
Genome-Wide SNP Analysis of Male and Female Rice Field Frogs, Hoplobatrachus rugulosus, Supports a Non-Genetic Sex Determination System






Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimens and DNA Extraction
2.2. DArT Sequencing, Genotyping and Analysis
2.3. Chromosome Preparation
2.4. Microsatellite Repeat Motifs, Telomeric (TTAGGG)n FISH Mapping
3. Results
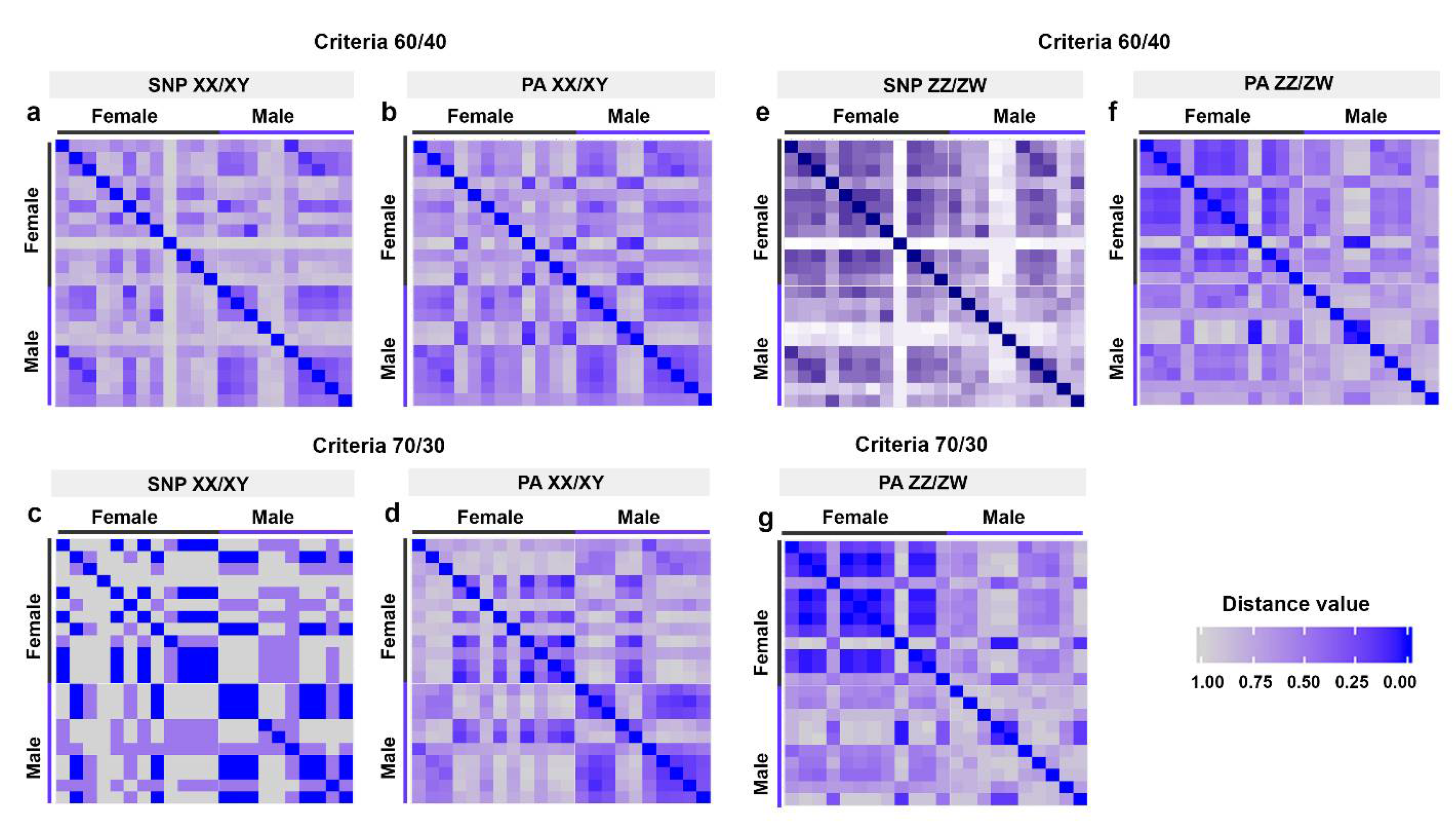
3.1. Determination of Sex System and Identification of Sex-Linked Loci in Rice Field Frog
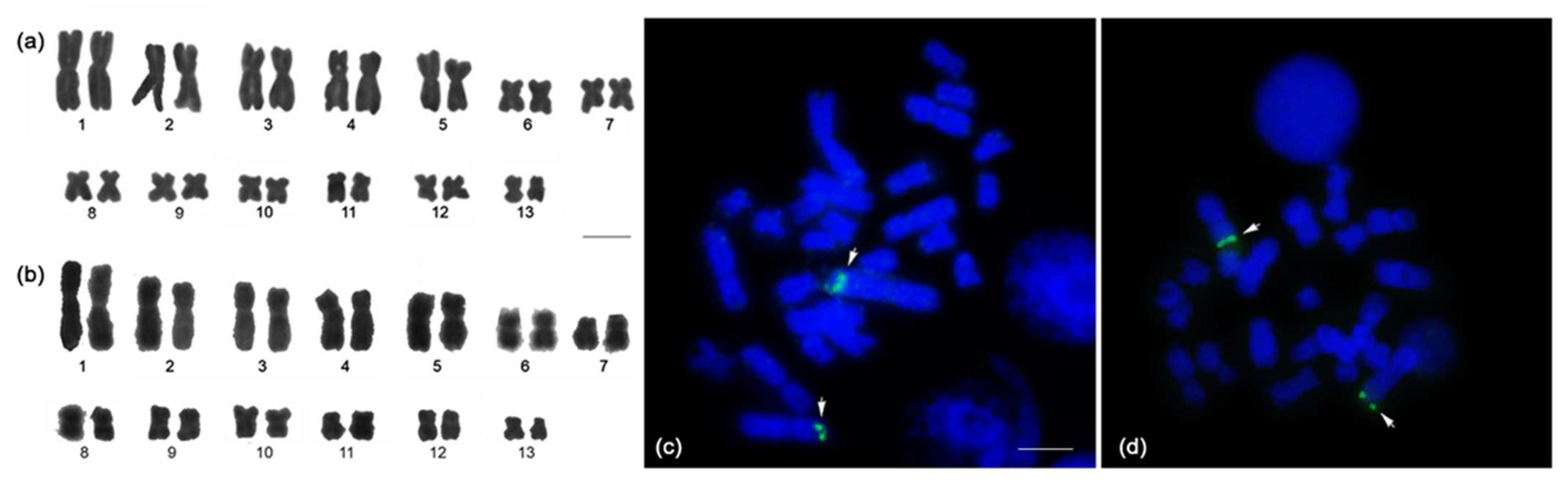
3.2. Karyotype
3.3. Chromosomal Locations of the Telomeric (TTAGGG)n Sequences and Microsatellite Repeat Motifs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- King, H.D. Studies on sex-determination in amphibians. II. Biol. Bull. 1909, 16, 27–43. [Google Scholar] [CrossRef] [Green Version]
- Witschi, E. Studies on sex differentiation and sex determination in amphibians. IV. The geographical distribution of the sex races of the European grass frog (Rana temporaria, L.). A contribution to the problem of the evolution of sex. J. Exp. Zool. 1930, 56, 149–165. [Google Scholar] [CrossRef]
- Tamschick, S.; Rozenblut-Koscisty, B.; Ogielska, M.; Lehmann, A.; Lymberakis, P.; Hoffmann, F.; Lutz, I.; Kloas, W.; Stock, M. Sex reversal assessments reveal different vulnerability to endocrine disruption between deeply diverged anuran lineages. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezaz, T.; Srikulnath, K.; Graves, J.A.M. Origin of amniote sex chromosomes: An ancestral super-sex chromosome, or common requirements. J. Hered. 2017, 108, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Miura, I. Sex determination and sex chromosomes in amphibia. Sex Dev. 2017, 11, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Miura, Y.; Qureshi, H.; Ryoo, C.; Dinenis, C.P.; Li, J.; Mandli, K.T.; Deodatis, G.; Bienstock, D.; Lazrus, H.; Morss, R. A methodological framework for determining an optimal coastal protection strategy against storm surges and sea level rise. Nat. Hazards 2021, 107, 1821–1843. [Google Scholar] [CrossRef]
