
FloCan—A Revised Checklist for the Flora of the Canary Islands



Abstract
:1. Introduction
2. Materials and Methods
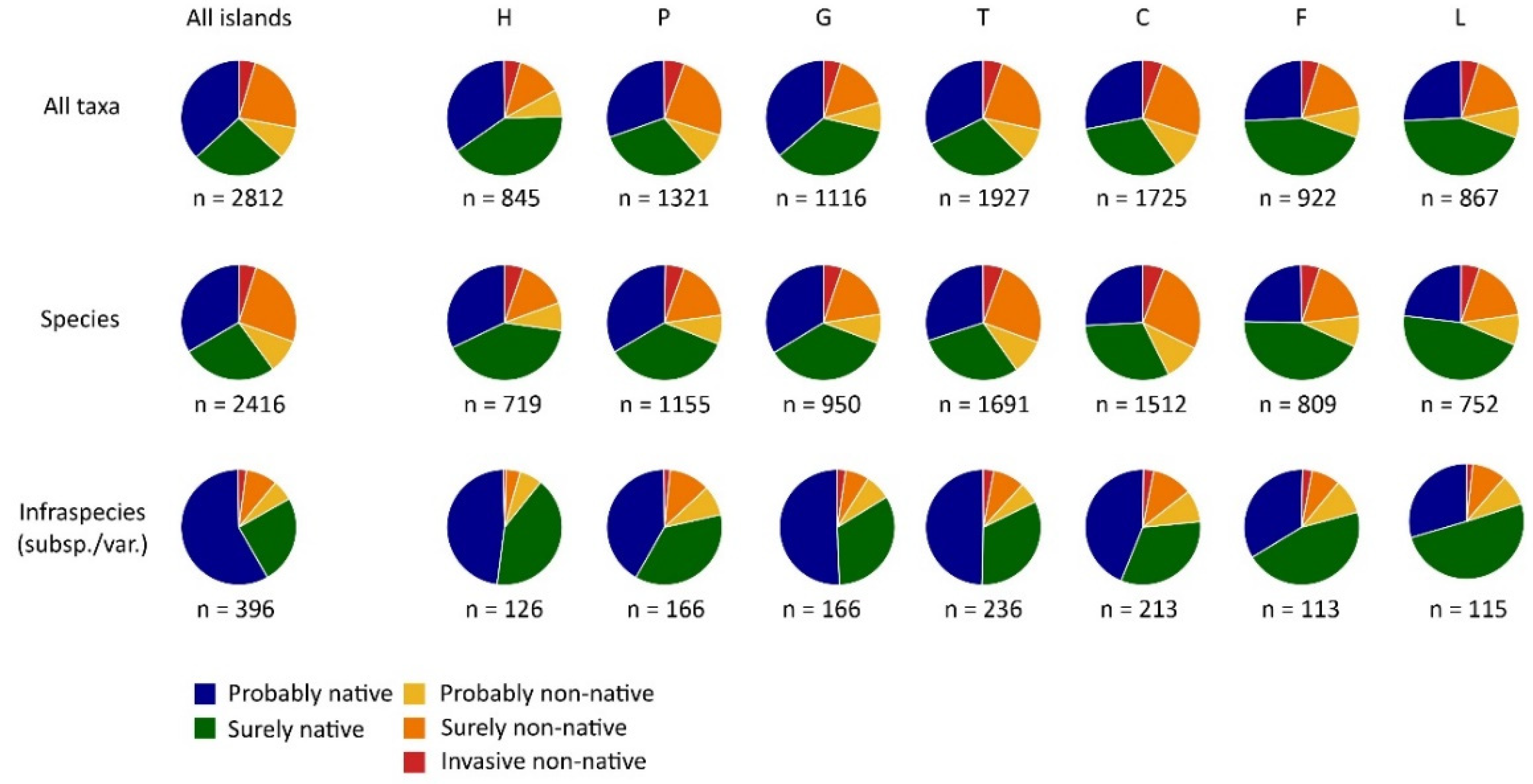
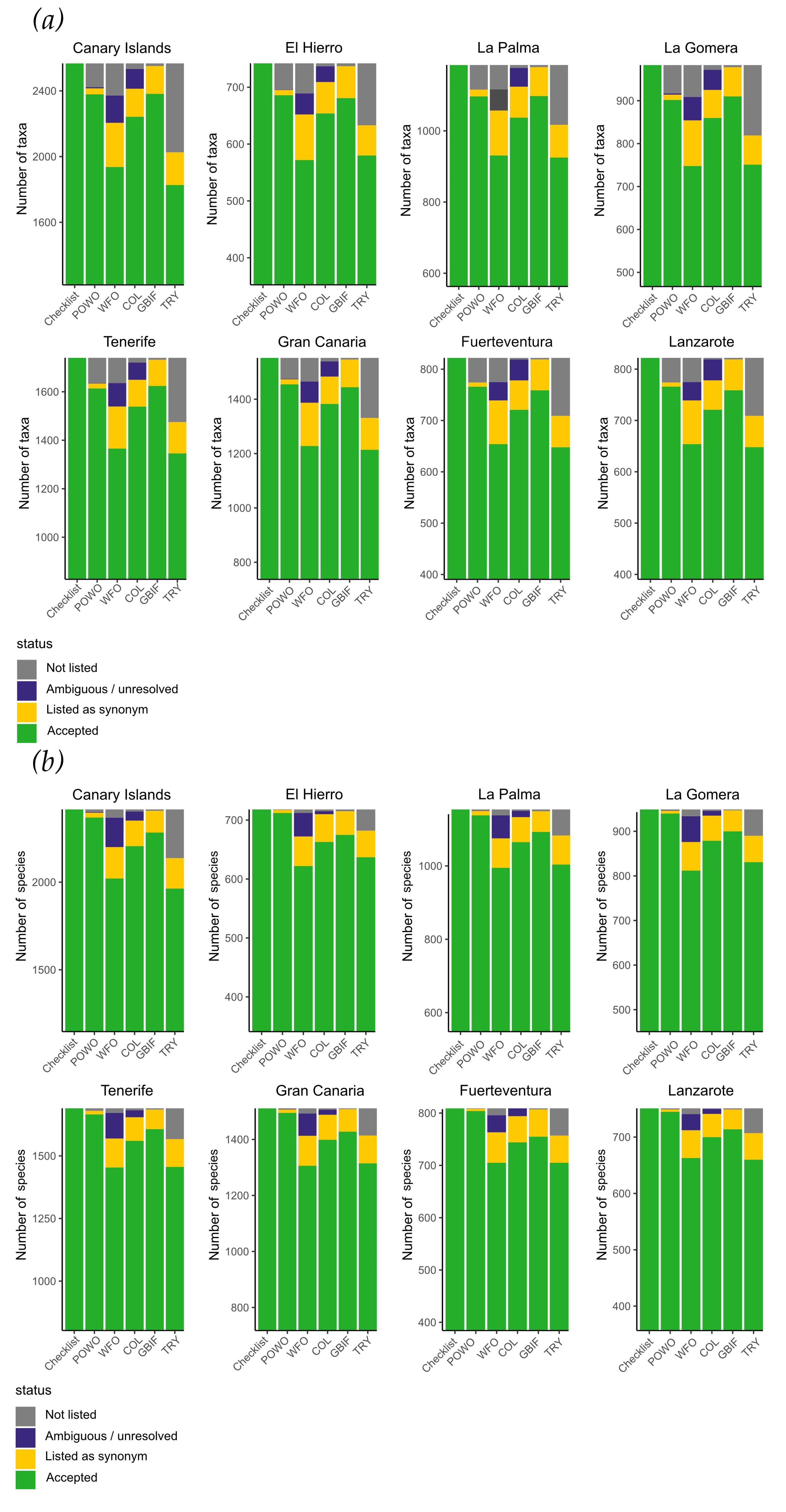
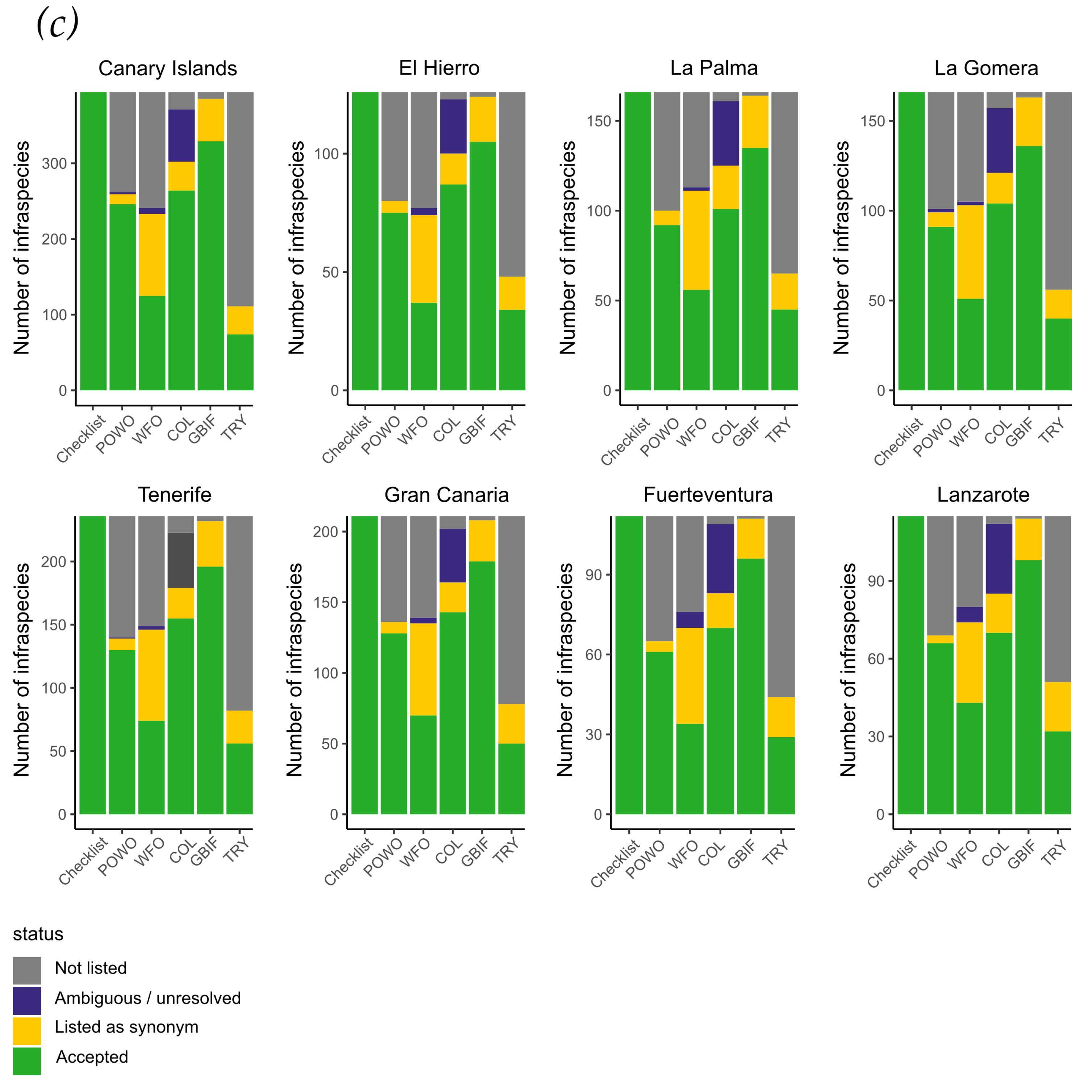
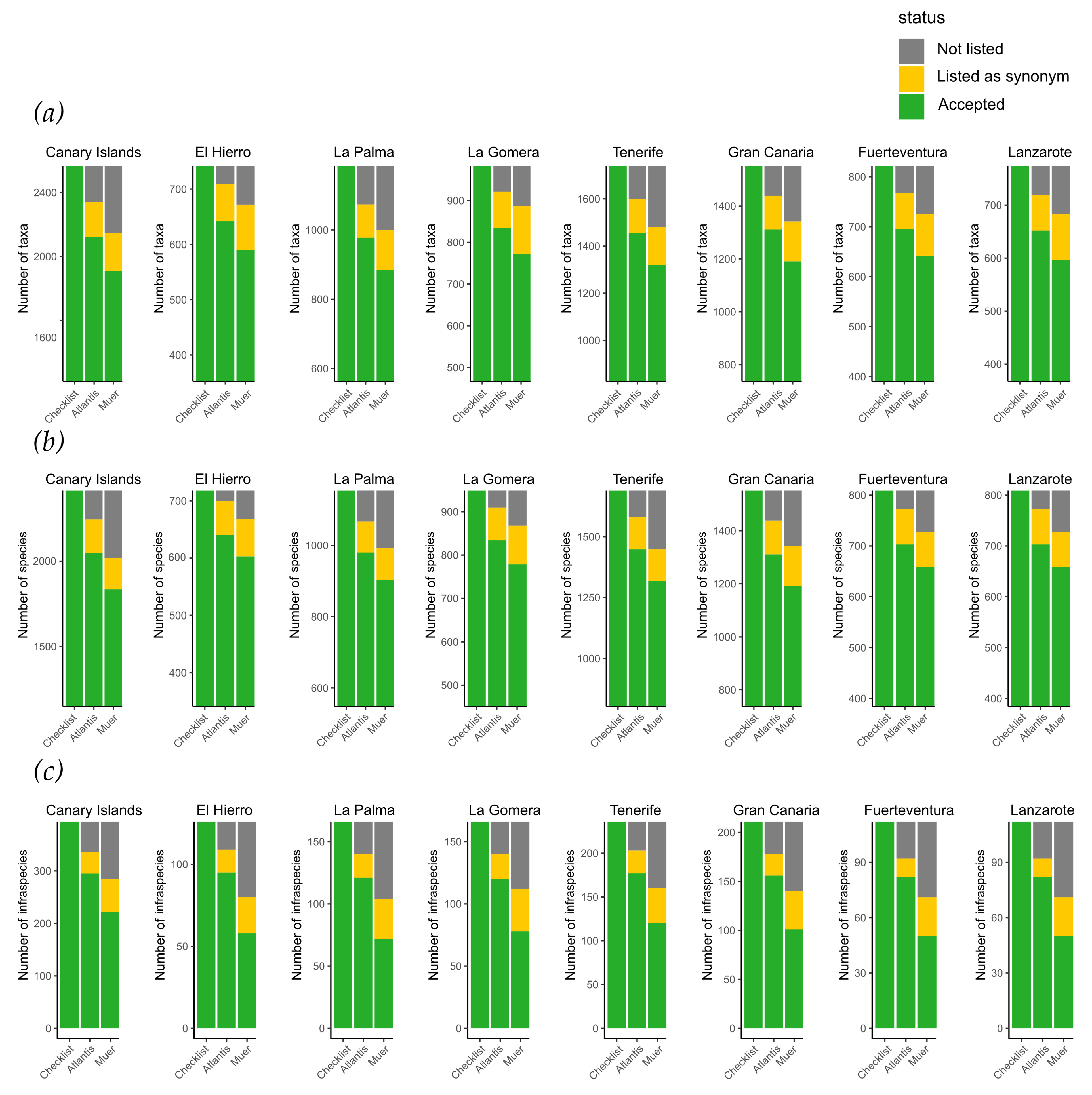
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Humboldt, A.V.; Bonpland, A. Viage á Las Regiones Equinocciales del Nuevo Continente; Casa de Rosa: Paris, France, 1826. [Google Scholar]
- Viera y Clavijo, D.J. Diccionario de Historia Natural de las Islas Canarias, o Índice Alfabético Descriptivo de sus Tres Reinos, Animal, Vegetal y Mineral; Excma. Mancomunidad de Cabildos de Las Palmas, Plan Cultural: Las Palmas de Gran Canaria, Spain, 1799. [Google Scholar]
- Darwin, C. Voyage of the Beagle. In The Narrative of the Voyages of H.M. Ships Adventure and Beagle; Henry Colburn: London, UK, 1839; Volume 3. [Google Scholar]
- Webb, P.B.; Berthelot, S. Histoire naturelle des Iles Canaries. In Phytogeographia Canariensis; Béthune: Paris, France, 1844; 1430p. [Google Scholar]
- Christ, H. Vegetation und Flora der Canarischen Inseln. Engler’s Bot. Jahrbücher 1885, 6, 458–526. [Google Scholar]
- Christ, H. Specilegium canariense. Engler’s Bot. Jahrbücher 1887, 9, 86–172. [Google Scholar]
- Pitard, J.; Proust, L. Les Iles Canaries. In Flore de l’Archipel; Béthune: Paris, France, 1908; 503p. [Google Scholar]
- Eriksson, O.; Hansen, A.; Sunding, P. Flora of Macaronesia: Checklist of Vascular Plants; Sommerfeltia: Umea, Sweden, 1974. [Google Scholar]
