
Evaluation of the Use of Autonomous Reef Monitoring Structures (ARMS) for Describing the Species Diversity of Two Coral Reefs in the Yucatan Peninsula, Mexico

 , , , , , ,
, , , , , ,  , , , , , , and add
Show full author list
, , , , , , and add
Show full author list
Abstract
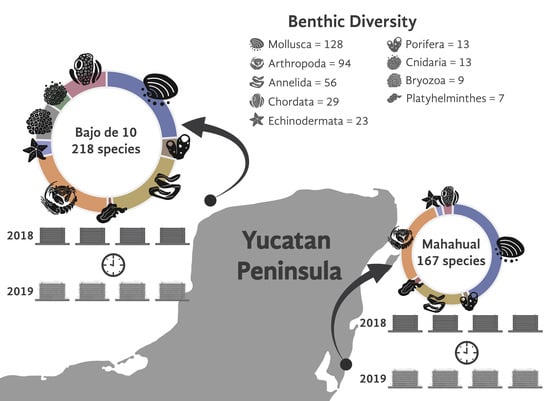
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling
2.3. Statistical Analyses
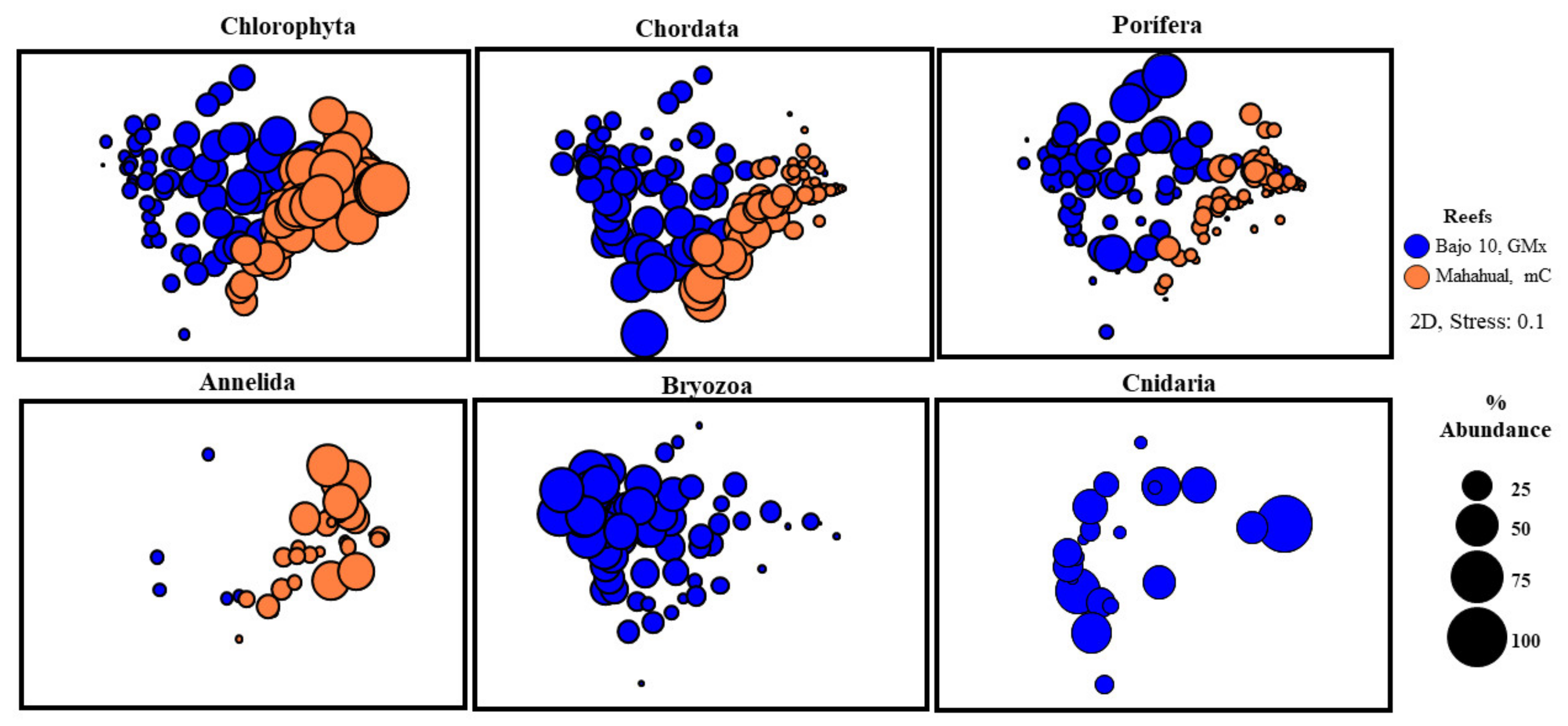
3. Results
3.1. Diversity of the Cryptofauna Assemblage
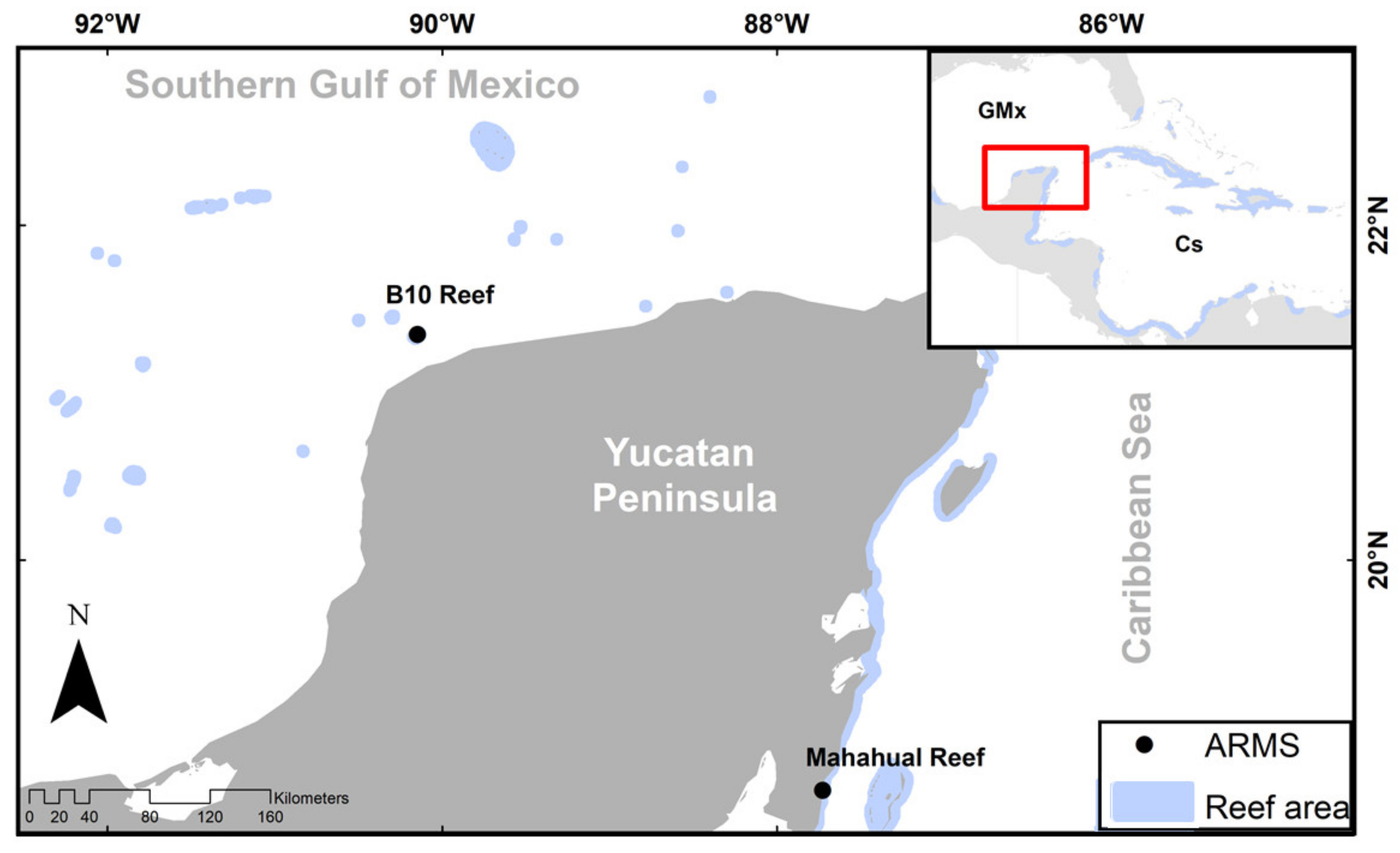
3.2. Taxonomic Distinctness
3.3. Abundance of Sessile Biota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ginsburg, R.N. Geological and biological roles of cavities in coral reefs. In Perspectives on Coral Reefs; Barnes, J.D., Ed.; Australian Institute of Marine Science: Townsville, Australia, 1983; pp. 148–153. [Google Scholar]
- Pearman, J.K.; Leray, M.; Villalobos, R.; Machida, R.J.; Berumen, M.L.; Knowlton, N.; Carvalho, S. Cross-shelf investigation of coral reef cryptic benthic organisms reveals diversity patterns of the hidden majority. Sci. Rep. 2018, 8, 1–17. [Google Scholar] [CrossRef]
- Jackson, J.B.C.; Winston, J.E. Ecology of cryptic coral reef communities. I. Distribution and abundance of major groups of encrusting organisms. J. Exp. Mar. Biol. Ecol. 1982, 57, 135–147. [Google Scholar] [CrossRef]
- Reaka, M.L. Adult-juvenile interactions in benthic reef crustaceans. Bull. Mar. Sci. 1987, 41, 108–134. [Google Scholar]
- Coen, L.D. Herbivory by Caribbean majid crabs: Feeding ecology and plant susceptibility. J. Exp. Mar. Biol. Ecol. 1988, 122, 257–276. [Google Scholar] [CrossRef]
- Rothans, T.C.; Miller, A.C. A link between biologically imported particulate organic nutrients and the detritus food web in reef communities. Mar. Biol. 1991, 110, 145–150. [Google Scholar] [CrossRef]
- Fleur van Duyl, C.; Moodley, L.; Nieuwland, G.; van Ijzerloo, L.; van Soest, R.W.M.; Houtekamer, M.; Meesters, E.H.; Middelburg, J.J. Coral cavity sponges depend on reef-derived food resources: Stable isotope and fatty acid constraints. Mar. Biol. 2011, 158, 1653–1666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plaisance, L.; Brainard, R.; Julian