
Diversity and Ecology of Diatoms in Pliocene Deposits of the Tunka Valley (Baikal Rift Zone)

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
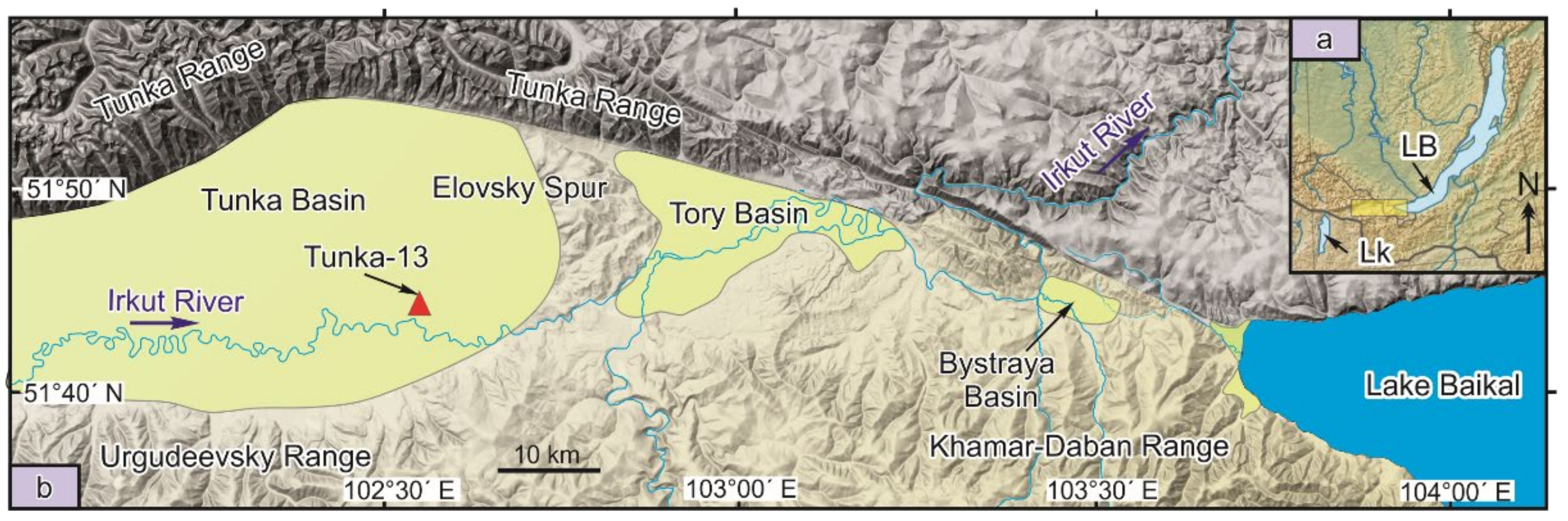
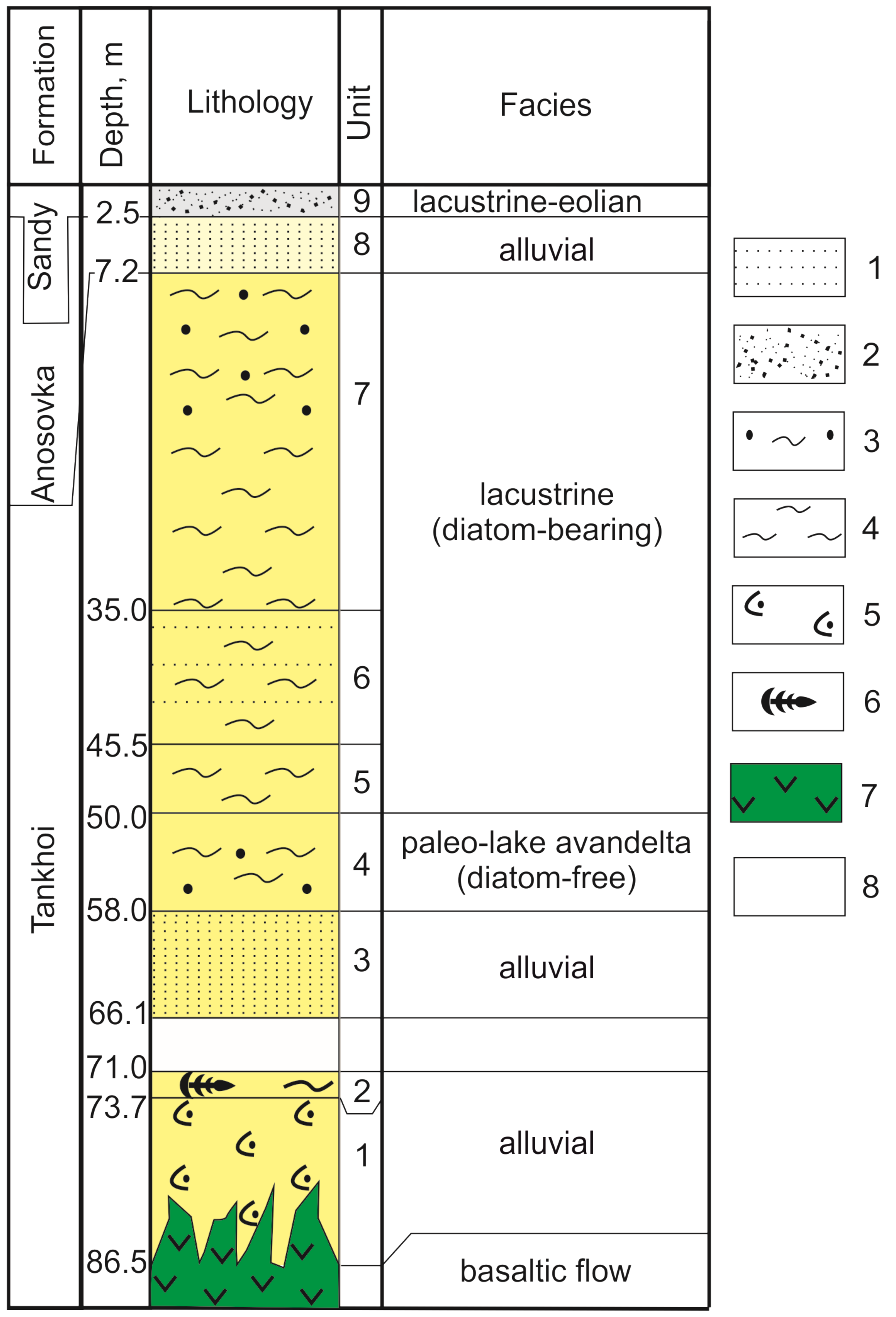
2.1. Core Sampling and Lithology
2.2. Diatom Analysis
3. Results
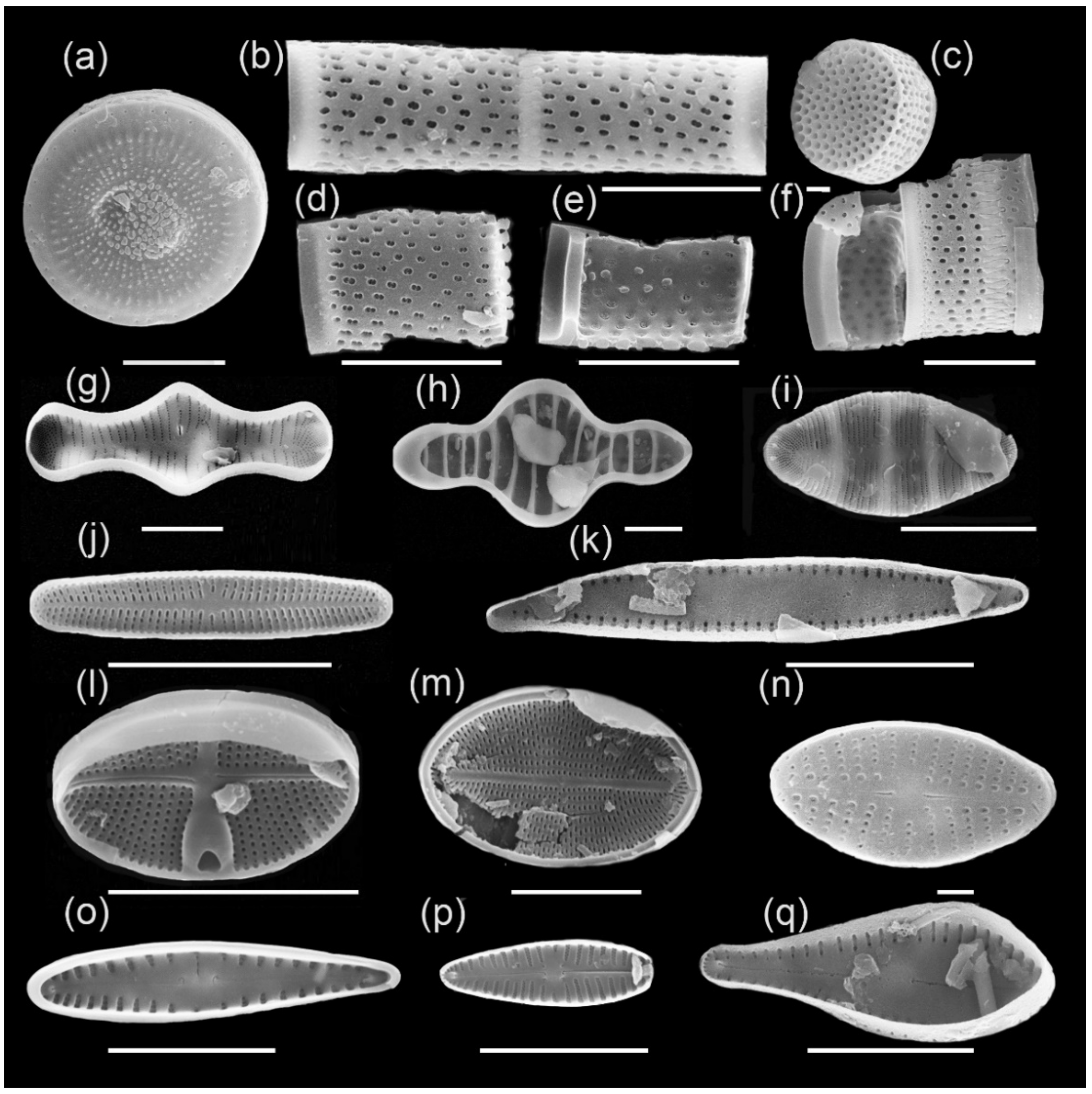
3.1. Diatom Flora
3.2. Diatom Biostratigraphy
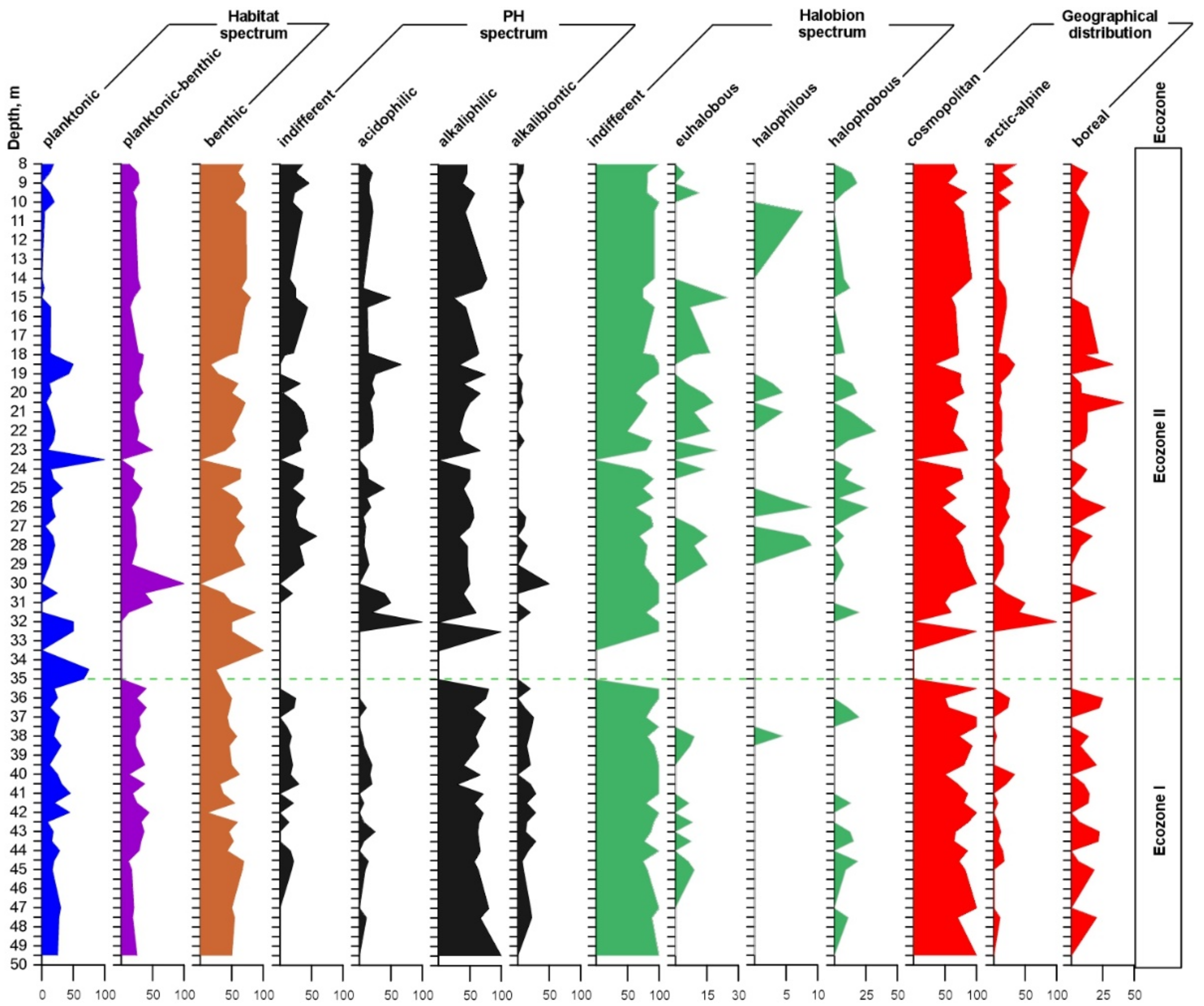
3.3. Ecological Analysis of Diatom Flora
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mats, V.D.; Ufimtsev, G.F.; Mandelbaum, M.M. The Cenozoic Baikal Rift Basin: Its Structure and Geological History; Academic Publishing House Geo: Novosibirsk, Russia, 2001; p. 252. (In Russian) [Google Scholar]
- Kuz’min, M.I.; Khursevich, G.K.; Prokopenko, A.A.; Fedenya, S.A.; Karabanov, E.B. Late Cenozoic Centric Diatoms of Lake Baikal; Academic Publishing House Geo: Novosibirsk, Russia, 2009; p. 370. (In Russian) [Google Scholar]
- Logachev, N.A.; Zorin, Y.A. Baikal Rift Zone: Structure and geodynamics. Tectonophysics 1992, 208, 273–286. [Google Scholar] [CrossRef]
- Rasskazov, S.V. Magmatism of the Baikal Rift System; Nauka: Novosibirsk, Russia, 1993; p. 288. (In Russian) [Google Scholar]
- Chuvashova, I.S.; Rasskazov, S.V. Sources of Magmatism in the Mantle of the Evolving Earth; ISU Publishing House: Irkutsk, Russia, 2014; p. 291. (In Russian) [Google Scholar]
- Cheremisinova, E.A. Diatom Flora of Neogene Sediments in Pribaikalie; Nauka: Novosibirsk, Russia, 1973; p. 83. (In Russian) [Google Scholar]
- Popova, S.M.; Mats, V.D.; Chernyaeva, G.P.; Shimaraeva, М.К.; Kul’chitsky, А.А.; Vorobyova, G.А.; Klimanova, V.М.; Kononov, Y.; Kravchinsky, А.Y.; Kulagina, N.V.; et al. Baikal Rift Zone: Paleolimnological Reconstructions; Nauka: Novosibirsk, Russia, 1989; p. 111. (In Russian) [Google Scholar]
- Lupikina, E.G.; Khursevich, G.K. New genus of the class Centrophyceae (Bacillariophyta). Algologiya 1991, 1, 67–70. [Google Scholar]
- Khursevich, G.K. Morphology and taxonomy of some centric diatom species from the Miocene sediments of the Dzhilinda and Tunka hollow. Mem. Calif. Acad. Sci. 1994, 17, 269–280. [Google Scholar]
- Likhoshway, E.V.; Pomazkina, G.V.; Nikiteeva, T.A. Centric diatoms from the Miocene deposits of the Baikal Rift Zone (Tunka depression). Russ. Geol. Geophys. 1997, 38, 1445–1452. [Google Scholar]
- Hassan, A.; Rasskazov, S.V.; Chuvashova, I.S.; Yasnygina, T.A.; Titova, L.A.; Kulagina, N.V.; Usoltseva, M.V. Identifying Upper Miocene—Lower Pliocene lacustrine sediments in dry Tunkabasin of the Baikal Rift Zone. Geodyn. Tectonophys. 2020, 11, 262–284. [Google Scholar] [CrossRef]