- Kunkel, G. Flora y Vegetación del Archipiélago Canario. Tratado Florístico. 2ª Parte. Dicotiledóneas; Ed. Edirca: Las Palmas de Gran Canaria, Spain, 1991; 312p. [Google Scholar]
- Kunkel, G. Die Kanarischen Inseln und ihre Pflanzenwelt; Gustav Fischer Verlag: Stuttgart, Jena, Germany; New York, NY, USA, 1993. [Google Scholar]
- Bramwell, D. Flora de las Islas Canarias; Rueda: Madid, Spain, 1997. [Google Scholar]
- Schönfelder, P.; Schönfelder, I. Die Kosmos-Kanarenflora; Kosmos: Stuttgart, Germany, 2012; 319p. [Google Scholar]
- Santos Guerra, A. Vegetacion y Flora de La Palma; Editorial Interinsular Canaria: Santa Cruz de Tenerife, Spain, 1983; 348p. [Google Scholar]
- Fernandez-Palacios, J.M.; Martin Esquivel, J.L. Naturaleza de las Islas Canarias; Editorial Interinsular Canaria: Santa Cruz de Tenerife, Spain, 2002. [Google Scholar]
- Pott, R.; Hüppe, J.; Wildpret de la Torre, W. Die Kanarischen Inseln—Natur- und Kulturlandschaften; Ulmer: Stuttgart, Germany, 2003. [Google Scholar]
- Montelongo, V.; Bramwell, D.; Fernández-Palacios, O. Parolinia glabriuscula (Brassicaceae), una nueva especie para Gran Canaria (Islas Canarias, España). Bot. Macaronésica 2003, 24, 67–72. [Google Scholar]
- Gil Gonzalez, J.; Gil Gonzales, M.L.; Morales Mateos, J.B.; Mesa Coello, R. Vicia vulcanorum (Fabaceae) a new species from the island of Lanzarote (Canary Islands). Collect. Bot. 2012, 31, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Bañares Baudet, Á.; Acevedo Rodríguez, A.; Rebolé Beaumont, Á. Monanthes subrosulata, a new species of M. sect. Sedoidea (Crassulaceae) from La Palma, Canary Islands, Spain. Willdenowia 2013, 43, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Crawford, D.J.; Mort, M.E.; Archibald, J.K. Tolpis santosii (Asteraceae, Chichorieae), a new species from La Palma, Canary Islands. Vieraea 2013, 41, 169–175. [Google Scholar]