Caley, M.; Knowlton, N. Using DNA barcoding and standardized sampling to compare geographic and habitat differentiation of crustaceans: A Hawaiian Islands example. Diversity 2011, 3, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Costello, M.J.; Basher, Z.; McLeod, L.; Asaad, I.; Claus, S.; Vandepitte, L.; Yasuhara, M.; Gislason, H.; Edwards, M.; Appeltans, W.; et al. Methods for the study of marine biodiversity. In The GEO Handbook on Biodiversity Observation Networks; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Monroy-Velázquez, L.; Rodríguez-Martínez, R.; Blanchon, P.; Alvarez, F. The use of artificial substrate units to improve inventories of cryptic crustacean species on Caribbean coral reefs. PeerJ 2020, 8, e10389. [Google Scholar] [CrossRef]
- Obst, M.; Exter, K.; Allcock, A.L.; Arvanitidis, C.; Axberg, A.; Bustamante, M.; Cancio, I.; Carreira-Flores, D.; Chatzinikolaou, E.; Chatzigeorgiou, G.; et al. A marine biodiversity observation network for genetic monitoring of hard-bottom communities (ARMS-MBON). Front. Mar. Sci. 2020, 7, 1–9. [Google Scholar] [CrossRef]
- Zimmerman, T.L.; Martin, J.W. Artificial reef matrix structures (Arms): An inexpensive and effective method for collecting coral reef-associated invertebrates. Gulf Caribb. Res. 2004, 16, 59–64. [Google Scholar] [CrossRef]
- Smithsonian Global ARMS Program. Available online: https://naturalhistory.si.edu/research/global-arms-program (accessed on 4 November 2021).
- Anderson, M.J.; Underwood, A.J. Effects of substratum on the recruitment and development of an intertidal estuarine fouling assemblage. J. Exp. Mar. Biol. Ecol. 1994, 184, 217–236. [Google Scholar] [CrossRef]
- Guerra-Castro, E.J.; Cruz-Motta, J.J. Ecology of fouling assemblages associated with mangrove’s roots: An artificial substrate for manipulative experiments. J. Exp. Mar. Biol. Ecol. 2014, 457, 31–40. [Google Scholar] [CrossRef]
- Glasby, T.M.; Connell, S.D.; Holloway, M.G.; Hewitt, C.L. Nonindigenous biota on artificial structures: Could habitat creation facilitate biological invasions? Mar. Biol. 2007, 151, 887–895. [Google Scholar] [CrossRef]
