S.W.; Lee, J.Y. Genome Insights into the Novel Species Jejubacter calystegiae, a Plant Growth-Promoting Bacterium in Saline Conditions. Diversity 2021, 13, 24. https://doi.org/10.3390/d13010024
Jiang LM, Lee YJ, Han HL, Lee MH, Jeong JC, Kim CY, Kim SW, Lee JY. Genome Insights into the Novel Species Jejubacter calystegiae, a Plant Growth-Promoting Bacterium in Saline Conditions. Diversity. 2021; 13(1):24. https://doi.org/10.3390/d13010024
Chicago/Turabian StyleJiang, Ling Min, Yong Jae Lee, Ho Le Han, Myoung Hui Lee, Jae Cheol Jeong, Cha Young Kim, Suk Weon Kim, and Ji Young Lee. 2021. "Genome Insights into the Novel Species Jejubacter calystegiae, a Plant Growth-Promoting Bacterium in Saline Conditions" Diversity 13, no. 1: 24. https://doi.org/10.3390/d13010024
APA StyleJiang, L. M., Lee, Y. J., Han, H. L., Lee, M. H., Jeong, J. C., Kim, C. Y., Kim, S. W., & Lee, J. Y. (2021). Genome Insights into the Novel Species Jejubacter calystegiae, a Plant Growth-Promoting Bacterium in Saline Conditions. Diversity, 13(1), 24. https://doi.org/10.3390/d13010024