- Lambert, M.; Skelly, D.K.; Ezaz, T. Sex-linked markers in the North American green frog (Rana clamitans) developed using DArTseq provide early insight into sex chromosome evolution. BMC Genom. 2016, 17, 844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sopniewski, J.; Shams, F.; Scheele, B.C.; Kefford, B.J.; Ezaz, T. Identifying sex-linked markers in Litoria aurea: A novel approach to understanding sex chromosome evolution in an amphibian. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wessels, S.; Krause, I.; Floren, C.; Schütz, E.; Beck, J.; Knorr, C. ddRADseq reveals determinants for temperature-dependent sex reversal in Nile tilapia on LG23. BMC Genom. 2017, 18, 531. [Google Scholar] [CrossRef] [Green Version]
- Koomgun, T.; Laopichienpong, N.; Singchat, W.; Panthum, T.; Phatcharakullawarawat, R.; Kraichak, E.; Sillapaprayoon, S.; Ahmad, S.F.; Muangmai, N.; Peyachoknagul, S.; et al. Genome complexity reduction high-throughput genome sequencing of green iguana (Iguana iguana) reveal a paradigm shift in understanding sex-chromosomal linkages on homomorphic X and Y sex chromosomes. Front. Genet. 2020, 11, 1217. [Google Scholar] [CrossRef]
- Laopichienpong, N.; Kraichak, E.; Singchat, W.; Sillapaprayoon, S.; Muangmai, N.; Suntrarachun, S.; Baicharoen, S.; Peyachoknagul, S.; Chanhome, L.; Ezaz, T.; et al. Genome-wide SNP analysis of Siamese cobra (Naja kaouthia) reveals the molecular basis of transitions between Z and W sex chromosomes and supports the presence of an ancestral super-sex chromosome in amniotes. Genomics 2021, 113, 624–636. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.H.M.; Panthum, T.; Ponjarat, J.; Laopichiengpong, N.; Kraichak, E.; Singchat, W.; Muangmai, N.; Peyachoknagul, S.; Na-Nakorn, U.; Srikulnath, K. An investigation of ZZ/ZW and XX/XY sex determination systems in North African catfish (Clarias gariepinus, Burchell 1822). Front. Genet. 2021, 11, 1719. [Google Scholar] [CrossRef]