- Menge, B.A.; Foley, M.M.; Pamplin, J.; Murphy, G.; Pennington, C. Supply-side ecology, barnacle recruitment, and rocky intertidal community dynamics: Do settlement surface and limpet disturbance matter? J. Exp. Mar. Biol. Ecol. 2010, 392, 160–175. [Google Scholar] [CrossRef]
- Menge, B.A.; Ashkenas, L.R.; Matson, A. Use of artificial holes in studying community development in cryptic marine habitats in a tropical rocky intertidal region. Mar. Biol. 1983, 142, 129–142. [Google Scholar] [CrossRef]
- Warwick, R.M.; Clarke, K.R. New “biodiversity” measures reveal a decrease in taxonomic distinctness with increasing stress. Mar. Ecol. Prog. Ser. 1995, 129, 301–305. [Google Scholar] [CrossRef] [Green Version]
- Leray, M.; Knowlton, N. DNA barcoding and metabarcoding of standardized samples reveal patterns of marine benthic diversity. Proc. Natl. Acad. Sci. USA 2015, 112, 2076–2081. [Google Scholar] [CrossRef] [Green Version]
- Al-Rshaidat, M.M.D.; Snider, A.; Rosebraugh, S.; Devine, A.M.; Devine, T.D.; Plaisance, L.; Knowlton, N.; Leray, M. Deep COI sequencing of standardized benthic samples unveils overlooked diversity of Jordanian coral reefs in the northern Red Sea. Genome 2016, 59, 724–737. [Google Scholar] [CrossRef]
- Ransome, E.; Geller, J.B.; Timmers, M.; Leray, M.; Mahardini, A.; Sembiring, A.; Collins, A.G.; Meyer, C.P. The importance of standardization for biodiversity comparisons: A case study using autonomous reef monitoring structures (ARMS) and metabarcoding to measure cryptic diversity on Mo’orea coral reefs, French Polynesia. PLoS ONE 2017, 12, e0175066. [Google Scholar] [CrossRef] [Green Version]
- Martínez-López, O. La taxonomía integral y su importancia para la conservación. Cienc. Conserv. 2015, 6, 54–64. [Google Scholar]
- Felder, D.L.; Camp, D.K. Gulf of Mexico: Origin, Waters, and Biota; Texas A&M University Press: College Station, TX, USA, 2009; Volume 1, p. 1393. [Google Scholar]
- De la Barreda, B.; Metcalfe, S.E.; Boyd, D.S. Precipitation regionalization, anomalies and drought occurrence in the Yucatan Peninsula, Mexico. Int. J. Climatol. 2020, 40, 4541–4555. [Google Scholar] [CrossRef]
- Santander-Monsalvo, J.; Espejel, I.; Ortiz-Lozano, L. Distribution, uses, and anthropic pressures on reef ecosystems of Mexico. Ocean Coast. Manag. 2018, 165, 39–51. [Google Scholar] [CrossRef]