- Gil Gonzalez, J.; Morales Mateos, J.B.; Gil Gonzales, M.L.; Mesa Coello, R. Vicia voggenreiteriana (Fabaceae) a new species from the island of La Gomera (Canary Islands). Viaeraea 2013, 41, 189–201. [Google Scholar]
- Friesen, N.; Herden, T.; Schönfelder, P. Allium canariense (Amaryllidaceae), a species endemic of the Canary Islands. Phytotaxa 2015, 221, 1–20. [Google Scholar] [CrossRef]
- Atlantis Biota Database. Available online: https://www.biodiversidadcanarias.es/biota/ (accessed on 5 July 2021).
- Mauchamp, A.; Atkinson, R. Rapid, Recent and Irreversible Habitat Loss: Scalesia Forest in the Galapagos Islands; Galapagos Report 2009–2010; GNPD; GCREG; CDF; GC: Galápagos, Ecuador, 2010; pp. 108–112. [Google Scholar]
- Wittmer, M.C. The dodo and the tambalacoque tree: An obligate mutualism reconsidered. Oikos 1991, 61, 133–137. [Google Scholar] [CrossRef] [Green Version]
- de Nascimento, L.; Willis, K.J.; Fernandez-Palacios, J.M.; Criado, C.; Whittaker, R.J. The long-term ecology of the lost forests of La Laguna, Tenerife (Canary Islands). J. Biogeogr. 2009, 36, 499–514. [Google Scholar] [CrossRef]
- Bañares Baudet, Á.; Marrero Gómez, M.V.; Scholz, S. Taxonomic and nomenclatural notes on Crassulaceae of the Canary Islands, Spain. Willdenowia 2008, 38, 475–489. [Google Scholar] [CrossRef] [Green Version]
