
Spatial Patterns of Coral Community Structure in the Toliara Region of Southwest Madagascar and Implications for Conservation and Management

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
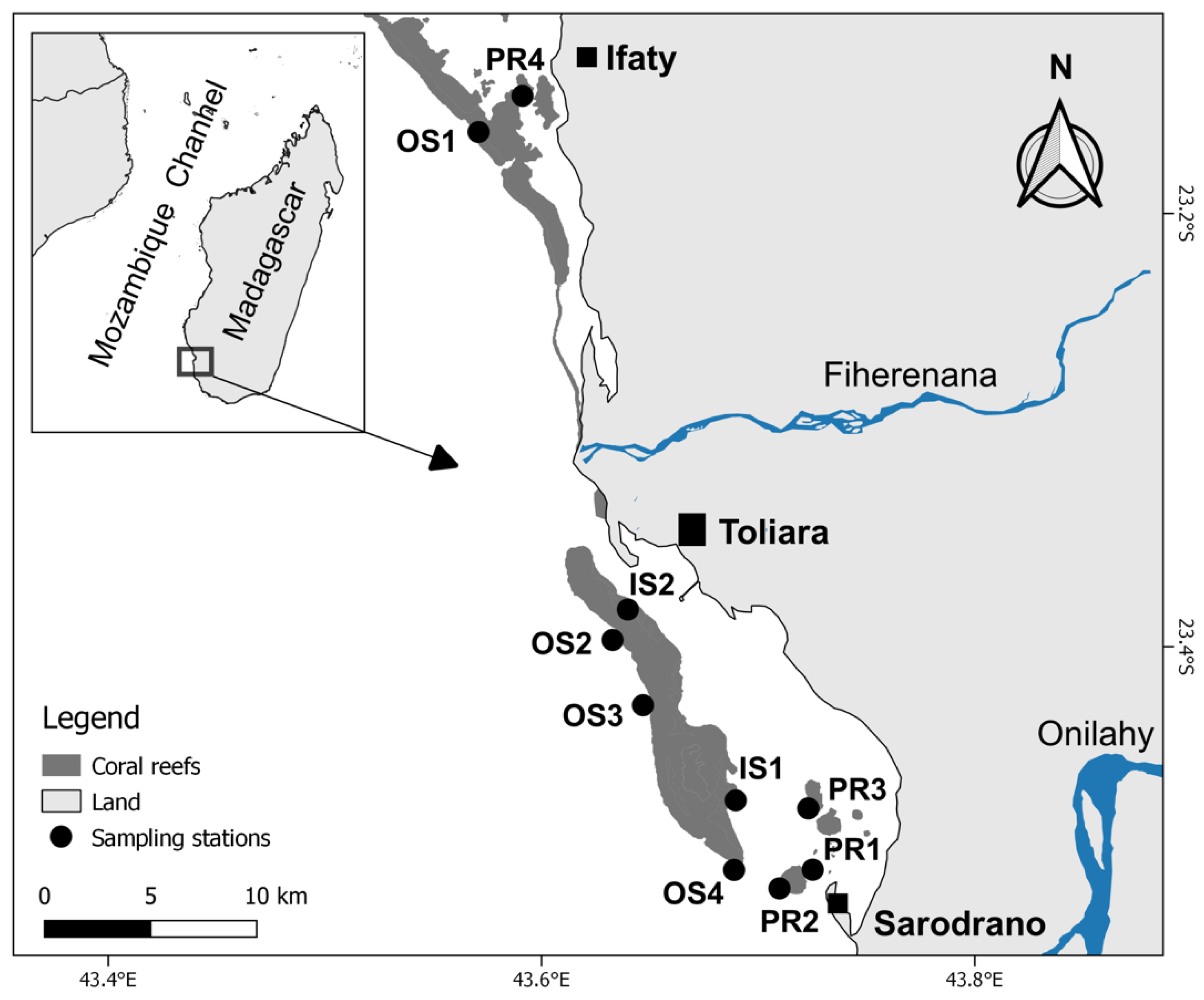
2.1. Study Area
2.2. Sampling Strategy
2.3. Data Analysis
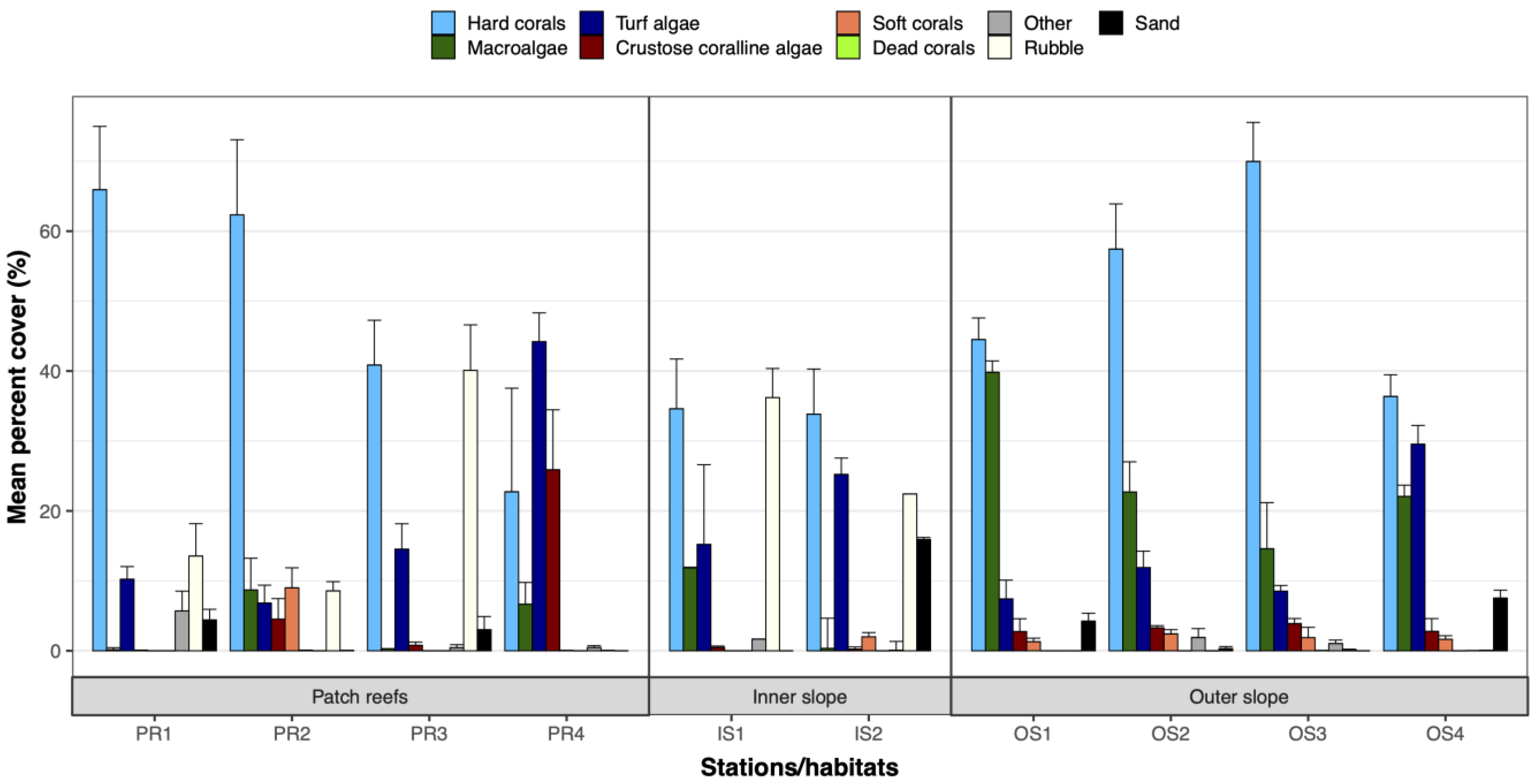
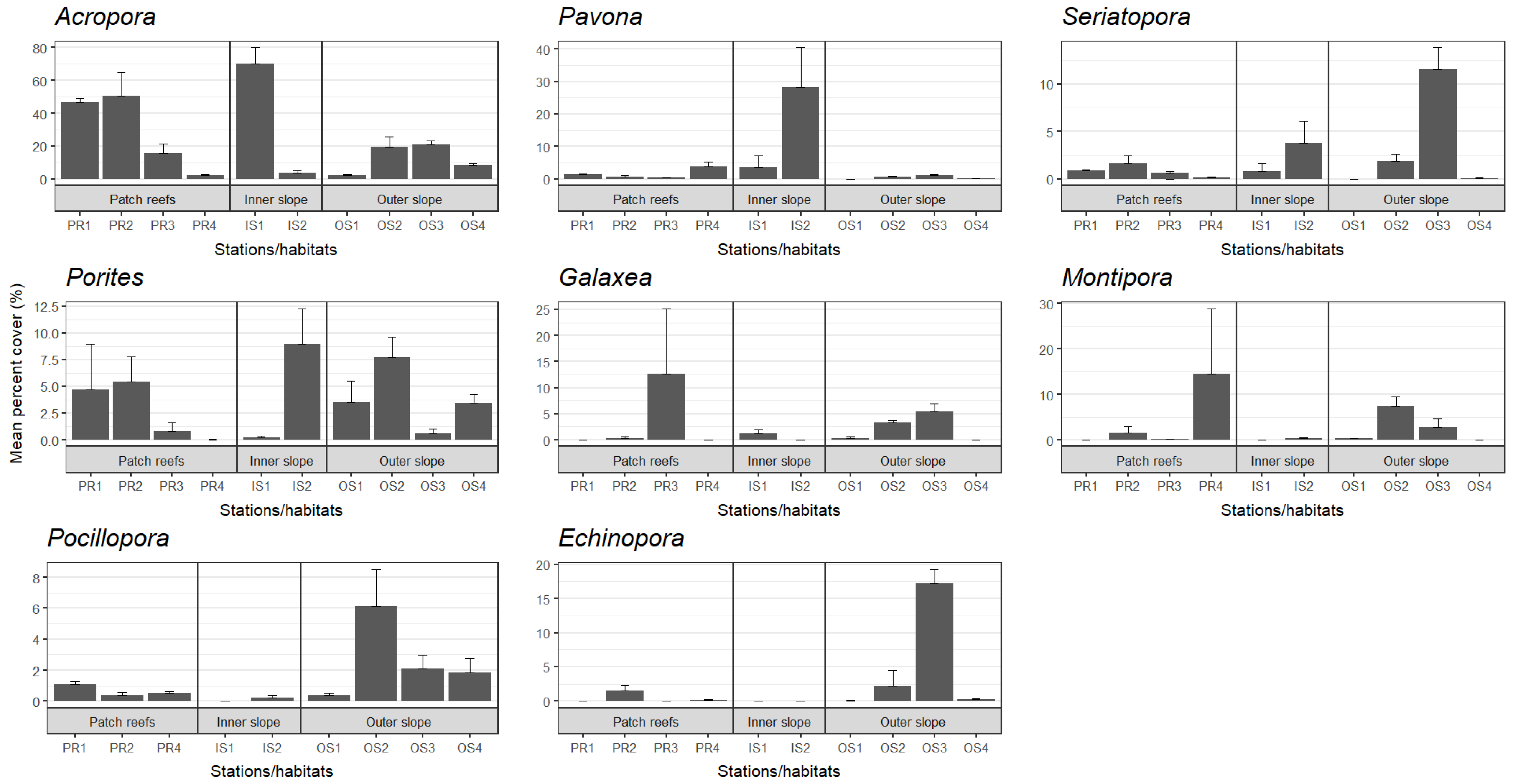
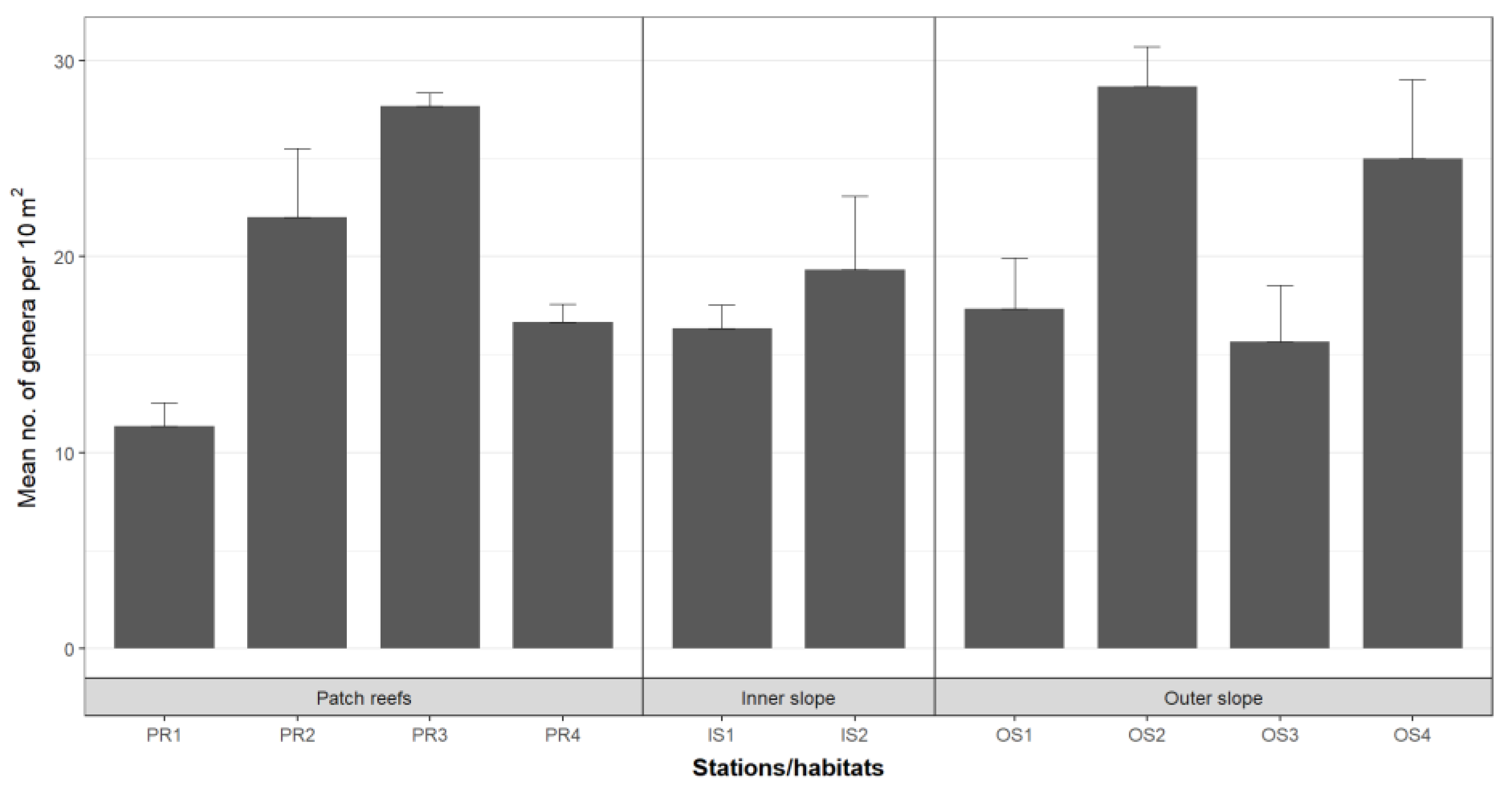
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. Available online: http://www.ncbi.nlm.nih.gov/pubmed/10706275 (accessed on 18 August 2021). [CrossRef] [PubMed]
- Brooks, T.M.; Mittermeier, R.A.; Da Fonseca, G.A.B.; Gerlach, J.; Hoffmann, M.; Lamoreux, J.F.; Mittermeier, C.G.; Pilgrim, J.D.; Rodrigues, A.S.L. Global biodiversity conservation priorities. Science 2006, 313, 58–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooke, A. Madagascar: A guide to marine biodiversity; Wildlife Conservation Society: Antananarivo, Madagascar, 2012; p. 172. [Google Scholar]
- Veron, J.E.N.; Turak, E. Zooxanthellate scleractinia of Madagascar. Reef corals of Northwest Madagascar. In A Rapid Marine Biodiversity Assessment of Northwest Madagascar; McKenna, S.A., Allen, G.R., Eds.; Bulletin of the Rapid Assessment Program 31, Conservation International: Washington, DC, USA, 2003; pp. 23–25. [Google Scholar]
- Obura, D.; Di Carlo, G.; Rabearisoa, A.; Oliver, T. A Rapid Marine Biodiversity Assessment of the Coral Reefs of Northeast Madagascar; RAP Bulletin of Biological Assessment 61; Conservation International: Arlington, VA, USA, 2011; p. 100. [Google Scholar]
- Fricke, R.; Mahafina, J.; Behivoke, F.; Jaonalison, H.; Léopold, M.; Ponton, D. Annotated checklist of the fishes of Madagascar, southwestern Indian Ocean, with 158 new records. FishTaxa 2018, 3, 1–432. [Google Scholar]
- Pichon, M. The coral reefs of Madagascar. In Biogeography and Ecology in Madagascar; Battistini, R., Richard-Vindard, G., Eds.; The Hague: Dordrecht, The Netherlands, 1972; pp. 367–410. [Google Scholar]
- Ahamada, S.; Bijoux, J.; Cauvin, B.; Hagan, A.; Harris, A.; Koonjul, M.; Meunier, S.; Quod, J.P. Status of coral reefs of the South-West Indian Ocean islands states: Comoros, Madagascar, Mauritius, Reunion, Seychelles. In Status of Coral Reefs of the World: 2008; Wilkinson, C.R., Ed.; Australian Institute of Marine Science: Townsville, Australia, 2008; pp. 105–118. [Google Scholar]