- Ardisson, P.L.; May-Kú, M.A.; Herrera-Dorantes, M.T.; Arellano-Guillermo, A. El sistema arrecifal mesoamericano-méxico: Consideraciones para su designación como zona marítima especialmente sensible. Hidrobiologica 2011, 21, 261–280. [Google Scholar]
- Tunnel, J.W. Natural versus human impacts to southern Gulf of Mexico coral reef resources. In Proceedings of the 7th International Coral Reef Symposium—UOG Station, Guam, Micronesia, 22–27 June 1992; Volume 1, pp. 300–306. [Google Scholar]
- Tunnell, J.W.; Chávez, E.A.; Withers, K.; Earle, S. Coral Reefs of the Southern Gulf of Mexico; Texas A&M Universsity Press: Corpus Christi, TX, USA, 2007. [Google Scholar]
- NOAA Autonomous Reef Monitoring Structures (ARMS) Overview. Available online: https://naturalhistory.si.edu/research/global-arms-program#:~:text=What%20is%20an%20Autonomous%20Reef,sample%20without%20destroying%20natural%20habitat (accessed on 4 November 2021).
- Palomino-Alvarez, L.A.; Castillo-Cupul, R.E.; Suárez-Mozo, N.Y.; Ortigosa, D.; Paz-Ríos, C.E.; Cervantes-Campero, G.; Muciño-Reyes, M.; Homá-Canché, P.; Hernández-Díaz, Y.Q.; Sotelo, R.; et al. Autonomous Reef Monitoring Structures (ARMS). Benthic Marine Fauna Processing Manual. 2021. Available online: https://zenodo.org/record/5655251 (accessed on 4 November 2021).
- Burke, L.; Maidens, J. Caribbean Reefs at Risk Threat Index (Polygon); World Resources Institute: Washington DC, USA, 2004; Available online: https://databasin.org/datasets/0363d08f572d45fdbb1e675a08a52545/ (accessed on 5 March 2021).
- Clarke, K.R.; Warwick, R.M. A taxonomic distinctness index and its statistical properties. J. Appl. Ecol. 1998, 35, 523–531. [Google Scholar] [CrossRef]
- Warwick, R.M.; Clarke, K.R. Taxonomic distinctness and environmental assessment. J. Appl. Ecol. 1998, 35, 532–543. [Google Scholar] [CrossRef]
- Warwick, R.M.; Somerfield, P.J. All animals are equal, but some animals are more equal than others. J. Exp. Mar. Biol. Ecol. 2008, 366, 184–186. [Google Scholar] [CrossRef]
- OBIS Ocean Biodiversity Information System. Available online: www.iobis.org (accessed on 6 May 2021).