- Peroni, A.; Peroni, G. Considerazioni tassonomiche sul genere Cystopteris Bernh. (Athyriaceae: Pteridophyta) di El Hierro (Isole Canarie). Ann. Mus. Cov. Rovereto Sez. Arch. St. Sc. Nat. 2006, 21, 207–210. [Google Scholar]
- Vitales, D.; Garnatje, T.; Pellicer, J.; Vallès, J.; Santos-Guerra, A.; Sanmartin, I. The explosive radiation of Cheirolophus (Asteraceae, Cardueae) in Macaronesia. BMC Evol. Biol. 2014, 14, 118. [Google Scholar] [CrossRef] [Green Version]
- Hansen, A.; Sunding, P. Flora of Macaronesia: Checklist of Vascular Plants, 4th ed.; Sommerfeltia: Umea, Sweden, 1993; Volume 17, pp. 1–295. [Google Scholar]
- Hohenester, A.; Welss, W. Exkursionsflora für die Kanarischen Inseln. Mit Ausblicken auf ganz Makaronesien; Eugen Ulmer: Stuttgart, Germany, 1993. [Google Scholar]
- Bramwell, D.; Bramwell, Z. Flores Silvestres de las Islas Canarias, 4th ed.; Rueda Ed.: Madrid, Spain, 2001; 437p. [Google Scholar]
- Izquierdo, I.; Martín, J.L.; Zurita, N.; Arechavaleta, M. (Eds.) Lista de especies silvestres de Canarias (hongos, plantas y animales terrestres). In Consejería de Medio Ambiente y Ordenación Territorial; Gobierno de Canarias: Las Palmas de Gran Canaria, Spain, 2004; 500p. [Google Scholar]
- Arechavaleta, M.; Rodriguez, S.; Zurita, N.; Garcia, A. Lista de Especies Silvestres de Canarias. Hongos, Plantas y Animales Terrestres; Gobierno de Canarias: Las Palmas de Gran Canaria, Spain, 2010; 579p. [Google Scholar]
- Muer, T.; Sauerbier, H.; Cabrera Calixto, F. Die Farn- und Blütenpflanzen der Kanarischen Inseln; Markgraf Publishers: Weikersheim, Germany, 2016. [Google Scholar]
- Flora de Canarias (Gil). Available online: http://www.floradecanarias.com/ (accessed on 5 July 2021).