- Rasskazov, S.V.; Lyamina, N.A.; Chernyaeva, G.P.; Luzina, I.V.; Rudnev, A.F.; Rezanov, I.N. Cenozoic Stratigraphy of the Vitim Plateau: Phenomenon of Long-Term Rifting in the South of East Siberia; Academic Publishing House Geo: Novosibirsk, Russia, 2007; p. 193. (In Russian) [Google Scholar]
- Logatchev, N.A. Cenozoic continental deposits in the Baikal type basins. Izv. AN SSSR 1958, 4, 18–29. (In Russian) [Google Scholar]
- Grachev, M.A.; Likhoshvay, E.V.; Vorobyova, S.S.; Khlystov, O.M.; Bezrukova, E.V.; Veinberg, E.V.; Goldberg, E.L.; Granina, L.Z.; Kornakova, E.G.; Lazo, F.I.; et al. Paleoclimate signals of the Upper Pleistocene in sediments of Lake Baikal. Geol. Geophys. 1997, 38, 957–980. [Google Scholar]
- Skvortzow, B.W.; Meyer, C.I. A Contribution to the diatoms of Baikal Lake. Proc. Sungaree River Biol. Stn. 1928, 1, 1–55. [Google Scholar]
- Skvortzow, B.W. Bottom diatoms from Olhongate of Lake Baikal. Philipp. J. Sci. 1937, 62, 293–377. [Google Scholar]
- Gleser, S.I.; Jousé, A.P.; Makarova, I.V.; Proschkina-Lavrenko, A.I.; Sheshukova-Poretzkaya, V.S. The Diatoms of the USSR Fossil and Recent; Nauka: Leningrad, Russia, 1974; p. 403. (In Russian) [Google Scholar]
- Houk, V. Atlas of Freshwater Centric Diatoms with a Brief Key and Descriptions: Part. I. Melosiraceae, Orthoseiraceae, Paraliaceae and Aulacoseiraceae; Fottea: Olomouc, Czech Republic, 2003; pp. 1–27. [Google Scholar]
- Houk, V.; Klee, R. Atlas of Freshwater Centric Diatoms with a Brief Key and Descriptions: Part. II. Melosiraceae and Aulacoseiraceae; Fottea: Olomouc, Czech Republic, 2007; pp. 85–255. [Google Scholar]
- Kozyrenko, T.; Strelnikova, N.; Khursevich, G.; Tsoy, I.; Yakovschikova, T.; Mukhina, V.; Olshtynskaja, A.; Semina, G. The Diatoms of Russia and Adjacent Countries Fossil and Recent II; Strelnikova, N., Tsoy, I., Eds.; St. Petersburg University Press: St. Petersburg, Russia, 2008; p. 171. (In Russian) [Google Scholar]
- Gleser, S.I.; Makarova, I.V.; Moisseeva, A.I.; Nokolaev, V.A. (Eds.) The Diatoms of the USSR. Fossil and Recent; Nauka: Leningrad, Russia, 1988; Volume 2, Issue 1, p. 116. (In Russian) [Google Scholar]
- Gleser, S.I.; Makarova, I.V.; Moisseeva, A.I.; Nokolaev, V.A. (Eds.) The Diatoms of the USSR. Fossil and Recent; Nauka: St. Petersburg, Russia, 1992; Volume 2, Issue 2, p. 125. (In Russian) [Google Scholar]