- Obura, D. Resilience and climate change: Lessons from coral reefs and bleaching in the Western Indian Ocean. Estuar. Coast. Shelf Sci. 2005, 63, 353–372. [Google Scholar] [CrossRef]
- Harmelin-Vivien, M. Trophic relationships of reef fishes in Tuléar (Madagascar). Oceanol. Acta 1981, 4, 365–374. [Google Scholar]
- Rasoarimalala, F. Variation de la Biodiversité Spécifique des Peuplements de Poissons sur le Grand Récif de Toliara (Sud-Ouest de Madagascar), entre 1970 et 1997. Master’s Thesis, University of Toliara, Toliara, Madagascar, 2001. [Google Scholar]
- Pichon, M. Recherches sur les peuplements à dominance d’anthozoaires dans les récifs coralliens de Tuléar (Madagascar). Atoll Res. Bull. 1978, 222, 1–447. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, C.R.C. Biodiversity patterns in Indian Ocean corals, and effects of taxonomic error in data. Biodivers. Conserv. 1998, 7, 847–868. [Google Scholar] [CrossRef]
- Laroche, J.; Ramananarivo. A preliminary survey of the artisanal fishery on coral reefs of the Tulear Region (Southwest Madagascar). Coral Reefs 1995, 14, 193–200. [Google Scholar] [CrossRef]
- Humber, F.; Andriamahaino, E.T.; Beriziny, T.; Botosoamananto, R.; Godley, B.; Gough, C.; Pedron, S.; Ramahery, V.; Broderick, A. Assessing the small-scale shark fishery of Madagascar through community-based monitoring and knowledge. Fish. Res. 2017, 186, 131–143. [Google Scholar] [CrossRef]
- Lemahieu, A.; Scott, L.; Malherbe, W.S.; Mahatante, P.T.; Randrianarimanana, J.V.; Aswani, S. Local perceptions of environmental changes in fishing communities of southwest Madagascar. Ocean. Coast. Manag. 2018, 163, 209–221. [Google Scholar] [CrossRef]
- Ahamada, S.; Bigot, L.; Bijoux, J.; Maharavo, J.; Meunier, S.; Moyne-Picard, M.; Paupiah, N. Status of coral reefs in the South West Indian Ocean island node: Comoros, Madagascar, Mauritius, Reunion and Seychelles. In Status of CoralRreefs of the World: 2002; Wilkinson, C.R., Ed.; Australian Institute of Marine Science: Townsville, Australia, 2002; pp. 79–100. [Google Scholar]
- Cochrane, K.L.; Rakotondrazafy, H.; Aswani, S.; Chaigneau, T.; Downey-Breedt, N.; Lemahieu, A.; Paytan, A.; Pecl, G.; Plag-anyi, E.; Popova, E.; et al. Tools to enrich vul-nerability assessment and adaptation planning for coastal communities in data-poor regions: Application to a case study in Madagascar. Front. Mar. Sci. 2019, 5, 505. [Google Scholar] [CrossRef] [Green Version]