- Freiberg, M.; Winter, M.; Gentile, A.; Zizka, A.; Muellner-Riehl, A.N.; Weigelt, A.; Wirth, C. LCVP, The Leipzig catalogue of vascular plants, a new taxonomic reference list for all known vascular plants. Sci. Data 2020, 7, 416. [Google Scholar] [CrossRef]
- Kier, G.; Kreft, H.; Lee, T.G.; Jetz, W.; Ibisch, P.L.; Nowicki, C.; Mutke, J.; Barthlott, W. A global assessment of endemism and species richness across island and mainland regions. Proc. Natl. Acad. Sci. USA 2009, 106, 9322–9327. [Google Scholar] [CrossRef] [Green Version]
- Steinbauer, M.; Beierkuhnlein, C. Characteristic pattern of species diversity on the Canary Islands. Erdkunde 2010, 64, 57–71. [Google Scholar] [CrossRef]
- Lenzner, B.; Weigelt, P.; Kreft, H.; Beierkuhnlein, C.; Steinbauer, M. The general dynamic model of island biogeography revisited at the level of major flowering plant families. J. Biogeogr. 2017, 44, 1029–1040. [Google Scholar] [CrossRef]
- Fernández-Palacios, J.M.; Otto, R.; Borregard, M.K.; Kreft, H.; Price, J.P.; Steinbauer, M.J.; Weigelt, P.; Whittaker, R.J. Evolutionary winners are ecological losers among oceanic island plants. J. Biogeogr. 2021, 48, 2186–2198. [Google Scholar] [CrossRef]
- Cutts, V.; Hanz, D.M.; Barjas-Barbosa, M.P.; Algar, A.C.; Steinbauer, M.J.; Irl, S.D.H.; Kreft, H.; Weigelt, P.; Fernandez-Palacios, J.M.; Field, R. Scientific floras can be reliable sources for some trait data in a system with poor coverage in global trait databases. J. Veg. Sci. 2021, 32, e12996. [Google Scholar] [CrossRef]
- Hortal, J.; Lobo, J.M.; Jiménez-Valverde, A. Limitations of Biodiversity Databases: Case Study onSeed-Plant Diversity in Tenerife, Canary Islands. Conserv. Biol. 2007, 21, 853–863. [Google Scholar] [CrossRef] [PubMed]
- Wikipedia Spanish on Categoría: Flora endémica de Canarias. Available online: https://es.wikipedia.org/wiki/Categor%C3%ADa:Flora_end%C3%A9mica_de_Canarias (accessed on 28 June 2021).
- Wikipedia English on Endemic Plants of the Canary Islands. Available online: https://en.wikipedia.org/wiki/Category:Endemic_flora_of_the_Canary_Islands (accessed on 28 June 2021).
- Hobohm, C.; Janišová, M.; Steinbauer, M.; Landi, S.; Field, R.; Vanderplank, S.; Beierkuhnlein, C.; Grytnes, J.A.; Vetaas, O.R.; Fidelis, A.; et al. Global endemics-area relationships of vascular plants. Perspect. Ecol. Conserv. 2019, 17, 41–49. [Google Scholar] [CrossRef]
- Hohbohm, C.; Moro-Richter, M.; Beierkuhnlein, C. Distribution and Habitat Affinity of Endemic and Threatened Species—Global Assessment. In Perspectives for Biodiversity and Ecosystems; Hobohm, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2021. [Google Scholar]
- Thompson, J.D. Plant Evolution in the Mediterranean: Insights for Conservation; Oxford University Press: Oxford, UK, 2020. [Google Scholar]
- Marreiro, A.; Almeida, R.S.; Gonzalez-Martin, M. A new species of wild dragon tree, Dracaena (Dracaenaceae) from Gran Canaria and its taxonomic and biogeographic implications. Bot. J. Linn. Soc. 1998, 128, 291–314. [Google Scholar]
- Rodríguez-Sánchez, F.; Guzmán, B.; Valido, A.; Vargas, P.; Arroyo, J. Late Neogene history of the laurel tree (Laurus, L.; Lauraceae) based on phylogeographical analyses of Mediterranean and Macaronesian populations. J. Biogeogr. 2009, 36, 1270–1281. [Google Scholar] [CrossRef] [Green Version]
- Pyšek, P.; Jarosik, V.; Hulme, P.E.; Hejda, M.; Schaffner, U.; Vila, M.A. Global assessment of invasive plant impacts on resident species, communities and ecosystems: The interaction of impact measures, invading species traits and environment. Glob. Chang. Biol. 2011, 18, 1725–1737. [Google Scholar] [CrossRef]
- Ríos-Mesa, D.; Pereira-Lorenzo, S.; González-Díaz, A.; Hernádez-González, J.; González-Diaz, E.; Saúco, V.G. The status of Chestnut cultivation and utilization in the Canary Islands. Adv. Hortic. Sci. 2011, 25, 90–98. [Google Scholar]
- Irl, S.; Schweiger, A.; Steinbauer, M.; Ah-Peng, C.; Arévalo, J.R.; Beierkuhnlein, C.; Chiarucci, A.; Daehler, C.C.; Fernandez-Palacios, J.M.; Flores, O.; et al. Human impact, climate and dispersal strategies determine plant invasion on islands. J. Biogeogr. 2021. early view. [Google Scholar] [CrossRef]
- Irl, S.; Steinbauer, M.; Messinger, J.; Blume-Werry, G.; Palomares-Martínez, A.; Beierkuhnlein, C.; Jentsch, A. Burned and devoured—Introduced herbivores, fire and the endemic flora of the high elevation ecosystem on La Palma, Canary Islands. Arct. Antarct. Alp. Res. 2014, 46, 859–869. Available online: http://www.gobiernodecanarias.org/boc/2006/237/011.html (accessed on 28 October 2020). [CrossRef] [Green Version]