- Loseva, E.I. Atlas of the Late Pliocene Diatoms of River Kama Region; Nauka: Leningrad, Russia, 1982; p. 204. (In Russian) [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. Teil 1: Naviculaceae, Bd. 2/1: Süβwasser flora von Mitteleuropa; Jena VEB Gustav Fischer Verlag: Stuttgart, Germany, 1986; p. 876. [Google Scholar]
- Lange-Bertalot, H.; Krammer, K. Achnanthes eine Monographie der gattungmit Definition der Gattung Cocconeis und Nachträgenzu den Naviculaceae; Bibl. Diatom: Berlin, Germany; Stuttgart, Germany, 1989; p. 385. [Google Scholar]
- Lange-Bertalot, H. Diatoms of Europe: Navicula sensu stricto, 10 Genera Separated from Navicula sensu lato Frustulia; Gantner, A.R.G., Ed.; Ruggell: Lichtenstein, Germany, 2001; p. 526. [Google Scholar]
- Krammer, K. Die Cymbelloid en Diatomeen. Teil 2. Encyonema part., Encyonopsis and Cymbellopsis; Bd. 37; Bibl. Diatom: Berlin, Germany; Stuttgart, Germany, 1997; p. 469. [Google Scholar]
- Krammer, K. Diatoms of Europe: The genus Pinnularia; Gantner, A.R.G., Ed.; Ruggell: Lichtenstein, Germany, 2000; p. 703. [Google Scholar]
- Krammer, K. Diatoms of Europe: Cymbella; Gantner, A.R.G., Ed.; Ruggell: Lichtenstein, Germany, 2002; p. 584. [Google Scholar]
- Krammer, K. Diatoms of Europe: Cymbopleura, Delicata, Navicymbula, Gomphocymbellopsis, Afrocymbella; Gantner, A.R.G., Ed.; Ruggell: Lichtenstein, Germany, 2003; p. 530. [Google Scholar]
- Kulikovskiy, M.S.; Lange-Bertalot, H.; Metzeltin, D.; Witkowski, A. Diatoms of Europe: Lake Baikal: Hotspot of Endemic Diatoms I; Gantner, A.R.G., Ed.; Ruggell: Lichtenstein, Germany, 2012; Volume 23, p. 861. [Google Scholar]
- Kulikovskiy, M.S.; Lange-Bertalot, H.; Kuznetsova, I.V. Iconograph Diatom: Lake Baikal: Hotspot of Endemic Diatoms II; Gantner, A.R.G., Ed.; Ruggell: Lichtenstein, Germany, 2015; Volume 26, p. 656. [Google Scholar]
- Kulikovskiy, M.S.; Glushchenko, A.M.; Genkal, S.I.; Kuznetsova, I.V. Identification Book of Diatoms from Russia; Dorofeyuk, N.I., Ed.; Filigran: Yaroslav, Russia, 2016; p. 804. (In Russian) [Google Scholar]
- Pomazkina, G.B.; Rodionova, Е.V. Diatoms of the Family Cymbellaceae of Lake Baikal: Atlas and Key; Nauka: Novosibirsk, Russia, 2014; p. 242. (In Russian) [Google Scholar]
- Pomazkina, G.B.; Rodionova, Е.V.; Sherbakova, T.A. Benthic Diatoms of the Family Naviculaceae of Lake Baikal: Atlas and Key; Nauka: Novosibirsk, Russia, 2018; p. 315. (In Russian) [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. 2021. Available online: http://www.algaebase.org (accessed on 5 June 2021).