- Nguyen, D.H.M.; Ponjarat, J.; Laopichienpong, N.; Kraichak, E.; Panthum, T.; Singchat, W.; Ahmad, S.F.; Muangmai, N.; Duengkae, P.; Peyachoknagul, S.; et al. Genome-wide SNP analysis suggests male heterogamety in bighead catfish (Clarias macrocephalus). Aquaculture 2021, 545, 737005. [Google Scholar] [CrossRef]
- Frost, D.R. Amphibian Species of the World: An Online Reference. American Museum of Natural History, New York, USA. Version 6.0 (Date of Access). Electronic Database. 2016. Available online: http://research.amnh.org/herpetology/amphibia/index.html (accessed on 9 June 2021).
- Alam, M.S.; Igawa, T.; Khan, M.M.R.; Islam, M.M.; Kuramoto, M.; Matsui, M.; Kurabayashi, A.; Sumida, M. Genetic divergence and evolutionary relationships in six species of genera Hoplobatrachus and Euphlyctis (Amphibia: Anura) from Bangladesh and other Asian countries revealed by mitochondrial gene sequences. Mol. Phylogenet. Evol. 2008, 48, 515–527. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhang, J.Y.; Zheng, R.Q.; Yu, B.G.; Yang, G. Complete nucleotide sequence and gene organization of the mitochondrial genome of Paa spinosa (Anura: Ranoidae). Gene 2009, 447, 86–96. [Google Scholar] [CrossRef]
- Chen, G.; Wang, B.; Liu, J.; Xie, F.; Jiang, J. Complete mitochondrial genome of Nanorana pleskei (Amphibia: Anura: Dicroglossidae) and evolutionary characteristics. Curr. Zool. 2011, 57, 785–805. [Google Scholar] [CrossRef]
- Pyron, R.A.; Wiens, J.J. A large-scale phylogeny of amphibia including over 2800 species, and a revised classification of extant frogs, salamanders, and caecilians. Mol. Phylogenet. Evol. 2011, 61, 543–583. [Google Scholar] [CrossRef]
- Wiegmann, A.F.A. Amphibian. 1834, pp. 433–522. Available online: http://www.marinespecies.org/aphia.php?p=taxdetails&id=1458139 (accessed on 9 June 2021).
- Diesmos, A.; Van-Dijk, P.P.; Inger, R.; Iskandar, D.; Lau, M.W.N.; Ermi, Z.; Shunqing, L.; Baorong, G.; Kuangyang, L.; Zhigang, Y.; et al. Hoplobatrachus rugulosus . In The IUCN Red List of Threatened Species. Version 2014.3; Internet References; 2004; Available online: http://www.iucnredlist.org/details/full/58300/0 (accessed on 9 June 2021).