- Harding, S.; Randriamanantsoa, B.; Hardy, T.; Curd, A. Coral Reef Monitoring and Biodiversity Assessment to Support the Planning of a Proposed MPA at Andavadoaka; Wildlife Conservation Societ: London, UK, 2006; p. 100. [Google Scholar]
- Harding, S.; Randriamanantsoa, B. Coral reef monitoring in marine reserves of northern Madagascar. In Ten years after Bleaching—Facing the Consequences of Climate Change in the Indian Ocean; Coral Degradation in the Indian Ocean: Status Report; Obura, D., Tamelander, J., Linden, O., Eds.; CORDIO, Coastal Oceans Research and Development in the Indian Ocean/Sida-SAREC: Mombasa, Kenya, 2008; pp. 93–106. [Google Scholar]
- Obura, D.; Gudka, M.; Rabi, F.A.; Gian, S.B.; Bijoux, J.; Freed, S.; Maharavo, J.; Mwaura, J.; Porter, S.; Sola, E.; et al. Coral Reef Status Report for the Western Indian Ocean (2017); Global Coral Reef Monitoring Network (GCRMN)/International Coral Reef Initiative (ICRI): Nairobi, Kenya, 2017; p. 144. [Google Scholar]
- Gough, C.L.A.; Dewar, K.M.; Godley, B.J.; Zafindranosy, E.; Broderick, A.C. Evidence of overfishing in small-scale fisheries in Madagascar. Front. Mar. Sci. 2020, 7, 317. [Google Scholar] [CrossRef]
- Payet, E.; Dumas, P.; Pennober, G. Modélisation de l’érosion hydrique des sols sur un bassin versant du sud-ouest de Madagascar, le Fiherenana. VertigO-La Rev. Électronique Sci. Environ. 2012, 11, 1–25. [Google Scholar] [CrossRef]
- Sheridan, C.; Baele, J.; Kushmaro, A.; Fréjaville, Y.; Eeckhaut, I. Terrestrial runoff influences white syndrome prevalence in SW Madagascar. Mar. Environ. Res. 2014, 101, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.; Manahira, G.; Sheppard, A.; Gouch, C.; Sheppard, C. Demise of Madagascar’s once great barrier reef: Changes in coral reef conditions over 40 years. Atoll Res. Bull. 2010, 574, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Bruggemann, J.H.; Rodier, M.; Guillaume, M.M.; Andréfouët, S.; Arfi, R.; Cinner, J.E.; Pichon, M.; Ramahatratra, F.; Rasoamanendrika, F.; Zinke, J.; et al. Wicked social-ecological problems forcing unprecedented change on the latitudinal margins of coral reefs: The case of Southwest Madagascar. Ecol. Soc. 2012, 17, 47. [Google Scholar] [CrossRef]