- Sládeèek, V. Diatoms as indicators of organic Pollution. Acta Hydrochim. Et Hydrobiol. 1986, 14, 555–566. [Google Scholar] [CrossRef]
- Getsen, M.V.; Stenina, A.S. Algae (Systematic List): In Flora and Fauna of Water Bodies in the European North; Nauka: Leningrad, Russia, 1978; pp. 109–150. (In Russian) [Google Scholar]
- Van Dam, H.; Mertens, A.; Sinkeldam, J. A coded checklist and ecological indicator values of freshwater diatoms from the Netherlands. Netherland J. Aquat. Ecol. 1994, 1, 117–133. [Google Scholar]
- Barinova, S.S.; Medvedeva, L.A. Atlas of Algae as Saprobic Indicators (Russian Far East); Dal’nauka Press: Vladivostok, Russia, 1996; p. 364. (In Russian) [Google Scholar]
- Barinova, S.S.; Medvedeva, L.A.; Anisimova, O.V. Algae as Indicators of Environmental Quality; Institute Nature Conservation Press: Moscow, Russia, 2000; p. 150. (In Russian) [Google Scholar]
- Barinova, S.S.; Medvedeva, L.A.; Anisimova, O.V. Biodiversity of Environmental Indicator Algae; Pilies Studio: Tel Aviv, Israel, 2006; p. 498. (In Russian) [Google Scholar]
- Loseva, E.I.; Stenina, A.S.; Marchenko-Vagapova, T.I. Cadastre of the Fossil and Recent Diatoms from Northeastern Europe; Geoprint: Syktyvkar, Russia, 2004; p. 160. (In Russian) [Google Scholar]
- Stenina, A.S.; Sterlyagova, I.N. Bacillariophyta in epilithon of the Shchugor River (Urals, Komi Republic). Bot. Zhurnal 2017, 102, 1107–1122. [Google Scholar] [CrossRef]
- Stenina, A.S. Annotated list of Bacillariophyta of the Shchugor River (Urals, Komi Republic). Bot. Zhurnal 2019, 104, 41–57. [Google Scholar] [CrossRef]
- Gavshin, V.M.; Arkhipov, S.A.; Bobrov, V.A.; Melgunov, M.S.; Makarova, I.V.; Sukhorukov, F.V.; Stepin, A.S.; Khlystov, O.M.; Kireev, A.D.; Budashkina, V.V. Distribution of natural radioactive elements in Holocene–Pleistocene deep-water sediments of Lake Baikal and paleoclimate reconstruction. Geol. Geophys. 1998, 39, 1045–1058. [Google Scholar]
- Korotkiy, A.M.; Grebennikova, T.A.; Pushkar, B.C.; Razzhigaeva, N.G.; Volkov, V.G.; Ganzei, L.A.; Mokhova, L.M.; Bazarova, V.B.; Makarova, T.R. Climatic Changes in the South of the Far East in the Late Cenozoic (Miocene-Pleistocene); Publishing House of the Far Eastern State University: Vladivostok, Russia, 1996; p. 56. (In Russian) [Google Scholar]
- Lavrushin, Y.A.; Alekseev, M.N. Late Cenozoic paleoclimatic events in the Arctic ocean. Stratigraphy. Geol. Correl. 1993, 1, 48–58. [Google Scholar]
- Pushkar, V.S.; Cherepanova, M.V. Diatoms of the Pliocene and Anthropogen of the Northern Pacific (Stratigraphy and Paleoecology); Dal’nauka: Vladivostok, Russia, 2001; p. 226. (In Russian) [Google Scholar]
- Baskakova, L.A. Eventfulness and stages of Geological development in the Cenozoic of the Mainland part of the East of Russia. Pac. Geol. 2006, 25, 3–18. [Google Scholar]
- Zabelina, M.M.; Kiselev, I.A.; Proshkina-Lavrenko, A.I.; Sheshukova, V.S. Diatoms Keys to Freshwater Algae of the USSR; Soviet Science: Moscow, Russia, 1951; p. 619. (In Russian) [Google Scholar]
- Popovskaya, G.I.; Genkal, S.I.; Likhoshway, Y.V. Diatoms of the Plankton of Lake Baikal: Atlas and Key; Nauka: Novosibirsk, Russia, 2016; p. 180. (In Russian) [Google Scholar]
- Khursevich, G.K.; Karabanov, E.B.; Prokopenko, A.A.; Williams, D.F.; Kuz’min, M.I.; Fedenya, S.A.; Gvozdkov, A.N.; Kerber, E.V. Detailed diatom biostratigraphy of Baikal sediments during the brunhes chron and climatic factors of species formation. Russ. Geol. Geophys. 2001, 42, 108–129. [Google Scholar]
- Babanazarova, O.V.; Likhoshway, Y.V.; Sherbakov, D.Y. On the morphological variability of Aulacoseira baicalensis and Aulacoseira islandica (Bacillariophyta) of Lake Baikal, Russia. Phycologia 1996, 35, 113–123. [Google Scholar] [CrossRef]
- Endrikhinsky, A.S.; Cheremisinova, E.A. On the finding of Miocene deposits on the Vitim Plateau. Dokl. Acad. Sci. USSR 1970, 1, 885–888. [Google Scholar]
- Chernyaeva, G.P.; Lyamina, N.A.; Rasskazov, S.V. Biostratigraphy and conditions of accumulation of the Middle and Upper Miocene volcanogenic-sedimentary strata of the Dzhilinda depression in western Transbaikalia. Geol. Geophys. 2007, 48, 460–471. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
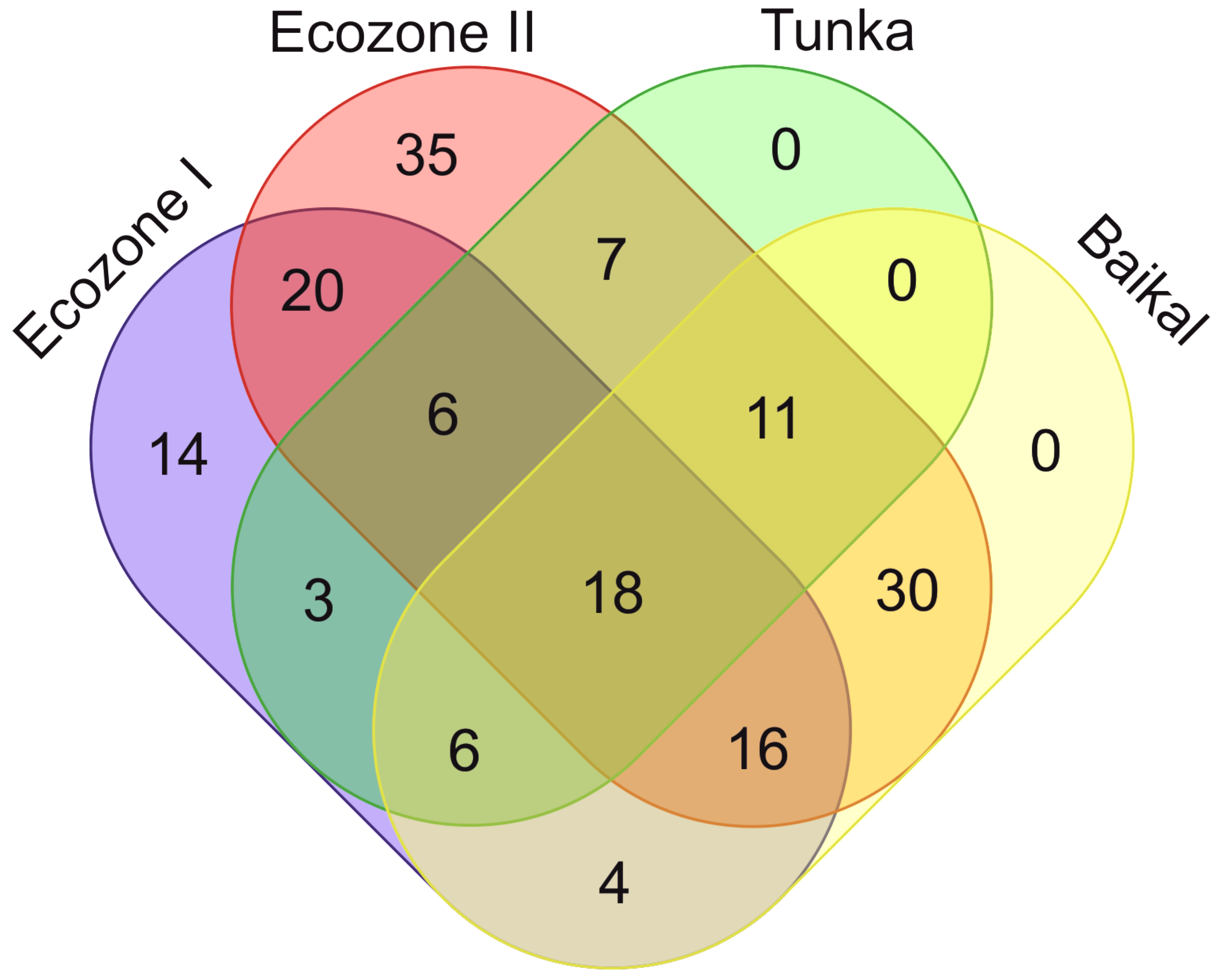{kind=link}
| № | Taxon | Diatom Ecozone | Ecological and Geographical Characteristics of Indicator Organisms | Fossil & Recent | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | Нab | Т | R | S1 | S2 | Sal | рН | А | Geo | |||
| Phylum Bacillariophyta Class Bacillariophyceae Subclass Bacillariophycidae Order Bacillariales Family Bacillariaceae | |||||||||||||
| 1. | Hantzschia amphioxys (Ehrenberg) Grunow | - | + | B | - | - | - | β-o | i | - | ind | k | R |
| 2. | Nitzschia alpina Hustedt | - | + | B | - | - | sx | o | - | - | acf | b | R |
| 3. | N. fonticola (Grunow) Grunow | - | + | B | - | - | - | o-β | i | 7.7 | alf | k | R |