- Meesawat, S.; Kitana, N.; Kitana, J. Hematology of wild caught Hoplobatrachus rugulosus in Northern Thailand. Asian Herpetol. Res. 2016, 7, 131–138. [Google Scholar] [CrossRef]
- Fei, L.; Ye, C.Y.; Jiang, J.P. Colored Atlas of Chinese Amphibians and Their Distributions; Sichuan Science and Technology Publishing House: Chengdu, China, 2012; pp. 572–573. [Google Scholar]
- Schmalz, T.D.; Zug, G.R. Observations on geographic variation in the Asian frog, Hoplobatrachus rugulosus (Anura: Ranidae). Hamadryad 2002, 27, 90–98. [Google Scholar]
- Bain, R.H.; Truong, N.Q. Herpetofaunal diversity of Ha Giang province in northeastern Vietnam, with descriptions of two new species. Am. Mus. Novit. 2004, 3453, 1–42. [Google Scholar] [CrossRef]
- Tokur, B.; Gürbüz, R.D.; Özyurt, G. Nutritional composition of frog (Rana esculanta) waste meal. Bioresour. Technol. 2008, 99, 1332–1338. [Google Scholar] [CrossRef] [PubMed]
- Ruamthum, W.; Visetson, S.; Milne, J.R.; Bullangpoti, V. Effect of glyphosate-based herbicide on acetylcholinesterase activity in tadpoles, Hoplobatrachus rugulosus. Commun. Agric. Appl. Biol. Sci. 2011, 76, 923–930. [Google Scholar] [PubMed]
- Hasan, M.; Kuramoto, M.; Islam, M.M.; Alam, M.S.; Khan, M.M.R.; Sumida, M. A new species of genus Hoplobatrachus (Anura, Dicroglossidae) from the coastal belt of Bangladesh. Zootaxa 2012, 3312, 45–48. [Google Scholar] [CrossRef]
- Pansook, A.; Khonsue, W.; Piyapattanakorn, S.; Pariyanonth, P. Phylogenetic relationships among Hoplobatrachus rugulosus in Thailand as inferred from mitochondrial DNA sequences of the cytochrome-b gene (Amphibia, Anura, Dicroglossidae). Zool. Sci. 2012, 29, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Trachantong, W.; Promya, J.; Saenphet, S.; Saenphet, K. Effects of atrazine herbicide on metamorphosis and gonadal development of Hoplobatrachus rugulosus. Maejo Int. J. Sci. 2013, 7, 60–71. [Google Scholar]
- Yu, T.; Yang, G.; Busam, M.; Deng, Y. Plasticity in metamorphic traits of rice field frog (Rana limnocharis) tadpoles: The interactive effects of rearing temperature and food level. Asian Herpetol. Res. 2016, 7, 265–270. [Google Scholar] [CrossRef]
- Kusrini, M.D. Edible Frog Harvesting in Indonesia: Evaluating Its Impact and Ecological Context. Ph.D. Thesis, School of Tropical Biology, James Cook University, Douglas, Australia, 2005. [Google Scholar]
- Ding, G.H.; Lin, Z.H.; Fan, X.L.; Ji, X. The combined effects of food supply and larval density on survival, growth and metamorphosis of Chinese tiger frog (Hoplobatrachus rugulosa) tadpoles. Aquaculture 2015, 435, 398–402. [Google Scholar] [CrossRef]
- Sheridan, J.A. Ecology and behavior of Polypedates leucomystax (Anura: Rhacophoridae) in Northeast Thailand. Herpetol. Rev. 2008, 39, 165–169. [Google Scholar]
- Moreira, C.R.; Henriques, M.B.; Ferreira, C.M. Frog farms as proposed in agribusiness aquaculture: Economic viability based in feed conversion. Bol. Inst. Pesca. 2013, 39, 389–399. [Google Scholar]
- Graham, K.M.; Kouba, A.J.; Langhorne, C.J.; Marcec, R.M.; Willard, S.T. Biological sex identification in the endangered dusky gopher frog (Lithobates sevosa): A comparison of body size measurements, secondary sex characteristics, ultrasound imaging, and urinary hormone analysis methods. Reprod. Biol. Endocrinol. 2016, 14, 41. [Google Scholar] [CrossRef] [Green Version]
- Hillis, D.M.; Green, D.M. Evolutionary changes of heterogametic sex in the phylogenetic history of amphibians. J. Evol. Biol. 1990, 3, 49–64. [Google Scholar] [CrossRef]
- Sarre, S.D.; Ezaz, T.; Georges, A. Transitions between sex-determining systems in reptiles and amphibians. Annu. Rev. Genom. Hum. Genet. 2011, 12, 391–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbott, J.K.; Nordén, A.K.; Hansson, B. Sex chromosome evolution: Historical insights and future perspectives. Proc. R. Soc. B Biol. Sci. 2017, 284, 20162806. [Google Scholar] [CrossRef] [Green Version]
- Conte, M.; Zucchi, M.I.; Andrade, G.V.; Souza, A.P.; Recco-Pimentel, S.M. Study of closely related species within the Physalaemus cuvieri group (Anura): Contribution of microsatellite markers. Genet. Mol. Res. 2011, 10, 1434–1444. [Google Scholar] [CrossRef]