- Frankham, R. Do island populations have less genetic variation than mainland populations? Heredity 1997, 78, 311–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Mazuecos, M.; Vargas, P. Genetically depauperate in the continent but rich in oceanic islands: Cistus monspeliensis (Cistaceae) in the Canary Islands. PLoS ONE 2011, 6, e17172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Verdugo, C.; Calleja, J.A.; Vargas, P.; Silva, L.; Moreira, O.; Pulido, F. Polyploidy and microsatellite variation in the relict tree Prunus lusitanica L.: How effective are refugia in preserving genotypic diversity of clonal taxa? Mol. Ecol. 2013, 22, 1546–1557. [Google Scholar] [CrossRef] [PubMed]
- Mort, M.E.; Soltis, D.E.; Soltis, P.S.; Francisco-Ortega, J.; Santos-Guerra, A. Phylogenetics and evolution of the Macaronesian clade of Crassulaceae inferred from nuclear and chloroplast sequence data. Syst. Bot. 2002, 27, 271–288. [Google Scholar]
- Fernández-Palacios, J.M.; de Nascimento, L.; Otto, R.; Delgado, J.D.; García-del-Rey, E.; Arévalo, J.R.; Whittaker, R.J. A reconstruction of Palaeo-Macaronesia, with particular reference to the long-term biogeography of the Atlantic island laurel forests. J. Biogeogr. 2011, 38, 226–246. [Google Scholar] [CrossRef]
- Van den Boogard, P. The origin of the Canary Island Seamount Province—New ages of old seamounts. Sci. Rep. 2017, 3, 2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weigelt, P.; Steinbauer, M.; Cabral, J.S.; Kreft, H. Late Quaternary climate change shapes island biodiversity. Nature 2016, 532, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Aparicio, A.; Martín-Hernanz, S.; Parejo-Farnés, C.; Arroyo, J.; Lavergne, S.; Yeşilyurt, E.B.; Yeşilyurt, M.-L.; Rubio, E.; Albaladejo, R.G. Phylogenetic reconstruction of the genus Helianthemum (Cistaceae) using plastid and nuclear DNA-sequences: Systematic and evolutionary inferences. Taxon 2017, 66, 868–885. [Google Scholar] [CrossRef] [Green Version]
- Sánchez Meseguer, A.; Aldasoro, J.J.; Sanmartín, I. Bayesian inference of phylogeny, morphology and range evolution reveals a complex evolutionary history in St. John’s wort (Hypericum). Mol. Phylogenetics Evol. 2013, 67, 379–403. [Google Scholar] [CrossRef] [PubMed]
- Graham, R.E.; Reyes-Betancourt, J.A.; Chapman, M.A. Inter-island differentiation and contrasting patterns of diversity in the iconic Canary Island sub-alpine endemic Echium wildpretii (Boraginaceae). Syst. Biodivers. 2021, 19, 507–521. [Google Scholar] [CrossRef]
- Schmitt, T. Molekulare Biogeographie; Haupt Verlag: Bern, Germany, 2020; 504p. [Google Scholar]
- Fernández-Palacios, J.M.; Negrín, Z.; Fernández Lugo, S.; Arévalo, J.R.; de Nascimento, L. Terrestrial Biota Checklist of the Chinijo Archipelago and Lobos (Canary Islands). Rev. Sci. Insul. 2018, 1, 51–86. [Google Scholar] [CrossRef]
- Puppo, P.; Curto, M.; Gusmão-Guedes, J.; Cochofel, J.; Pérez de Paz, P.L.; Bräuchler, C.; Meimberg, H. Molecular phylogenetics of Micromeria (Lamiaceae) in the Canary Islands, diversification and inter-island colonization patterns inferred from nuclear genes. Mol. Phylogenetics Evol. 2015, 89, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Plants of the World Online (Kew). Available online: http://www.plantsoftheworldonline.org/ (accessed on 5 June 2021).
- World Flora Online. Available online: http://www.worldfloraonline.org/ (accessed on 5 June 2021).
- The Plant List. Available online: http://www.theplantlist.org/ (accessed on 5 June 2021).
- Roskov, Y.; Abucay, L.; Orrell, T.; Nicolson, D.; Bailly, N.; Kirk, P.M.; Bourgoin, T.; DeWalt, R.E.; Decock, W.; De Wever, A.; et al. (Eds.) Species 2000 & ITIS Catalogue of Life, 2018 Annual Checklist. Species 2000; Naturalis: Leiden, The Netherlands, 2018; Available online: www.catalogueoflife.org/annual-checklist/2018 (accessed on 5 June 2021).
- GBIF. Available online: https://www.gbif.org/ (accessed on 5 June 2021).
- TRY. Available online: https://www.try-db.org/TryWeb/Home.php (accessed on 5 June 2021).
- Kattge, J.; Bönisch, G.; Díaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Tautenhahn, S.; Werner, G.D.A.; Aakala, T.; Abedi, M.; et al. TRY plant trait database—Enhanced coverage and open access. Glob. Chang. Biol. 2020, 26, 119–188. [Google Scholar] [CrossRef] [Green Version]
- Chamberlain, S.; Szocs, E. Taxize—Taxonomic search and retrieval in R. F1000Research 2013, 2, 191. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- International Plant Names Index IPNI. Available online: https://www.ipni.org/ (accessed on 5 June 2021).