- Andréfouët, S.; Guillaume, M.M.M.; Delval, A.; Rasoamanendrika, F.M.A.; Blanchot, J.; Bruggemann, J.H. Fifty years of changes in reef flat habitats of the Grand Récif of Toliara (SW Madagascar) and the impact of gleaning. Coral Reefs 2013, 32, 757–768. [Google Scholar] [CrossRef]
- Quod, J.P.; Bigot, L. Coral bleaching in the Indian Ocean islands: Ecological conse-quences and recovery in Madagascar, Comoros, Mayotte and Reunion. In Coral Reef Degradation in the Indian Ocean (CORDIO); Sout-er, D., Obura, D., Linden, O., Eds.; CORDIO SAREC Marine Science Program: Stockholm, Sweden, 2000; pp. 108–113. [Google Scholar]
- Todinanahary, G.G.; Refoty, M.E.; Terrana, L.; Lavitra, T.; Eeckhaut, I. Previously unlisted scleractinian species recorded from the Great Reef of Toliara, southwest Madagascar. West. Indian Ocean J. Mar. Sci. 2018, 17, 67. [Google Scholar] [CrossRef] [Green Version]
- Nadon, M.; Griffiths, D.; Doherty, E.; Harris, A. The status of coral reefs in the remote region of Andavadoaka, Southwest Madagascar. West. Indian Ocean J. Mar. Sci. 2009, 6, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Vermeij, M.J.A.; Frade, P.R.; Jacinto, R.I.R.; Debrot, A.O.; Bak, R.P.M. Effects of reproductive mode on habitat-related differences in the population structure of eight Caribbean coral species. Mar. Ecol. Prog. Ser. 2007, 351, 91–102. [Google Scholar] [CrossRef]
- Williams, G.J.; Smith, J.E.; Conklin, E.J.; Gove, J.M.; Sala, E.; Sandin, S.A. Benthic communities at two remote Pacific coral reefs: Effects of reef habitat, depth, and wave energy gradients on spatial patterns. PeerJ 2013, 1, e81. [Google Scholar] [CrossRef] [Green Version]
- Doropoulos, C.; Ward, S.; Roff, G.; Gonzalez-Rivero, M.; Mumby, P. Linking demographic processes of juvenile corals to benthic recovery trajectories in two common reef habitats. PLoS ONE 2015, 10, e0128535. [Google Scholar] [CrossRef] [Green Version]
- Mahafina, J. Perception et Comportements des Pêcheurs pour une Gestion Durable de la Biodiversité et de la Pêcherie Récifale: Application au Niveau des Réserves Marines Temporaires du Sud-Ouest de Madagascar. Ph.D. Thesis, University of Toliara, Toliara, Madagascar, 2011. [Google Scholar]
- Ratsimbazafy, H.; Lavitra, T.; Kochzius, M.; Hugé, J. Emergence and diversity of marine protected areas in Madagascar. Mar. Policy 2019, 105, 91–108. [Google Scholar] [CrossRef]