- Miura, I.; Ohtani, H.; Nakamura, M.; Ichikawa, Y.; Saitoh, K. The origin and differentiation of the heteromorphic sex chromosomes Z, W, X, and Y in the frog Rana rugosa, inferred from the sequences of a sex-linked gene, ADP/ATP translocase. Molec. Biol. Evol. 1998, 15, 1612–1619. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, N.; Vuille, Y.; Loman, J.; Perrin, N. Sex-chromosome differentiation and ‘sex races’ in the common frog (Rana temporaria). Proc. R. Soc. B. 2015, 282, 20142726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, N.; Vuille, Y.; Brelsford, A.; Merila, J.; Perrin, N. The genetic contribution to sex determination and number of sex chromosomes vary among populations of common frogs (Rana temporaria). Heredity 2016, 117, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Phuge, S.K. High temperatures influence sexual development differentially in male and female tadpoles of the Indian skipper frog, Euphlyctis cyanophlyctis. J. Biosci. 2017, 42, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Traijitt, T.; Kitana, N.; Kitana, J. Pattern of gonadal sex differentiation in the rice field frog Hoplobatrachus rugulosus (Anura: Dicroglossidae). Zool. Stud. 2020, 59, e51. [Google Scholar] [CrossRef] [PubMed]
- Chaichorfa, E.; Duangjai, A. Effects of temperature on sex determination on Thai native frog, Rana Rugulosa (Wiegmann) culture in northern part of Thailand. In Proceedings of the Agricultural Conference, Khon Kaen University, Khon Kaen, Thailand, 26–27 January 2009; pp. 182–184. [Google Scholar]
- Tang, Y.; Chen, Z.Q.; Lin, Y.F.; Chen, J.Y.; Ding, G.H.; Ji, X. The combined effects of temperature and aromatase inhibitor on metamorphosis, growth, locomotion, and sex ratio of tiger frog (Hoplobatrachus rugulosus) tadpoles. PeerJ 2020, 8, e8834. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Zhang, J.; Li, P.; Zheng, R.; Shao, C. Do cryptic species exist in Hoplobatrachus rugulosus? An examination using four nuclear genes, the Cyt b gene and the complete MT genome. PLoS ONE 2015, 10, e0124825. [Google Scholar] [CrossRef]
- Kitano, J.; Mori, S.; Peichel, C.L. Sexual dimorphism in the external morphology of the threespine stickleback (Gasterosteus aculeatus). Copeia 2007, 2007, 336–349. [Google Scholar] [CrossRef]
- Wyneken, J.; Epperly, S.P.; Crowder, L.B.; Vaughan, J.; Blair Esper, K. Determining sex in posthatchling loggerhead sea turtles using multiple gonadal and accessory duct characteristics. Herpetologica 2007, 63, 19–30. [Google Scholar] [CrossRef]
- Haczkiewicz, K.; Ogielska, M. Gonadal sex differentiation in frogs: How testes become shorter than ovaries. Zool. Sci. 2013, 30, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Supikamolseni, A.; Ngaoburanawit, N.; Sumontha, M.; Chanhome, L.; Suntrarachun, S.; Peyachoknagul, S.; Srikulnath, K. Molecular barcoding of venomous snakes and species-specific multiplex PCR assay to identify snake groups for which antivenom is available in Thailand. Genet. Mol. Res. 2015, 14, 13981–13997. [Google Scholar] [CrossRef]
- Jaccoud, D.; Peng, K.; Feinstein, D.; Kilian, A. Diversity arrays: A solid state technology for sequence information independent genotyping. Nucleic Acids Res. 2001, 29, e25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilian, A.; Wenzl, P.; Huttner, E.; Carling, J.; Xia, L.; Blois, H.; Caig, V.; Heller-Uszynska, K.; Jaccoud, D.; Hopper, C.; et al. Diversity arrays technology: A generic genome profiling technology on open platforms. Methods Mol. Biol. 2012, 888, 67–89. [Google Scholar] [CrossRef]
- Ren, R.; Ray, R.; Li, P.; Xu, J.; Zhang, M.; Liu, G.; Yao, X.; Kilian, A.; Yang, X. Construction of a high-density DArTseq SNP-based genetic map and identification of genomic regions with segregation distortion in a genetic population derived from a cross between feral and cultivated-type watermelon. Mol. Genet. Genom. 2015, 290, 1457–1470. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Sven, K.; Klaus, R. HapEstXXR: Multi-Locus Stepwise Regression. 2019. Available online: https://rdrr.io/cran/HapEstXXR/man/coding.baseline.allele.html (accessed on 9 June 2021).