- Nezadal, W.; Lindacher, R.; Welss, W. Lokalendemiten und Phytodiversität der westkanadischen Inseln La Palma und La Gomera. Feddes Repert. 1999, 110, 19–30. [Google Scholar] [CrossRef]
- Barone, R. Notas corológicas sobre dos especies de Atriplex, L. (Chenopodiaceae) en Tenerife, Islas Canarias. Botánica Macaronésica 2003, 24, 161–164. [Google Scholar]
- Leibbach, R.; Peter, R. Bemerkungen zu den Orchideen von La Palma (Kanarische Inseln). J. Eur. Orch. 2007, 39, 55–70. [Google Scholar]
- Otto, R.; Scholz, H.; Scholz, S. Supplements to the flora of the Canary Islands, Spain: Poaceae. Willdenowia 2008, 38, 491–496. [Google Scholar] [CrossRef] [Green Version]
- Prina, A.O.; Martinez-Laborde, J.B. A taxonomic revision of Crambe section Dendrocrambe (Brassicaceae). Bot. J. Linn. Soc. 2008, 156, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Verloove, F.; Reyes-Betancort, J. Additions to the flora of Tenerife (Canary Islands, Spain). Collect. Bot. 2011, 30, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Sánchez de Lorenzo Cáceres, J. Sobre la presencia de Balanites aegyptiaca (L.) Del. (Zygophyllaceae) en Tenerife, Islas Canarias. Bouteloua 2012, 10, 82–84. [Google Scholar]
- Reisigl, H. Gefährdete endemische Blütenpflanzen der Trockeninsel Fuerteventura: Herkunft, Ökologie, Gesellschaft. Bauhinia 2013, 24, 39–52. [Google Scholar]
- Verloove, F. New xenophytes from Gran Canaria (Canary Islands, Spain), with emphasis on naturalized and (potentially) invasive species. Collect. Bot. 2013, 32, 59–82. [Google Scholar] [CrossRef] [Green Version]
- Santos Guerra, A.; Reyes Betancort, J.A.; Padrón Mederos, M.A.; Mesa Coello, R. Plantas poco o nada conocidas de la flora vascular silvestre de las Islas Canarias. Bot. Complut. 2013, 37, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Verloove, F.; Guiggi, A. Some new xenophytes from Fuerteventura (Canary Islands, Spain). Bouteloua 2013, 13, 13,–42. [Google Scholar]
- Santos Guerra, A.; Reyes-Betancort, J. Nuevas adiciones y citas de interés para la flora autóctona de las islas Canarias. Vieraea 2014, 42, 249–258. [Google Scholar]
- Otto, R.; Verloove, F. New xenophytes from La Palma (Canary Islands, Spain), with emphasis on naturalized and (potentially) invasive species. Collect. Bot. 2016, 35, e001. [Google Scholar]
- Marrero Rodríguez, Á. Eucaliptos en Gran Canaria, identificación y corología. Hacia una reseña histórica. Botánica Macaronésica 2016, 29, 91–137. [Google Scholar]
- Salas-Pascual, M.; Quintana Vega, G. Salvinia molesta D. S. Mitch. (Salviniaceae), nueva cita para Canarias y España. Botánica Macaronésica 2016, 29, 73–81. [Google Scholar]
- Verloove, F.; Alves, P. New vascular plant records for the western part of the Iberian Peninsula (Portugal and Spain). Folia Bot. Extrem. 2016, 10, 5–23. [Google Scholar]
- Galan de Mera, A.; Linares-Perea, E.; Vicente-Orellana, J.A. Taraxacum (Asteraceae) in the Azores, Madeira and the Canary Island. Ann. Bot. Fenn. 2017, 54, 273–285. [Google Scholar] [CrossRef]
- Verloove, F. New xenophytes from the Canary Islands (Gran Canaria and Tenerife; Spain). Acta Bot. Croat. 2017, 76, 120–131. [Google Scholar] [CrossRef] [Green Version]
- Verloove, F.; Ojeda-Land, E.; Smith, G.F.; Guiggi, A.; Reyes-Betancort, J.A.; Samarín, C.; González Hernández, A.; Barone, R. New records of naturalised and invasive cacti (Cactaceae) from Gran Canaria and Tenerife, Canary Islands, Spain. Bradleya 2017, 35, 58–79. [Google Scholar] [CrossRef]
- Otto, R.; Verloove, F. New xenophytes from La Palma (Canary Islands, Spain), with emphasis on naturalized and (potentially) invasive species—Part 2. Collect. Bot. 2018, 37, e005. [Google Scholar]
- Expósito, A.B.; Siverio, A.; Bermejo, L.A.; Sobrino-Vesperinas, E. Checklist of alien plant species in a natural protected area: Anaga Rural Park (Tenerife, Canary Islands) effect of human infrastructures on their abundance. Plant Ecol. Evol. 2018, 151, 142–152. [Google Scholar] [CrossRef]
- Verloove, F.; Salas-Pascual, M.; Marrero Rodríguez, Á. New records of alien plants for the flora of Gran Canaria (Canary Islands, Spain). Flora Mediterr. 2018, 28, 119–135. [Google Scholar]
- Portero Alvarez, A.M.; Martin-Carbajal Gonzalez, J.; Reyes-Betancort, J.; Mesa Coello, R.A. Lotus gomerythus (Fabaceae-Loteae) Spec. Nova. Botanica Macaronesica 2019, 30, 89–98. [Google Scholar]
- Marrero, A. Vicia tenoi (Fabaceae) una nueca especie del macizo de Teno, Tenerife (Islas Canarias). Bot. Macaronesica 2019, 30, 153–156. [Google Scholar]
- Arango Toro, O. Aeoniun lioi (Crassulaceae): Una nueva especie de Tenerife, Islas Canarias. Bot. Macaronesica 2019, 30, 7–22. [Google Scholar]
- Verloove, F.; Thiede, J.; Marrero Rodríguez, Á.; Salas-Pascual, M.; Reyes-Betancort, J.; Ojeda-Land, E.; Smith, G. A synopsis of feral Agave and Furcraea (Agavaceae, Asparagaceae s. lat.) in the Canary Islands (Spain). Plant Ecol. Evol. 2019, 152, 470–498. [Google Scholar] [CrossRef]