| 4. | N. frustulum (Kützing) Grunow | - | + | B | - | - | - | - | hl | - | - | b | R |
| 5. | N. recta Hantzsch ex Rabenhorst | - | + | B | st | es | x | i | 6–9 | alf | k | R | |
| Order Cocconeidales Family Achnanthidiaceae | |||||||||||||
| 6. | Achnanthidium lineare W. Smith | - | + | B | - | - | - | x-o | i | - | ind | k | R |
| 7. | A. minutissimum (Kützing) Czarnecki | + | + | B | eterm | st-str | es | β | i | 4.3–9.2 | alf | k | R |
| 8. | A. obliquum Mereschkowsky | + | + | B | - | - | - | - | - | - | - | - | R |
| 9. | Eucocconeis flexella (Kützing) Meister | - | + | B | - | - | sx | o | mh | - | ind | a-a | R |
| 10. | E. laevis (Østrup) Lange-Bertalot | + | - | B | - | - | - | o | - | - | - | - | R |
| 11. | Planothidium dubium (Grunow) Round & Bukhtiyarova | - | + | B | - | - | sx | - | i | - | alf | - | R |
| 12. | P. ellipticum (Cleve) M.B. Edlund | + | - | B | - | str | sx | - | i | - | alf | k | R |
| 13. | P. frequentissimum (Lange-Bertalot) Lange-Bertalot | - | + | B | - | - | - | x-o | oh | - | alf | k | R |
| 14. | P. haynaldii (Schaarschmidt) Lange-Bertalot | + | - | B | - | - | sx | β-α | - | - | alf | k | R |
| 15. | P. joursacense (Héribaud) Lange-Bertalot | + | + | B | - | - | sx | - | - | - | - | - | R |
| 16. | P. lanceolatum (Brébisson ex Kützing) Lange-Bertalot | + | + | P-B | warm | st-str | sx | x-o | i | 7.5–8.1 | alf | k | R |
| 17. | P. linkei (Hustedt) Lange-Bertalot | + | + | B | - | - | - | - | hl | - | - | - | R |
| 18. | P. rostratum (Østrup) Lange-Bertalot | - | + | B | - | - | sx | - | - | - | - | - | R |
| 19. | Psammothidium marginulatum (Grunow) Bukhtiyarova & Round | - | + | B | - | st-str | sx | β-o | - | 5.4 | acf | a,k | R |
| 20. | Gololobovia obliqua (W. Gregory) Kulikovskiy, Glushchenko & Kociolek | - | + | B | - | - | - | - | - | - | - | - | R |
| 21. | Skabitschewskia oestrupii (A. Cleve) Kulikovskiy & Lange-Bertalot | - | + | B | - | - | - | o | i | - | ind | a-a | R |
| 22. | S. peragalloi (Brun & Héribaud) Kulikovskiy & Lange-Bertalot | - | + | B | - | - | sx | o | i | - | ind | b | R |
| Family Cocconeidaceae | |||||||||||||
| 23. | Cocconeis placentula Ehrenberg | + | + | P-B | temp | st-str | es | o-β | i | 5.5–9 | alf | k | R |
| 24. | Brebissonia lanceolata (C.Agardh) R.K. Mahoney & Reimer | - | + | B | - | - | - | - | - | - | - | - | R |
| 25. | Cymbella aff. laevis Nägeli | + | - | B | cool | - | sx | - | i | - | ind | b | R |
| 26. | C. amplificata Krammer | - | + | B | - | - | - | - | i | - | - | b | R |
| 27. | C. cistula (Ehrenberg) O. Kirchner | - | + | B | - | st-str | sx | o-β | i | 8 | alf | k | R |
| 28. | C. cymbiformis C. Agardh | - | + | B | temp | - | sx | - | i | 6.2–9 | alf | k | R |
| 29. | C. stuxbergii (Cleve) Cleve | - | + | B | - | - | - | - | - | - | - | - | R |
| 30. | Cymbopleura cuspidata (Kützing) Krammer | + | + | B | temp | - | - | o-α | i | 6.7 | ind | k | R |
| 31. | C. incerta (Grunow) Krammer | - | + | B | - | - | - | - | i | - | ind | a-a | R |
| 32. | C. subcuspidata (Krammer) Krammer | - | + | B | - | - | - | o | i | - | ind | a-a | R |
| 33. | Didymosphenia geminata (Lyngbye) Mart. Schmidt | - | + | B | - | st-str | sx | x | i | - | ind | a-a | R |
| 34. | Gomphonella olivacea (Hornemann) Rabenhorst | + | + | B | - | - | es | β-α | i | 7.5–8 | alf | k | R |
| Order Cymbellales Family Anomoeoneidaceae | |||||||||||||
| 35. | Adlafia minuscula var. muralis (Grunow) Lange-Bertalot | - | + | B | - | - | sp | o-β | i | - | ind | k | R |
| Family Gomphonemataceae | |||||||||||||
| 36. | Encyonema minutum (Hilse) D.G. Mann | - | + | B | - | st-str | es | o-β | oh | 6.2 | ind | k | R |
| 37. | E. silesiacum (Bleisch) D.G. Mann | - | + | B | - | st-str | sx | x-o | i | 6.2–7.7 | ind | k | R |
| 38. | Encyonopsis cesatii (Rabenhorst) Krammer | - | + | B | - | - | - | x-o | - | - | - | - | R |
| 39. | Gomphonema angustatum (Kützing) Rabenhorst | + | + | P-B | - | - | - | β | i | - | alf | k | R |
| 40. | G. intricatum Kützing | + | + | P-B | - | - | - | x-o | - | - | ind | k | R |
| 41. | G. acuminatum Ehrenberg | - | + | P-B | - | - | x-β | i | - | alf | k | R | |
| 42. | G. olivaceum var. minutissimum Hustedt | + | + | B | - | - | - | - | i | - | alf | b | R |
| 43. | G. sphaerophorum Ehrenberg | + | + | B | - | - | - | o | i | - | alf | b | R |
| 44. | G. ventricosum W. Gregory | - | + | P | cool | - | - | o-x | i | - | ind | k | R |
| 45. | Placoneis exigua (W. Gregory) Mereschkovsky | - | + | B | - | - | es | x-o | i | - | alf | k | R |
| 46. | P. gastrum (Ehrenberg) Mereschkowsky | + | + | B | - | - | sx | x-o | i | - | ind | k | R |
| 47. | Reimeria sinuata (W. Gregory) Kociolek & Stoermer | - | + | B | - | st | sx | - | i | - | ind | k | R |
| Family Rhoicospheniaceae | |||||||||||||
| 48. | Gomphosphenia grovei var. lingulata (Hustedt) Lange-Bertalot | + | + | B | - | str | es | β-α | i | - | - | k | F |
| Order Eunotiales Family Eunotiaceae | |||||||||||||