- Chew, J.S.; Oliveira, C.; Wright, J.M.; Dobson, M.J. Molecular and cytogenetic analysis of the telomeric (TTAGGG)n repetitive sequences in the Nile tilapia, Oreochromis niloticus (Teleostei: Cichlidae). Chromosoma 2002, 111, 45–52. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Bertollo, L.A.C. Initial steps in XY chromosome differentiation in Hoplias malabaricus and the origin of an X1 X2 Y sex chromosome system in this fish group. Heredity 2010, 105, 554–561. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, K.; O’Meally, D.; Azad, B.; Georges, A.; Sarre, S.D.; Graves, J.A.M.; Matsuda, Y.; Ezaz, T. Amplification of microsatellite repeat motifs is associated with the evolutionary differentiation and heterochromatinization of sex chromosomes in Sauropsida. Chromosoma 2016, 125, 111–123. [Google Scholar] [CrossRef]
- Singchat, W.; O’Connor, R.E.; Tawichasri, P.; Suntronpong, A.; Sillapaprayoon, S.; Suntrarachun, S.; Muangmai, N.; Baicharoen, S.; Peyachoknagul, S.; Srikulnath, K.; et al. Chromosome map of the Siamese cobra: Did partial synteny of sex chromosomes in the amniote represent “a hypothetical ancestral super-sex chromosome” or random distribution? BMC Genom. 2018, 19, 939. [Google Scholar] [CrossRef]
- Singchat, W.; Ahmad, S.F.; Laopichienpong, N.; Suntronpong, A.; Panthum, T.; Griffin, D.K.; Srikulnath, K. Snake W sex chromosome: The shadow of ancestral amniote super-sex chromosome. Cells 2020, 9, 2386. [Google Scholar] [CrossRef]
- Srikulnath, K.; Azad, B.; Singchat, W.; Ezaz, T. Distribution and amplification of interstitial telomeric sequences (ITSs) in Australian dragon lizards support frequent chromosome fusions in Iguania. PLoS ONE 2019, 14, e0212683. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, Y.; Chapman, V.M. Application of fluorescence in situ hybridization in genome analysis of the mouse. Electrophoresis 1995, 16, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Srikulnath, K.; Nishida, C.; Matsubara, K.; Uno, Y.; Thongpan, A.; Suputtitada, S.; Apisitwanich, S.; Matsuda, Y. Karyotypic evolution in squamate reptiles: Comparative gene mapping revealed highly conserved linkage homology between the butterfly lizard (Leiolepis reevesii rubritaeniata, Agamidae, Lacertilia) and the Japanese four-striped rat snake (Elaphe quadrivirgata, Colubridae, Serpentes). Chromosome Res. 2009, 17, 975. [Google Scholar] [CrossRef] [PubMed]
- Furman, B.L.S.; Evans, B.J. Sequential turnovers of sex chromosomes in African clawed frogs (Xenopus) suggest some genomic regions are good at sex determination. G3 2016, 6, 3625–3633. [Google Scholar] [CrossRef] [Green Version]
- Lambert, M.R.; Smylie, M.S.; Roman, A.J.; Freidenburg, L.K.; Skelly, D.K. Sexual and somatic development of wood frog tadpoles along a thermal gradient. J. Exp. Zool. A Ecol. Integr. Physiol. 2018, 329, 72–79. [Google Scholar] [CrossRef]
- Chang, C.; Wang, A.; Guo, P. The karyotype of the frog (Rana tigrina rugulosa). Zool. Res. 1984, 5, 33–38. [Google Scholar]
- Supaprom, T.; Baimai, A. Characterization of C-banded mitotic chromosomes in four species of anuran amphibians from Thailand. Amphib. Reptil. 2005, 26, 367–371. [Google Scholar] [CrossRef] [Green Version]
- Donsakul, T. Karyotype from liver cells of seven amphibians (amphibian, Anura) from Thailand. Scimsu 2009, 28, 162–170. [Google Scholar]
- Alam, M.S.; Islam, M.M.; Khan, M.M.R.; Hasan, M.; Wanichanon, R.; Sumida, M. Postmating isolation in six species of three genera (Hoplobatrachus, Euphlyctis and Fejervarya) from family Dicroglossidae (Anura) with special reference to spontaneous production of allotriploids. Zool. Sci. 2012, 29, 743–752. [Google Scholar] [CrossRef]