- Horton, T.; Gofas, S.; Kroh, A.; Poore, G.C.B.; Read, G.; Rosenberg, G.; Stöhr, S.; Bailly, N.; Boury-Esnault, N.; Brandão, S.N.; et al. Improving nomenclatural consistency: A decade of experience in the world register of marine species. Eur. J. Taxon. 2017, 389, 1–24. [Google Scholar] [CrossRef]
- Palomino-Alvarez, L.A.; Vital, X.G.; Suárez-Mozo, N.Y.; Castillo-Cupul, R.; Ugalde, D.; Cervantes-Campero, G.; Muciño-Reyes, M.; Homá-Canché, P.; Hernández-Díaz, Y.Q.; Sotelo, R.; et al. Matrix Aggregation of Species of Phyla Annelida (Polychaeta), Mollusca, Arthropoda (Decapoda, Stomatopoda, Amphipoda, and Chelicerata), and Echinodermata Registered of the Caribbean Sea and Gulf of Mexico Region by OCEAN Biodiversity Information Systems o. 2021. Available online: https://zenodo.org/record/5525313#.YYT1txwRVPY (accessed on 4 November 2021).
- Palomino-Alvarez, L.A.; Vital, X.G.; Suárez-Mozo, N.Y.; Castillo-Cupul, R.; Ugalde, D.; Cervantes-Campero, G.; Muciño-Reyes, M.; Homá-Canché, P.; Hernández-Díaz, Y.Q.; Sotelo, R.; et al. Database of Incidence per Reef and Región of the Research “Evaluation of the Use of Autonomous Reef Monitoring Structures (ARMS) for Capturing the Biological Diversity of Two Coral Reefs in the Yucatán Península, México”. 2021. Available online: https://zenodo.org/record/5525272#.YYT2hRwRVPY (accessed on 4 November 2021).
- Krebs, C.J. Estimating density: Quadrats counts. In Ecological Methodology; Addison-Wesley Educational Publishers, Inc.: Boston, MA, USA, 1999; p. 620. ISBN 032-1021-738. [Google Scholar]
- Kohler, K.; Gill, S. Coral point count with excel extensions (CPCe): A visual basic program for the determination of coral and substrate coverage using random point count methodology. Comput. Geosci. 2006, 32, 1259–1269. [Google Scholar] [CrossRef]
- Palomino-Alvarez, L.A.; Vital, X.G.; Suárez-Mozo, N.Y.; Castillo-Cupul, R.; Ugalde, D.; Cervantes-Campero, G.; Muciño-Reyes, M.; Homá-Canché, P.; Hernández-Díaz, Y.Q.; Sotelo, R.; et al. Abundance of Benthic Sessile Fauna Associated with ARMS from the Research “Evaluation of the use of Autonomous Reef Monitoring Structures (ARMS) for Capturing the Biological Diversity of Two Coral Reefs in the Yucatán Península, México” Using the Point Intersection Method (400 Points per Plate on Each Side) with the CPCe v4.1 Software (Coral Point Count with Excel extension v4.1). 2021. Available online: https://zenodo.org/record/5525336#.YYT3nhwRVPY (accessed on 4 November 2021).
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.; Somerfield, P.; Warwick, R. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation and Interpretation, 3rd ed.; Primer-E: Plymouth, UK, 2014; 262p. [Google Scholar]
- Palomino-Alvarez, L.A.; Vital, X.G.; Suárez-Mozo, N.Y.; Castillo-Cupul, R.; Ugalde, D.; Cervantes-Campero, G.; Muciño-Reyes, M.; Homá-Canché, P.; Hernández-Díaz, Y.Q.; Sotelo, R.; et al. Checklist of cryptbiont assemblages in coral reef of two subregions of the great caribbean sea using ARMS. Zenodo 2021. Available online: https://www.gbif.org/dataset/054cfdf9-ccff-4eb3-a815-ee934cf11e04 (accessed on 4 November 2021).
- Palomino-Alvarez, L.A.; Vital, X.G.; Suárez-Mozo, N.Y.; Castillo, R.; Ugalde, D.; Cervantes-Campero, G.; Muciño-Reyes, M.; Homá-Canché, P.; Hernández, Q.; Sotelo, R.; et al. Cryptobiont assemblages’s dataset of coral reefs using ARMS in B10 Reef and Mahahual Reef of the Great Caribbean. OBIS 2021. Available online: https://www.gbif.org/dataset/054cfdf9-ccff-4eb3-a815-ee934cf11e04, (accessed on 4 November 2021).
- Perkins, T.H. Chrysopetalum, Bhawania and two new genera of Chrysopetalidae (Polychaeta), principally from Florida. Proc. Biol. Soc. Wash. 1985, 98, 856–915. [Google Scholar]
- Read, G.; Fauchald, K. World Register of Polychaeta. Available online: http://www.marinespecies.org/polychaeta (accessed on 6 May 2021).