| 49. | Eunotia aff. exigua (Brébisson ex Kützing) Rabenhorst | - | + | B | - | - | es | o-β | hb | 3.4–8 | acf | k | R |
| 50. | E. aff. spatulata J. Veselá& J.R. Johansen | - | + | B | - | - | - | - | - | - | - | - | R |
| 51. | E. arcus Ehrenberg | - | + | B | - | - | - | x-β | i | - | acf | k | R |
| 52. | E. bidens Ehrenberg | + | - | B | cool | - | - | - | hb | - | acf | k | R |
| 53. | E. incisa W. Smith ex W. Gregory | - | + | B | - | str | es | α-β | - | 5.0 | acf | k | R |
| 54. | E. minor (Kützing) Grunow | + | - | B | - | - | es | x | i | 5.2 | acf | k | R |
| 55. | E. pectinalis (Kützing) Rabenhorst | - | + | B | temp | - | - | x, x-o, x-β | hb | 5.8–7.0 | ind | k | R |
| 56. | E. polydentula Hustedt | - | + | B | - | - | - | x-β | hb | - | acf | k | R |
| 57. | E. praerupta Ehrenberg | - | + | P-B | cool | st-str | sx | β | - | - | acf | k | R |
| 58. | E. robusta Ralfs | + | + | B | - | - | - | o | - | - | - | - | R |
| 59. | E. tenella (Grunow) Hustedt | + | + | B | - | str | es | o-β | hb | 5.1 | acf | a-a | R |
| 60. | E. veneris (Kützing) De Toni | - | + | B | - | - | - | β-o | hb | - | acf | k | R |
| Order Fragilariales Family Fragilariaceae | |||||||||||||
| 61. | Fragilaria aff. pararumpens Lange-Bertalot, G. Hofmann & Werum | - | + | P-B | - | - | - | - | - | - | - | - | R |
| 62. | F. capucina Desmazières | + | + | B | - | - | es | o | i | 7.7 | alf | k | R |
| 63. | F. radians (Kützing) D.M. Williams & Round | - | + | B | - | st-str | sx | - | - | - | - | Ha, Pt | R |
| 64. | F. aequalis Heiberg | - | + | B | - | - | - | - | - | - | - | - | R |
| 65. | F. vaucheriae (Kützing) J.B. Petersen | - | + | P-B | - | - | sx | o-β | i | 7.8 | alf | k | R |
| 66. | Fragilariforma constricta (Ehrenberg) D.M. Williams & Round | - | + | B | - | - | - | - | i | 5.2 | acf | a-a | R |
| 67. | F. virescens (Ralfs) D.M. Williams & Round | - | + | P-B | - | st | es | o | i | 6.8 | ind | k | R |
| 68. | Odontidium hyemale (Roth) Kützing | - | + | P-B | cool | st-str | sx | β-o | hb | 6.5–7.5 | ind | k | R |
| 69. | O. mesodon (Kützing) Kützing | + | + | B | cool | st-str | sx | o-β | hb | - | - | k | R |
| 70. | Punctastriata lancettula (Schumann) P.B. Hamilton & Siver | + | + | B | cool | - | es | o | i | 7.8 | alb | b | R |
| Family Staurosiraceae | |||||||||||||
| 71. | Pseudostaurosira brevistriata (Grunow) D.M. Williams & Round | + | + | P-B | - | st-str | - | x-o | i | 7.2 | alf | k | R |
| 72. | P. elliptica (Schumann) Edlund, E. Morales & Spaulding | + | + | B | - | - | - | β-α | - | - | - | k | R |
| 73. | P. parasitica (W. Smith) E. Morales | - | + | B | - | - | - | β-α | - | - | - | - | R |
| 74. | Staurosira aff. leptostauron (Ehrenberg) Kulikovskiy & Genkal | + | + | B | - | - | - | - | - | - | - | - | R |
| 75. | S. venter (Ehrenberg) Cleve & J.D. Möller | + | - | P-B | warm | st-str | sx | β | i | 5.5–9 | alf | k | R |
| 76. | S. binodis (Ehrenberg) Lange-Bertalot | + | + | B | - | - | - | o-β | i | - | alf | k | R |
| 77. | S. construens Ehrenberg | + | - | P-B | temp | st-str | sx | o | i | 5.5–9 | alf | k | R |
| 78. | S. construens var. triundulata (Reichelt) Bukhtiyarova | + | + | B | - | - | - | - | i | - | alf | k | R |
| 79. | S. leptostauron (Ehrenberg) Kulikovskiy & Genkal | + | + | B | - | st | es | α-β | hb | 8.4 | alf | b | R |
| 80. | S. subsalina (Hustedt) Lange-Bertalot | - | + | P-B | - | st | es | o | hl | - | alf | k | R |
| 81. | S. tabellaria (W. Smith) Leuduger-Fortmorel | + | - | B | - | - | - | - | - | - | - | - | R |
| 82. | Staurosirella lanceolata (Hustedt) E. Morales, C.Wetzel & L.Ector | + | + | B | - | - | - | - | - | - | - | - | R |
| 83. | S. martyi (Héribaud) E. Morales & K.M. Manoylov | + | + | P-B | - | st-str | es | o-α | i | 7.5–9 | alf | k | R |
| 84. | S.pinnata (Ehrenberg) D.M. Williams & Round | + | + | B | temp | st-str | es | β-α | hl | 6.2–9.3 | alf | k | R |
| Order Licmophorales Family Ulnariaceae | |||||||||||||
| 85. | Hannaea arcus (Ehrenberg) R.M. Patrick | - | + | B | str | es | o | i | - | alf | a-a | R | |
| 86. | H. baicalensis Genkal, Popovskaya & Kulikovskiy | - | + | P | - | - | - | - | - | - | - | - | R |
| 87. | Ulnaria acus (Kützing) Aboal | - | + | P | - | st-str | es | o-α | i | - | alb | k | R |
| 88. | U. contracta (Østrup) E. Morales & M.L. Vis | - | + | P | - | - | es | - | - | - | - | - | R |
| 89. | U. ulna (Nitzsch) Compère | + | + | P-B | - | - | - | o-α | i | - | alf | k | R |
| Order Naviculales Family Amphipleuraceae | |||||||||||||
| 90. | Frustulia vulgaris (Thwaites) De Toni | - | + | P-B | - | st | es | x-β | i | - | alf | k | R |
| Family Brachysiraceae | |||||||||||||
| 91. | Nupela impexiformis (Lange-Bertalot) Lange-Bertalot | - | + | B | - | - | es | - | - | 7.0 | ind | - | R |
| Family Cavinulaceae | |||||||||||||
| 92. | Cavinula cocconeiformis (W.Gregory ex Greville) D.G. Mann & A.J. Stickle | + | + | P-B | - | str | es | o | i | 6.9 | ind | a-a | R |