- Suttichaiya, A.; Khammanichanh, A.; Patawang, I.; Sriuttha, M.; Tanamtong, A.; Neeratanaphan, L. Chromosome aberrations of East Asian bullfrog (Hoplobatrachus rugulosus) around a gold mine area with arsenic contamination. Environmentasia 2016, 9, 67–76. [Google Scholar] [CrossRef]
- Tengjaroenkul, B.; Intamat, S.; Sriuttha, M.; Tanomtong, A.; Neeratanaphan, L. Cytotoxicity evaluation of the East Asian bullfrog (Hoplobatrachus rugulosus) in an agricultural area affected by chlorpyrifos. Cytologia 2017, 82, 175–181. [Google Scholar] [CrossRef]
- Saba, N.; Tripathi, N.K. Preliminary cytogenetic study and report of ZZ/ZW sex chromosomes in the bullfrog, Hoplobatrachus tigerinus (Anura, Amphibia) from high altitude area of Jammu and Kashmir, India. Nucleus 2014, 57, 55–59. [Google Scholar] [CrossRef]
- Srikulnath, K.; Uno, Y.; Nishida, C.; Ota, H.; Matsuda, Y. Karyotype reorganization in the Hokou gecko (Gekko hokouensis, Gekkonidae): The process of microchromosome disappearance in Gekkota. PLoS ONE 2015, 10, e0134829. [Google Scholar] [CrossRef] [PubMed]
- Gamble, T.; Coryell, J.; Ezaz, T.; Lynch, J.; Scantlebury, D.P.; Zarkower, D. Restriction site-associated DNA sequencing (RAD-seq) reveals an extraordinary number of transitions among gecko sex-determining systems. Mol. Biol. Evol. 2015, 32, 1296–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shams, F.; Dyer, F.; Thompson, R.; Duncan, R.P.; Thiem, J.D.; Kilian, A.; Ezaz, T. Application of DArT seq derived SNP tags for comparative genome analysis in fishes; An alternative pipeline using sequence data from a non-traditional model species, Macquaria ambigua. PLoS ONE 2019, 14, e0226365. [Google Scholar] [CrossRef] [PubMed]
- Diamanti-Kandarakis, E.; Bourguignon, J.P.; Giudice, L.C.; Hauser, R.; Prins, G.S.; Soto, A.M.; Zoeller, R.T.; Gore, A.C. Endocrine-disrupting chemicals: An endocrine society scientific statement. Endocr. Rev. 2009, 30, 293–342. [Google Scholar] [CrossRef]
- Hu, J.; Liu, J.; Lv, X.; Yu, L.; Li, J.; Lan, S.; Yang, Y. In situ assessment of genetic and epigenetic alterations in frog Rana plancyi and Rana limnocharis inhabiting aquatic ecosystems associated with Pb/Zn/Cu mining. Sci. Total Environ. 2021, 779, 146139. [Google Scholar] [CrossRef]
- Hallowell, E. Report upon the Reptilia of the North Pacific exploring expedition, under command of Capt. John Roger. Proc. Acad. Nat. Sci. Phila. 1860, 12, 1–9. [Google Scholar]
- Yu, B.G.; Zheng, R.Q.; Zhang, Y.; Liu, C.T. Geographic variation in body size and sexual size dimorphism in the giant spiny frog Paa spinosa (David, 1875) (Anura: Ranoidae). J. Nat. Hist. 2010, 44, 1729–1741. [Google Scholar] [CrossRef]
- Nakamura, M. Sex determination in amphibians. In Seminars in Cell & Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2009; Volume 20, pp. 271–282. [Google Scholar] [CrossRef]
- Rhen, T.; Schroeder, A.L. The genetics of thermosensitive sex determination. Temperature 2016, 4, 109–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, W.J.; Veltsos, P. The diversity and evolution of sex chromosomes in frogs. Genes 2021, 12, 483. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Pokorná, M.; Altmanová, M.; Kratochvíl, L. Cretaceous park of sex determination: Sex chromosomes are conserved across iguanas. Biol. Lett. 2014, 10, 20131093. [Google Scholar] [CrossRef]