- Liñero, A.; Vasquez, G.R. Nereidae (polychaeta, errantia) del golfo de cariaco, venezuela. Bol. Inst. Oceanogr. Venez. Univ. Oriente 1979, 18, 3–12. [Google Scholar]
- Knight-Jones, P.; Giangrande, A. Two new species of an atypical group of pseudobranchiomma jones (polychaeta: Sabellidae). Hydrobiologia 2003, 496, 95–103. [Google Scholar] [CrossRef]
- Aguado, M.T.; San Martín, G. Syllidae (Polychaeta) from Lebanon with two new reports for the Mediterranean Sea. Cah. Biol. Mar. 2007, 48, 207–224. [Google Scholar]
- Katsanevakis, S.; Bogucarskis, K.; Gatto, F.; Vandekerkhove, J.; Deriu, I.; Cardoso, A.C. Building the European Alien Species Information Network (EASIN): A novel approach for the exploration of distributed alien species data. BioInvasions Rec. 2012, 1, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Zenetos, A.; Gofas, S.; Verlaque, M.; Çinar, M.E.; García Raso, J.G.; Bianchi, C.N.; Morri, C.; Azzurro, E.; Bilecenoglu, M.; Froglia, C.; et al. Alien species in the mediterranean sea by a contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part I. Spatial distribution. Mediterr. Mar. Sci. 2010, 11, 381–493. [Google Scholar] [CrossRef] [Green Version]
- Bamber, R.N.; El Nagar, A.; Arango, C.P. Pycnobase: World Pycnogonida Database. Available online: http://www.marinespecies.org/pycnobase%20on%202021-09-06 (accessed on 25 June 2021).
- Bravo, M.F.M.; Müller, H.G.; Arango, C.P.; Tigreros, P.; Melzer, R.R. Morphology of shallow-water sea spiders from the Colombian Caribbean. Spixiana 2009, 32, 9–34. [Google Scholar]
- Prata, J.; Lucena, R.A.; Lima, S.F.B.; Souza, J.W.S.; Christoffersen, M.L. Species richness of Pycnogonida and Echinodermata associated with the reef ecosystems of Morro de São Paulo on Tinharé Island in Northeastern Brazil. Int. J. Dev. Res. 2020, 10, 34943–34951. [Google Scholar]
- Paz-Ríos, C.E.; Simões, N.; Pech, D. Species richness and spatial distribution of benthic amphipods (crustacea: Peracarida) in the alacranes reef national park, gulf of Mexico. Mar. Biodivers. 2019, 49, 673–682. [Google Scholar] [CrossRef]
- Winfield, I.; Muciño-Reyes, M.; Cházaro-Olvera, S.; Ortiz, M.; Lozano-Aburto, M. Benthic amphipods (crustacea: Peracarida) from sisal reef system and coastline, northwest Yucatán Peninsula, Gulf of Mexico. Rev. Mex. Biodivers. 2020, 91, e903071. [Google Scholar] [CrossRef] [Green Version]
- Souza-Filho, J.F.; Souza, A.M.T.; Valério-Berardo, M.T. Six new species of the genus Chevalia Walker, 1904 (Amphipoda, Corophiidea, Chevaliidae) from Brazilian waters, with a key to world species of the genus. Zootaxa 2010, 2713, 25–51. [Google Scholar] [CrossRef]
- Kunkel, B.W. The amphipoda of bermuda. Trans. Connect. Acad. Arts Sci. 1910, 16, 1–116. [Google Scholar]
- Krapp-Schickel, T.; Vader, W. On some maerid genera (crustacea, amphipoda, maeridae) collected by the hourglass cruises (florida). Part 1: Genera anamaera, ceradocus, clessidra gen. nov., jerbarnia, maera, meximaera, with a key to world Ceradocus. J. Nat. Hist. 2009, 43, 2057–2086. [Google Scholar] [CrossRef]
- World List of Bryozoa. Available online: https://www.catalogueoflife.org/data/dataset/1081 (accessed on 8 October 2021).
- Leloup, E. Contribution à la connaissance de la faune des chitons de Villefranche-sur-Mer et des environs (France, Méditerranée) [Contribution to the knowledge of the fauna of chitons of Villefranche-sur-Mer and the surroundings (France, Mediterranean)]. Bull. Musée R. D’histoire Nat. Belg. Meded. Van Het K. Nat. Mus. Van België 1934, 10, 1–20. [Google Scholar]
- Costello, M.J. European Register of Marine Species: A Check- List of the Marine Species in Europe and a Bibliography of Guides to Their Identification; Muséum National d’Histoire Naturelle: Paris, France, 2001; Volume 50. [Google Scholar]
- Schuchert, P. Hydroids (Cnidaria, Hydrozoa) of the Danish expedition to the Kei Islands. Steenstrupia 2003, 27, 137–256. [Google Scholar]
- Galea, H.R.; Schuchert, P. Some thecate hydroids (Cnidaria: Hydrozoa) from off new caledonia collected during KANACONO and KANADEEP expeditions of the french tropical deep-sea benthos program. Eur. J. Taxon. 2019, 562, 1–70. [Google Scholar] [CrossRef]
- Billard, A. Note sur quelques espèces la plupart nouvelles de Synthécides et de Sertularides du Siboga. Bull. Soc. Zool. Fr. 1925, 49, 646–652. [Google Scholar]
- Van der Land, J. UNESCO-IOC Register of Marine Organisms (URMO). Available online: http://www.marinespecies.org/urmo/ (accessed on 7 June 2021).