- Maina, J.; de Moel, H.; Vermaat, J.; Bruggemann, J.H.; Guillaume, M.; Grove, C.A.; Madin, J.; Mertz-Kraus, R.; Zinke, J. Linking coral river runoff proxies with climate variability, hydrology and land-use in Madagascar catchments. Mar. Pollut. Bull. 2012, 64, 2047–2059. [Google Scholar] [CrossRef]
- Matyas, C.J. Tropical cyclone formation and motion in the Mozambique Channel: Cyclones in the Mozambique Channel. Int. J. Climatol. 2015, 35, 375–390. [Google Scholar] [CrossRef]
- Côté-Laurin, M.-C.; Benbow, S.; Erzini, K. The short-term impacts of a cyclone on seagrass communities in Southwest Madagascar. Cont. Shelf Res. 2017, 138, 132–141. [Google Scholar] [CrossRef]
- Gérard, K.; Roby, C.; Chevalier, N.; Thomassin, B.; Chenuil, A.; Féral, J.-P. Assessment of three mitochondrial loci variability for the crown-of-thorns starfish: A first insight into Acanthaster phylogeography. Comptes Rendus Biol. 2008, 331, 137–143. [Google Scholar] [CrossRef] [PubMed]
- McClanahan, T.R.; Ateweberhan, M.; Graham, N.A.J.; Wilson, S.K.; Sebastián, C.R.; Guillaume, M.M.; Bruggemann, J.H. Western Indian Ocean coral communities: Bleaching responses and susceptibility to extinction. Mar. Ecol. Progr. Ser. 2007, 337, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kohler, K.E.; Gill, S.M. Coral point count with excel extensions (CPCe): A visual basic program for the determination of coral and substrate coverage using random point count methodology. Comput. Geosci. 2006, 32, 1259–1269. [Google Scholar] [CrossRef]
- Adjeroud, M.; Poisson, E.; Peignon, C.; Penin, L.; Kayal, M. Spatial Patterns and short-term changes of coral assemblages along a cross-shelf gradient in the Southwestern Lagoon of New Caledonia. Diversity 2019, 11, 21. [Google Scholar] [CrossRef] [Green Version]
- Webster, F.J.; McMahon, K. An ASSESSMENT OF CORAL REEFs in Northwest Madagascar. In Coral Degradation in the Indian Ocean: Status Report; Linden, O., Souter, D., Wilhelmsson, D., Obura, D., Eds.; CORDIO, Department of Biology and Environmental Science, University of Kalmar: Kalmar, Sweden, 2002; pp. 190–201. [Google Scholar]
- Veron, J.E.N.; Turak, E. Reef corals of Northwest Madagascar. In A Rapid Marine Biodiversity Assessment of Northwest Madagascar; McKenna, S.A., Allen, G.R., Eds.; Bulletin of the Rapid Assessment Program 31, Conservation International: Washington, DC, USA, 2003; pp. 26–31. [Google Scholar]