- Acosta, A.; Martínez-Pacheco, M.L.; Díaz-Barba, K.; Porras, N.; Gutiérrez-Mariscal, M.; Cortez, D. Deciphering ancestral sex chromosome turnovers based on analysis of male mutation bias. Genome Biol. Evol. 2019, 11, 3054–3067. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.V.; Guzmán-Méndez, I.A.; Gamble, T.; Blumer, M.; Pinto, B.J.; Kratochvíl, L.; Rovatsos, M. Escaping the evolutionary trap? Sex chromosome turnover in basilisks and related lizards (Corytophanidae: Squamata). Biol. Lett. 2019, 15, 20190498. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| 60:40 Male:Female | 70:30 Male:Female | 80:20 Male:Female | 90:10 Male:Female | 100:0 Male:Female | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SNP 1 | PA 2 | SNP | PA | SNP | PA | SNP | PA | SNP | PA | |||||
| Before | After | Before | After | Before | After | Before | After | Before | After | Before | After | Before | After | |
| Total number of DArT analyses | 70,269 | - | 96,789 | - | 70,269 | - | 96,789 | - | - | - | - | - | - | - |
| Sex-linked loci | 142 | - | 7326 | - | 2 | - | 60 | - | - | - | - | - | - | - |
| Overall mean distance between males and females | 0.712 ± 0.020 | - | 0.582 ± 0.019 | - | 0.712 ± 0.031 | - | 0.673 ± 0.019 | - | - | - | - | - | - | - |
| Overall mean distance within females | 0.737 ± 0.023 | - | 0.665 ± 0.021 | - | 0.682 ± 0.052 | - | 0.033 ± 0.033 | - | - | - | - | - | - | - |
| Overall mean distance within males | 0.609 ± 0.036 | - | 0.507 ± 0.036 | - | 0.500 ± 0.062 | - | 0.462 ± 0.038 | - | - | - | - | - | - | - |
| 40:60 Male:Female | 30:70 Male:Female | 20:80 Male:Female | 10:90 Male:Female | 0:100 Male:Female | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SNP 1 | PA 2 | SNP | PA | SNP | PA | SNP | PA | SNP | PA | |||||
| Before | After | Before | After | Before | After | Before | After | Before | After | Before | After | Before | After | |
| Total number of DArT analyses | 70,269 | - | 96,789 | - | 70,269 | - | 96,789 | - | - | - | - | - | - | - |
| Sex-linked loci | 33] | - | 883 | - | - | - | 24 | - | - | - | - | - | - | - |
| Overall mean distance between males and females | 0.712 ± 0.020 | - | 0.582 ± 0.019 | - | - | - | 0.582 ± 0.019 | - | - | - | - | - | - | - |
| Overall mean distance within females | 0.737 ± 0.023 | - | 0.665 ± 0.021 | - | - | - | 0.665 ± 0.021 | - | - | - | - | - | - | - |
| Overall mean distance within males | 0.609 ± 0.036 | - | 0.507 ± 0.036 | - | - | - | 0.507 ± 0.036 | - | - | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panthum, T.; Singchat, W.; Laopichienpong, N.; Ahmad, S.F.; Kraichak, E.; Duengkae, P.; Muangmai, N.; Kitana, N.; Srikulnath, K. Genome-Wide SNP Analysis of Male and Female Rice Field Frogs, Hoplobatrachus rugulosus, Supports a Non-Genetic Sex Determination System. Diversity 2021, 13, 501. https://doi.org/10.3390/d13100501
Panthum T, Singchat W, Laopichienpong N, Ahmad SF, Kraichak E, Duengkae P, Muangmai N, Kitana N, Srikulnath K. Genome-Wide SNP Analysis of Male and Female Rice Field Frogs, Hoplobatrachus rugulosus, Supports a Non-Genetic Sex Determination System. Diversity. 2021; 13(10):501. https://doi.org/10.3390/d13100501
Chicago/Turabian StylePanthum, Thitipong, Worapong Singchat, Nararat Laopichienpong, Syed Farhan Ahmad, Ekaphan Kraichak, Prateep Duengkae, Narongrit Muangmai, Noppadon Kitana, and Kornsorn Srikulnath. 2021. "Genome-Wide SNP Analysis of Male and Female Rice Field Frogs, Hoplobatrachus rugulosus, Supports a Non-Genetic Sex Determination System" Diversity 13, no. 10: 501. https://doi.org/10.3390/d13100501
APA StylePanthum, T., Singchat, W., Laopichienpong, N., Ahmad, S. F., Kraichak, E., Duengkae, P., Muangmai, N., Kitana, N., & Srikulnath, K. (2021). Genome-Wide SNP Analysis of Male and Female Rice Field Frogs, Hoplobatrachus rugulosus, Supports a Non-Genetic Sex Determination System. Diversity, 13(10), 501. https://doi.org/10.3390/d13100501