- Newman, L.J.; Cannon, L.R.G. The genus Cycloporus (Platyhelminthes: Polycladida) from Australasian waters. Raffles Bull. Zool. 2002, 50, 287–299. [Google Scholar]
- Du Bois-Reymond, M.E. Polycladida from Curaçao and faunistically related regions. Stud. Fauna Curaçao Other Caribb. Isl. 1968, 26, 1–133. [Google Scholar]
- Quiroga, S.Y. Systematics and Taxonomy of Polyclad Flatworms with a Special Emphasis on the Morphology of the Nervous System. Ph.D. Thesis, University of New Hampshire, Durham, NH, USA, 2008. [Google Scholar]
- García, B.A. El Uso de ARMS (Autonomous Reef Monitoring Structures) Como Herramienta para el Estudio del Reclutamiento en Invertebrados Crípticos en La Perla del Golfo, Veracruz; Universidad Nacional Autónoma de México: Mexico City, Mexico, 2021. [Google Scholar]
- González-Gándara, C.; Solís-Marín, F.A.; de la Cruz-Francisco, V.; Granados-Barba, A.; Salas-Pérez, J.D.J.; Argüelles-Jiménez, J.; Escárcega-Quiroga, P.A. Riqueza y distribución de equinodermos en los arrecifes del norte y sur de Veracruz, México. Rev. Biol. Trop. 2015, 63, 183–193. [Google Scholar] [CrossRef]
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; Ferdaña, Z.A.; Finlayson, M.A.X.; Halpern, B.S.; Jorge, M.A.; Lombana, A.L.; Lourie, S.A.; et al. Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. BioScience 2007, 57, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Chamberlin, R.V. New polychaetous annelids from Laguna Beach, California. J. Entomol. Zool. Pomona Coll. 1919, 11, 1–23. [Google Scholar]
- Pearman, J.K.; Chust, G.; Aylagas, E.; Villarino, E.; Watson, J.R.; Chenuil, A.; Borja, A.; Cahill, A.E.; Carugati, L.; Danovaro, R.; et al. Pan-regional marine benthic cryptobiome biodiversity patterns revealed by metabarcoding autonomous Reef monitoring Structures. Mol. Ecol. 2020, 29, 4882–4897. [Google Scholar] [CrossRef] [PubMed]
- Plaisance, L.; Knowlton, N.; Paulay, G.; Meyer, C. Reef-associated crustacean fauna: Biodiversity estimates using semi-quantitative sampling and DNA barcoding. Coral Reefs 2009, 28, 977–986. [Google Scholar] [CrossRef] [Green Version]
- Plaisance, L.; Caley, M.J.; Brainard, R.E.; Knowlton, N. The diversity of coral reefs: What are we missing? PLoS ONE 2011, 6, e25026. [Google Scholar] [CrossRef] [Green Version]
- Engel, M.S.; Ceríaco, L.M.P.; Daniel, G.M.; Dellapé, P.M.; Löbl, I.; Marinov, M.; Reis, R.E.; Young, M.T.; Dubois, A.; Agarwal, I.; et al. The taxonomic impediment: A shortage of taxonomists, not the lack of technical approaches. Zool. J. Linn. Soc. 2021, 193, 381–387. [Google Scholar] [CrossRef]
- Ostrander, G.K.; Armstrong, K.M.; Knobbe, E.T.; Gerace, D.; Scully, E.P. Rapid transition in the structure of a coral reef community: The effects of coral bleaching and physical disturbance. Proc. Natl. Acad. Sci. USA 2000, 97, 5297–5302. [Google Scholar] [CrossRef] [Green Version]
- Rogers, C.S.; Miller, J. Permanent “phase shifts” or reversible declines in coral cover? Lack of recovery of two coral reefs in St. John, US Virgin Islands. Mar. Ecol. Prog. Ser. 2006, 306, 103–114. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | GMx | Cs | Phylum | Documented Distribution |
|---|---|---|---|---|
| Treptopale rudolphi Perkins, 1985 | ✓ | Annelida (Polychaeta) | Miami, USA [47], Caribbean [48] | |
| Rullierinereis cf. bahamensis (Hartmann-Schröder, 1958) | ✓ | Annelida (Polychaeta) | Bimini Islands, Bahamas [48], gulf of Cariaco, Venezuela [49] | |