- Ramanantsoa, J.D.; Penven, P.; Krug, M.; Gula, J.; Rouault, M. Uncovering a new current: The Southwest MAdagascar coastal current. Geophys. Res. Lett. 2018, 45, 1930–1938. [Google Scholar] [CrossRef] [Green Version]
- Bauman, A.G.; Pratchett, M.S.; Baird, A.H.; Riegl, B.; Heron, S.; Feary, D.A. Variation in the size structure of corals is related to environmental extremes in the Persian Gulf. Mar. Environ. Res. 2013, 84, 43–50. [Google Scholar] [CrossRef]
- Adjeroud, M.; Mauguit, Q.; Penin, L. The size-structure of corals with contrasting life-histories: A multi-scale analysis across environmental conditions. Mar. Environ. Res. 2015, 112, 131–139. [Google Scholar] [CrossRef]
- Grimsditch, G.; Pisapia, C.; Huck, M.; Karisa, J.; Obura, D.; Sweet, M. Variation in size frequency distribution of coral populations under different fishing pressures in two contrasting locations in the Indian Ocean. Mar. Environ. Res. 2017, 131, 146–155. [Google Scholar] [CrossRef] [Green Version]
- Hughes, T.P.; Jackson, J.B.C. Population dynamics and life histories of foliaceous corals. Ecol. Monogr. 1985, 55, 141–166. [Google Scholar] [CrossRef]
- Babcock, R.; Davies, P. Effects of sedimentation on settlement of Acropora millepora. Coral Reefs 1991, 9, 205–208. [Google Scholar] [CrossRef]
- Harrison, P.L. Sexual reproduction of scleractinian corals. In Coral Reefs: An Ecosystem in Transition, 1st ed.; Dubinsky, Z., Stambler, N., Eds.; Springer Science + Business Media, B.V.: Dordrecht, The Nehterlands, 2011; pp. 59–85. [Google Scholar]
- Connell, J.H.; Hughes, T.P.; Wallace, C.C. A 30-year study of coral abundance, recruitment, and disturbance at several scales in space and time. Ecol. Monogr. 1997, 67, 461–488. [Google Scholar] [CrossRef]
- Ninio, R.; Meekan, M. Spatial patterns in benthic communities and the dynamics of a mosaic ecosystem on the Great Barrier Reef, Australia. Coral Reefs 2002, 21, 95–104. [Google Scholar] [CrossRef]
- Cleary, D.F.R.; Polónia, A.R.M.; Renema, W.; Hoeksema, B.W.; Rachello-Dolmen, P.G.; Moolenbeek, R.G.; Budiyanto, A.; Yahmantoro; Tuti, Y.; Giyanto; et al. Variation in the composition of corals, fishes, sponges, echinoderms, ascidians, molluscs, foraminifera and macroalgae across a pronounced in-to-offshore environmental gradient in the Jakarta Bay–Thousand Islands coral reef complex. Mar. Pollut. Bull. 2016, 110, 701–717. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.M.; Scheiblin, R.E. Turf wars: Competition between foundation and turf-forming species on temperate and tropical reefs and its role in regime shifts. Mar. Ecol. Prog. Ser. 2018, 590, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Todinanahary, G.G.B.; Hasintantely, N.; Eeckhaut, I.; Lavitra, T. First evaluation of coral recruitment in Madagascar. WIO J. Mar. Sci. 2021, 20, 47–62. [Google Scholar]
- Loya, Y.; Sakai, K.; Yamazato, K.; Nakano, Y.; Sambali, H.; van Woesik, R. Coral bleaching: The winners and the losers. Ecol. Lett. 2001, 4, 122–131. [Google Scholar] [CrossRef]
- Guest, J.R.; Baird, A.H.; Maynard, J.A.; Muttaqin, E.; Edwards, A.J.; Campbell, S.; Yewdall, K.; Affendi, Y.A.; Chou, L.M. Contrasting patterns of coral bleaching susceptibility in 2010 suggest an adaptive response to thermal stress. PLoS ONE 2012, 7, e33353. [Google Scholar] [CrossRef]
- Adjeroud, M.; Kayal, M.; Peignon, C.; Juncker, M.; Mills, S.C.; Beldade, R.; Dumas, P. Ephemeral and localized outbreaks of the coral predator acanthaster cf. solaris in the Southwestern Lagoon of New Caledonia. Zool. Stud. 2018, 57, e4. [Google Scholar]
- Hirose, Y.; Aini, S.N.; Yamashiro, H. Contact reactions between individuals of the coral-killing sponge Terpios hoshinota. Zool. Stud. 2021, 60, 41. [Google Scholar]
- Hu, J.; Zhang, Y.; Xie, J.Y.; Qiu, J.W. A new species of predatory nudibranch (Gastropoda: Trinchesiidae) of the scleractinian coral Pavona decussata. Zool. Stud. 2020, 59, 30. [Google Scholar]
- Camp, E.; Edmondson, J.; Doheny, A.; Rumney, J.; Grima, A.; Huete, A.; Suggett, D. Mangrove lagoons of the Great Barrier Reef support coral populations persisting under extreme environmental conditions. Mar. Ecol. Prog. Ser. 2019, 625, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Lord, K.S.; Lesneski, K.; Bengtsson, Z.A.; Kuhn, K.M.; Madin, J.; Cheung, B.; Ewa, R.; Taylor, J.F.; Burmester, E.M.; Morey, J.; et al. Multi-year viability of a reef coral population living on mangrove roots suggests an important role for mangroves in the broader habitat mosaic of corals. Front. Mar. Sci. 2020, 7, 377. [Google Scholar] [CrossRef]
- Dumas, P.; Jimenez, H.; Peignon, C.; Wantiez, L.; Adjeroud, M. Small-scale habitat structure modulates the effects of no-take marine reserves for coral reef macroinvertebrates. PLoS ONE 2013, 8, e58998. [Google Scholar] [CrossRef] [PubMed]