| 93. | C. jaernefeltii (Hustedt) D.G. Mann & A.J. Stickle | + | + | B | - | - | - | o | i | - | acf | k | R |
| 94. | C. pseudoscutiformis (Hustedt) D.G. Mann & Stickle | - | + | P-B | - | st-str | sx | o | i | 6.7 | ind | a-a | R |
| 95. | C. scutelloides (W. Smith) Lange-Bertalot | + | + | B | - | - | - | o-β | - | - | - | - | R |
| 96. | C. scutiformis (Grunow) D.G. Mann & A.J. Stickle | + | + | B | - | - | - | - | i | - | ind | a-a | R |
| Family Diploneidaceae | |||||||||||||
| 97. | Diploneis elliptica (Kützing) Cleve | + | - | B | temp | sx | o-a | i | - | alf | k | R | |
| 98. | D. ovalis (Hilse) Cleve | + | + | B | - | - | sp | β | i | 6.5–9 | alb | b | R |
| 99. | D. parma Cleve | + | - | B | cool | - | - | o-β | i | - | alf | Ha | R |
| Family Naviculaceae | |||||||||||||
| 100. | Caloneis bacillum (Grunow) Cleve | - | + | B | temp | - | es | o | i | - | alf | k | R |
| 101. | Gyrosigma acuminatum (Kützing) Rabenhorst | - | + | B | cool | - | - | o-x | i | - | alf | k | R |
| 102. | Hippodonta costulata (Grunow) Lange-Bertalot, Metzeltin & Witkowski | + | - | B | temp | - | sx | - | hl | - | alf | b | R |
| 103. | H. coxiae Lange-Bertalot | - | + | B | - | - | - | - | - | - | alf | - | R |
| 104. | Navicula cryptocephala Kützing | + | + | P-B | - | - | - | x | i | - | alf | k | R |
| 105. | N. cryptotenella Lange-Bertalot in Krammer & Lange-Bertalot | - | + | P-B | - | - | - | b-a | i | - | alf | k | R |
| 106. | N. gregaria Donkin | + | - | B | - | - | es | x-β | mh | - | alf | k | R |
| 107. | N. johncarteri D.M. Williams in D.M. Williams & G. Reid | + | + | B | - | - | sx | x | - | - | - | - | R |
| 108. | N. menisculus Schumann | - | + | B | - | - | es | x-β | i | - | alf | k | R |
| 109. | N. meniscula var. muralis (Grunow) Lange Bertalot | - | + | B | R | ||||||||
| 110. | N. radiosa Kützing | + | + | B | temp | st-str | es | o | i | 5.-9 | ind | k | R |
| 111. | N. rhynchocephala Kützing | - | + | B | - | - | - | β | hl | 7.3–7.8 | alf | k | R |
| 112. | N. viridula (Kützing) Ehrenberg | + | - | B | - | - | es | o | hl | - | alf | k | R |
| Family Neidiaceae | |||||||||||||
| 113. | Neidium baicalense Jasnitsky | - | + | B | - | - | - | - | i | - | ind | - | R |
| 114. | N. bisulcatum (Lagerstedt) Cleve | - | + | B | - | - | es | o-β | hb | 5.2 | ind | b | R |
| 115. | N. dubium (Ehenberg) Cleve | - | + | B | - | - | - | x | i | - | alf | k | R |
| 116. | N. longiceps (W. Gregory) R. Ross | - | + | B | - | - | sp | o | i | 6.1 | acf | a-a | R |
| Family Sellaphoraceae | |||||||||||||
| 117. | Eolimna aboensis (Cleve) S.I. Genkal | + | + | B | - | - | - | - | i | - | ind | a-a | R |
| 118. | E. minima (Grunow) Lange-Bertalot | + | + | B | - | - | - | o-β | - | - | - | - | R |
| 119. | Sellaphora bacillum (Ehrenberg) D.G. Mann | + | - | B | - | st-str | sx | x-o | i | 7.-9 | alf | k | R |
| 120. | S. medioconvexa (Hustedt) C.E. Wetzel | + | - | B | - | - | - | - | hb | - | - | b | R |
| 121. | S. pupula (Kützing) Mereschkovsky | - | + | B | eterm | st | sp | o-x | hl | 5.2–9 | ind | k | R |
| 122. | S. stauroneioides (Lange-Bertalot) Veselá & J.R.Johansen | + | - | B | - | - | - | - | - | - | - | - | R |
| Family Stauroneidaceae | |||||||||||||
| 123. | Craticula subminuscula (Manguin) C.E. Wetzel & Ector | - | + | P-B | - | - | sp | α-β | - | - | - | - | R |
| 124. | Stauroneis anceps Ehrenberg | + | + | P-B | - | - | - | x | i | - | ind | k | R |
| 125. | S. phoenicenteron (Nitzsch) Ehrenberg | - | + | B | temp | - | es | x-o | i | 7.3 | ind | k | R |
| 126. | S. smithii Grunow | + | + | P-B | - | st-str | - | x-o | - | - | alf | k | R |
| Family Pinnulariaceae | |||||||||||||
| 127. | Pinnularia abaujensis var. linearis (Hustedt) R.M. Patrick | + | - | B | - | - | - | - | i | - | ind | b | R |
| 128. | P.aff. interrupta W. Smith | - | + | B | - | - | sp | β-o | i | 5.6 | acf | k | R |
| 129. | P. brebissonii (Kützing) Rabenhorst | + | - | B | - | - | es | o-β | - | - | - | k | R |
| 130. | P. eifeliana (Krammer) Krammer | - | + | B | - | - | - | o | - | - | - | - | R |
| 131. | P. interrupta var. minutissima Hustedt | - | + | B | - | - | - | - | i | - | - | b | R |
| 132. | P. microstauron (Ehrenberg) Cleve | + | + | B | temp | - | sp | x | i | - | ind | k | R |
| 133. | P.biundulata (O.Müller) Kulikovskiy & Genkal | - | + | B | - | - | - | - | - | - | - | - | R |
| 134. | P. nodosa (Ehrenberg) W. Smith | - | + | B | - | - | - | o | i | - | ind | a-a | R |
| 135. | P. schroederi (Hustedt) Cholnoky | + | - | B | - | - | es | - | - | - | - | - | R |
| 136. | P. sinistra Krammer | + | - | B | - | - | - | o | hb | - | acf | k | R |
| 137. | P. subcapitata W. Gregory | - | + | B | - | - | sp | x-o | i | 6.1 | ind | k | R |
| 138. | P. viridis (Nitzsch) Ehrenberg | + | + | P-B | temp | - | es | o-x | i | 7.1 | ind | k | R |
| Order Rhopalodiales Family Rhopalodiaceae | |||||||||||||
| 139. | Epithemia adnata (Kützing) Brébisson | - | + | B | temp | st | sx | β-α | i | 5.5–9 | alb | k | R |