| Pseudobranchiomma perkinsi Knight-Jones and Giangrande, 2003 | ✓ | Annelida (Polychaeta) | Florida, USA [50] | |
| Syllis bella (Chamberlin, 1919) | ✓ | ✓ | Annelida (Polychaeta) | California, USA [48], Mediterranean [51], recorded as an invasive species [52,53] |
| Ammothella spinifera Cole, 1904 | ✓ | Arthropoda (Chelicerata) | Pacific Ocean [54], Santa Marta and Cabo Arrecifes, Colombia [55], Morro Sao Paolo, Brasil [56] | |
| Aruga holmesi J.L. Barnard, 1955 | ✓ | Arthropoda (Amphipoda) | Gulf of Mexico [24], northern coast of Yucatan [57,58] | |
| Chevalia caetes Souza-Filho, Souza and Valério-Berardo, 2010 | ✓ | ✓ | Arthropoda (Amphipoda) | Penambuco, Brazil [59] |
| Clessidra tinkerensis (Kunkel, 1910) | ✓ | Arthropoda (Amphipoda) | Bermuda, West Florida, USA [60,61] | |
| Parasmittina bimucronata (Hincks, 1884) | ✓ | Bryozoa | Burma, Myanmar [62] | |
| Plumularia obliqua (Johnston, 1847) | ✓ | Cnidaria (Hydrozoa) | Villefranche-sur-Mer, France [63], Northwestern Atlantic [64] | |
| Geminella ceramensis (Billard, 1925) | ✓ | Cnidaria (Hydrozoa) | Philippines, Kei islands, Indonesia [65] and New Caledonia [66], Siboga [67], Western Central Pacific [68] | |
| Cycloporus variegatus Kato, 1934 | ✓ | Platyhelminthes | Australia [69] | |
| Pericelis cata Marcus and Marcus, 1968 | ✓ | Platyhelminthes | Curazao, Venezuela [70], Colombia [71] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palomino-Alvarez, L.A.; Vital, X.G.; Castillo-Cupul, R.E.; Suárez-Mozo, N.Y.; Ugalde, D.; Cervantes-Campero, G.; Muciño-Reyes, M.R.; Homá-Canché, P.; Hernández-Díaz, Y.Q.; Sotelo-Casas, R.; et al. Evaluation of the Use of Autonomous Reef Monitoring Structures (ARMS) for Describing the Species Diversity of Two Coral Reefs in the Yucatan Peninsula, Mexico. Diversity 2021, 13, 579. https://doi.org/10.3390/d13110579
Palomino-Alvarez LA, Vital XG, Castillo-Cupul RE, Suárez-Mozo NY, Ugalde D, Cervantes-Campero G, Muciño-Reyes MR, Homá-Canché P, Hernández-Díaz YQ, Sotelo-Casas R, et al. Evaluation of the Use of Autonomous Reef Monitoring Structures (ARMS) for Describing the Species Diversity of Two Coral Reefs in the Yucatan Peninsula, Mexico. Diversity. 2021; 13(11):579. https://doi.org/10.3390/d13110579
Chicago/Turabian StylePalomino-Alvarez, Lilian A., Xochitl G. Vital, Raúl E. Castillo-Cupul, Nancy Y. Suárez-Mozo, Diana Ugalde, Gabriel Cervantes-Campero, María R. Muciño-Reyes, Pedro Homá-Canché, Yoalli Quetzalli Hernández-Díaz, Rosa Sotelo-Casas, and et al. 2021. "Evaluation of the Use of Autonomous Reef Monitoring Structures (ARMS) for Describing the Species Diversity of Two Coral Reefs in the Yucatan Peninsula, Mexico" Diversity 13, no. 11: 579. https://doi.org/10.3390/d13110579
APA StylePalomino-Alvarez, L. A., Vital, X. G., Castillo-Cupul, R. E., Suárez-Mozo, N. Y., Ugalde, D., Cervantes-Campero, G., Muciño-Reyes, M. R., Homá-Canché, P., Hernández-Díaz, Y. Q., Sotelo-Casas, R., García-González, M., Avedaño-Peláez, Y. A., Hernández-González, A., Paz-Ríos, C. E., Lizaola-Guillermo, J. M., García-Venegas, M., Dávila-Jiménez, Y., Ortigosa, D., Hidalgo, G., ... Guerra-Castro, E. J. (2021). Evaluation of the Use of Autonomous Reef Monitoring Structures (ARMS) for Describing the Species Diversity of Two Coral Reefs in the Yucatan Peninsula, Mexico. Diversity, 13(11), 579. https://doi.org/10.3390/d13110579