- Marrero Rodríguez, Á. Adiciones corológicas a la flora vascular de Gran Canaria, especies xenófitas, ocasionales o potenciales invasoras. Botánica Macaronésica 2019, 30, 121–142. [Google Scholar]
- Otto, R.; Verloove, F. New xenophytes from La Palma (Canary Islands, Spain), with emphasis on naturalized and (potentially) invasive species—Part 3. Collect. Bot. 2020, 39, e001. [Google Scholar]
- Verloove, F.; Déniz Suárez, E.; Salas Pascual, M. New records of non-native vascular plants in Gran Canaria (Spain, Canary Islands). Flora Mediterr. 2020, 30, 121–136. [Google Scholar]
- White, O.W.; Reyes-Betancort, J.A.; Chapman, M.A.; Carine, M.A. Geographical isolation, habitat shifts and hybridisation in the diversification of the Macaronesian endemic genus Argyranthemum (Asteraceae). New Phytol. 2020, 228, 1953–1971. [Google Scholar] [CrossRef] [PubMed]
- Verloove, F. New records in vascular plants alien to Tenerife (Spain, Canary Islands). Biodivers. Data J. 2021, 9, e62878. [Google Scholar] [CrossRef] [PubMed]
- White, O.W.; Reyes-Betancort, J.A.; Chapman, M.A.; Carine, M.A. Recircumscription of the Canary Island endemics Argyranthemum broussonetii and A. callichrysum (Asteraceae: Anthemideae) based on evolutionary relationships and morphology. Willdenowia 2021, 51, 129–139. [Google Scholar] [CrossRef]
- Nürk, N.M.; Linder, H.P.; Onstein, R.E.; Larcombe, M.J.; Hughes, C.E.; Fernández, L.P.; Schlüter, P.M.; Valente, L.; Beierkuhnlein, C.; Cutts, V.; et al. Diversification in evolutionary arenas—Assessment and synthesis. Ecol. Evol. 2020, 10, 6163–6182. [Google Scholar] [CrossRef] [PubMed]
- Darwin, C. Letter no. 2130. To, J.D. Hooker, 1 August 1857. Darwin Correspondence Project. Available online: https://www.darwinproject.ac.uk/ (accessed on 5 July 2021).
- Irl, S.; Schweiger, A.; Medina, F.M.; Fernández-Palacios, J.M.; Harter, D.; Jentsch, A.; Provenzale, A.; Steinbauer, M.J.; Beierkuhnlein, C. An island view of endemic rarity—Environmental drivers and consequences for nature conservation. Divers. Distrib. 2017, 23, 1132–1142. [Google Scholar] [CrossRef] [Green Version]
- García-Verdugo, C.; Baldwin, B.G.; Fay, M.F.; Caujapé-Castells, J. Life history traits and patterns of diversification in oceanic archipelagos: A meta-analysis. Bot. J. Linn. Soc. 2014, 174, 334–348. [Google Scholar] [CrossRef] [Green Version]
- EU Biodiversity Strategy for 2030. Available online: https://www.eea.europa.eu/policy-documents/eu-biodiversity-strategy-for-2030-1 (accessed on 17 July 2021).
- Gangoso, L.; Donázar, J.A.; Scholz, S.; Palacios, C.J.; Hiraldo, F. Contradiction in conservation of island ecosystems: Plants, introduced herbivores and avian scavengers in the Canary Islands. Biodivers. Conserv. 2006, 15, 2231–2248. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Palacios, J.M.; Martin Esquivel, J.L. Naturaleza de las Islas Canarias—Ecologia y Conservation; Publicaciones Turquesa: Santa Cruz de Tenerife, Spain, 2001; 474p. [Google Scholar]
- Reyes-Betancort, J.A.; Santos-Guerra, A.; Guma, I.R.; Humphries, C.J.; Carine, M.A. Diversity, rarity and the evolution and conservation of the Canary Islands endemic flora. An. Jardín Botánico Madr. 2008, 65, 25–45. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxonomic Level | FloCan | Atlantis Biota | Muer et al. | POWO | COL | WFO | GBIF | TRY |
|---|---|---|---|---|---|---|---|---|
| Year | 2021 | 2021 | 2016 | 2021 | 2021 | 2021 | 2021 | 2021 |
| Families | 171 | 162 | 155 | 171 | 166 | 164 | 170 | 166 |
| Genera | 863 | 747 | 699 | 854 | 815 | 775 | 832 | 788 |
| Species | 2416 | 2050 | 1834 | 2370 | 2207 | 2022 | 2284 | 1965 |
| Infraspecifics | 396 | 295 | 222 | 247 | 264 | 125 | 329 | 74 |
| Taxonomic Level | FloCan | Atlantis Biota | Muer et al. |
|---|---|---|---|
| Year | 2021 | 2021 | 2016 |
| Species | 608 | 541 | 499 |
| Infraspecifics | 197 | 147 | 152 |
| Taxonomic Level | FloCan | Atlantis Biota | Muer et al. |
|---|---|---|---|
| Year | 2021 | 2021 | 2016 |
| Species | 964 | 781 | 662 |
| Infraspecifics | 67 | 44 | 16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beierkuhnlein, C.; Walentowitz, A.; Welss, W. FloCan—A Revised Checklist for the Flora of the Canary Islands. Diversity 2021, 13, 480. https://doi.org/10.3390/d13100480
Beierkuhnlein C, Walentowitz A, Welss W. FloCan—A Revised Checklist for the Flora of the Canary Islands. Diversity. 2021; 13(10):480. https://doi.org/10.3390/d13100480
Chicago/Turabian StyleBeierkuhnlein, Carl, Anna Walentowitz, and Walter Welss. 2021. "FloCan—A Revised Checklist for the Flora of the Canary Islands" Diversity 13, no. 10: 480. https://doi.org/10.3390/d13100480
APA StyleBeierkuhnlein, C., Walentowitz, A., & Welss, W. (2021). FloCan—A Revised Checklist for the Flora of the Canary Islands. Diversity, 13(10), 480. https://doi.org/10.3390/d13100480