| 140. | E. gibba (Ehrenberg) Kützing | + | + | B | temp | - | es | x-o | i | 6.2–9 | alb | k | R |
| 141. | E. sorex Kützing | - | + | B | temp | st | sx | o-α | i | 5.-9 | alf | k | R |
| Order Surirellales Family Surirellaceae | |||||||||||||
| 142. | Iconella linearis (W. Smith) Ruck & Nakov | - | + | P-B | - | - | es | o-β | i | 5.-9 | ind | Ha | R |
| 143. | Surirella librile (Ehrenberg) Ehrenberg | + | + | B | - | - | - | - | - | - | - | - | R |
| Order Tabellariales Family Tabellariaceae | |||||||||||||
| 144. | Diatoma moniliformis subsp. ovalis (F. Fricke) Lange-Bertalot, Rumrich & G. Hofmann | - | + | B | - | - | - | β-α | - | - | - | - | |
| 145. | D. tenuis C. Agardh | - | + | P-B | - | - | sx | - | hl | - | ind | k | R |
| 146. | D. vulgaris Bory | - | + | P-B | - | st-str | sx | β-α | i | 6.2–7.5 | ind | k | R |
| 147. | Meridion circulare (Greville) C. Agardh | + | + | B | - | str | es | o-β | hb | 8.0 | alf | k | R |
| 148. | Tabellaria fenestrata (Lyngbye) Kützing | + | + | P-B | - | - | - | x | - | - | ind | k | R |
| 149. | T. flocculosa (Roth) Kützing | + | + | P-B | eterm | st-str | es | o-α | hb | 5.-7 | acf | a,k | R |
| 150. | Tetracyclus ellippticus var. latissimus Hustedt | + | - | B | - | - | - | - | - | - | - | - | F |
| 151. | T. emarginatus (Ehrenberg) W. Smith | + | + | P-B | cool | st-str | - | - | i | - | acf | a-a | F |
| 152. | T. glans (Ehrenberg) F.W. Mills | + | + | B | - | - | - | o | i | - | acf | a-a | R |
| 153. | T. lapponicus Tynni | + | + | B | - | - | - | - | - | - | - | - | F |
| Order Thalassiophysales Family Catenulaceae | |||||||||||||
| 154. | Amphora aequalis Krammer | - | + | B | - | - | - | - | - | - | - | - | R |
| 155. | A.aff. inariensis Krammer | + | - | B | - | - | - | o-β | oh | - | alf | Ha | R |
| 156. | A. lybica Ehrenberg | - | + | B | - | - | es | - | hl | - | alf | k | R |
| 157. | A. ovalis (Kützing) Kützing | + | - | B | temp | st-str | sx | α-β | i | 6.2–9 | alf | k | R |
| 158. | A. pediculus (Kützing) Grunow | - | + | B | temp | st | es | o-α | i | - | alf | k | R |
| Class Coscinodiscophyceae Order Aulacoseirales Family Aulacoseiraceae | |||||||||||||
| 159. | Aulacoseira aff. baicalensis (Wislouch) Simonsen | + | + | P | - | - | - | - | - | - | - | - | R |
| 160. | A. ambigua (Grunow) Simonsen | + | + | P | - | st-st | sp | α-β | i | 6–8.5 | alf | k | R |
| 161. | A.ambigua f. curvata (Skabichevskij) Genkal | + | + | P | - | st-str | sp | o-β | i | 7.1 | alb | k | R |
| 162. | A. distans (Ehrenberg) Simonsen | + | + | P-B | cool | - | sp | x-o | i | 6.9 | acf | b | R |
| 163. | A. islandica (O. Müller) Simonsen | + | - | P | cool | - | es | o-x | i | - | acf | b | R |
| 164. | A. subarctica (O. Müller) E.Y. Haworth | + | - | P | - | st-str | - | α-β | i | 7.3 | alb | a, k | R |
| Order Melosirales Family Melosiraceae | |||||||||||||
| 165. | Melosira undulata (Ehrenberg) Kützing | + | + | P-B | temp | - | - | o-α | i | - | alb | k | R |
| Order Paraliales Family Radialiplicataceae | |||||||||||||
| 166. | Ellerbeckia arenaria f. teres (Brun) R.M. Crawford | + | + | P-B | - | st-str | - | o-α | i | - | alf | k | R |
| Class Mediophyceae Order Stephanodiscales Family Stephanodiscaceae | |||||||||||||
| 167. | Cyclotella tuncaica Nikiteeeva, Likhoshway & Pomazkina | + | + | P | - | - | - | - | - | - | - | - | F |
| 168. | Stephanodiscus tuncaensis Pomazkina & Likhoshway | + | + | P | - | - | - | - | - | - | - | - | F |
| Class Bacillariophyta classis incertae sedis Order Bacillariophyta ordo incertae sedis Family Bacillariophyta familia incertae sedis | |||||||||||||
| 169. | Gliwiczia calcar (Cleve) M. Kulikovskiy, Lange-Bertalot & A. Witkowski | + | + | B | - | - | - | o | i | - | alf | a-a | R |
| 170. | Navigeia decussis (Østrup) Bukhtiyarova | + | - | B | - | - | es | o-α | i | - | alf | b | R |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Titova, L.; Hassan, A.; Mikhailov, I.; Rodionova, E.; Rasskazov, S.; Usoltseva, М. Diversity and Ecology of Diatoms in Pliocene Deposits of the Tunka Valley (Baikal Rift Zone). Diversity 2021, 13, 479. https://doi.org/10.3390/d13100479
Titova L, Hassan A, Mikhailov I, Rodionova E, Rasskazov S, Usoltseva М. Diversity and Ecology of Diatoms in Pliocene Deposits of the Tunka Valley (Baikal Rift Zone). Diversity. 2021; 13(10):479. https://doi.org/10.3390/d13100479
Chicago/Turabian StyleTitova, Lyubov’, Abdulmonem Hassan, Ivan Mikhailov, Elena Rodionova, Sergei Rasskazov, and Мarina Usoltseva. 2021. "Diversity and Ecology of Diatoms in Pliocene Deposits of the Tunka Valley (Baikal Rift Zone)" Diversity 13, no. 10: 479. https://doi.org/10.3390/d13100479
APA StyleTitova, L., Hassan, A., Mikhailov, I., Rodionova, E., Rasskazov, S., & Usoltseva, М. (2021). Diversity and Ecology of Diatoms in Pliocene Deposits of the Tunka Valley (Baikal Rift Zone). Diversity, 13(10), 479. https://doi.org/10.3390/d13100479