- Todinanahary, G.G.B.; Behivoke, F.; Nomenisoa, A.L.; Ravelojaona, D.K.; Rakotoson, A.T.; Tatangirafeno, S.; Rakotonjanahary, F.; Tsiresy, G.; Mara, E.R.; Eeckhaut, I.; et al. Inventaire et Étude de Faisabilité de Sites Propices à l’algoculture, L’holothuriculture, la Gestion de L’exploitation de Poulpes et de Crabes Dans La Région Atsimo Andrefana, Rapport d’étude; MHSA—PRU (Contrat n° 166/C/PIC2/2016): Toliara, Madagascar, 2016; p. 357. [Google Scholar]
- Belle, E.M.S.; Stewart, G.W.; De Ridder, B.; Komeno, R.J.L.; Ramahatratra, F.; Remy-Zephir, B.; Stein-Rostaing, R.D. Es-tablishment of a community managed marine reserve in the Bay of Ranobe, southwest Madagascar. Madag. Conser. Dev. 2009, 4, 31–37. [Google Scholar]
- Okubo, N.; Taniguchi, H.; Motokawa, T. Successful methods for transplanting fragments of Acropora formosa and Acropora hyacinthus. Coral Reefs 2005, 24, 333–342. [Google Scholar] [CrossRef]
- Ferse, S.C.A.; Hein, M.Y.; Rölfer, L. A survey of current trends and suggested future directions in coral transplantation for reef restoration. PLoS ONE 2021, 16, e0249966. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Botosoamananto, R.L.; Todinanahary, G.; Razakandrainy, A.; Randrianarivo, M.; Penin, L.; Adjeroud, M. Spatial Patterns of Coral Community Structure in the Toliara Region of Southwest Madagascar and Implications for Conservation and Management. Diversity 2021, 13, 486. https://doi.org/10.3390/d13100486
Botosoamananto RL, Todinanahary G, Razakandrainy A, Randrianarivo M, Penin L, Adjeroud M. Spatial Patterns of Coral Community Structure in the Toliara Region of Southwest Madagascar and Implications for Conservation and Management. Diversity. 2021; 13(10):486. https://doi.org/10.3390/d13100486
Chicago/Turabian StyleBotosoamananto, Radonirina Lebely, Gildas Todinanahary, Andriamanjato Razakandrainy, Mahery Randrianarivo, Lucie Penin, and Mehdi Adjeroud. 2021. "Spatial Patterns of Coral Community Structure in the Toliara Region of Southwest Madagascar and Implications for Conservation and Management" Diversity 13, no. 10: 486. https://doi.org/10.3390/d13100486
APA StyleBotosoamananto, R. L., Todinanahary, G., Razakandrainy, A., Randrianarivo, M., Penin, L., & Adjeroud, M. (2021). Spatial Patterns of Coral Community Structure in the Toliara Region of Southwest Madagascar and Implications for Conservation and Management. Diversity, 13(10), 486. https://doi.org/10.3390/d13100486