
Potential Bioclimatic Ranges of Crop Pests Zabrus tenebrioides and Harpalus rufipes during Climate Change Conditions



Abstract
:1. Introduction
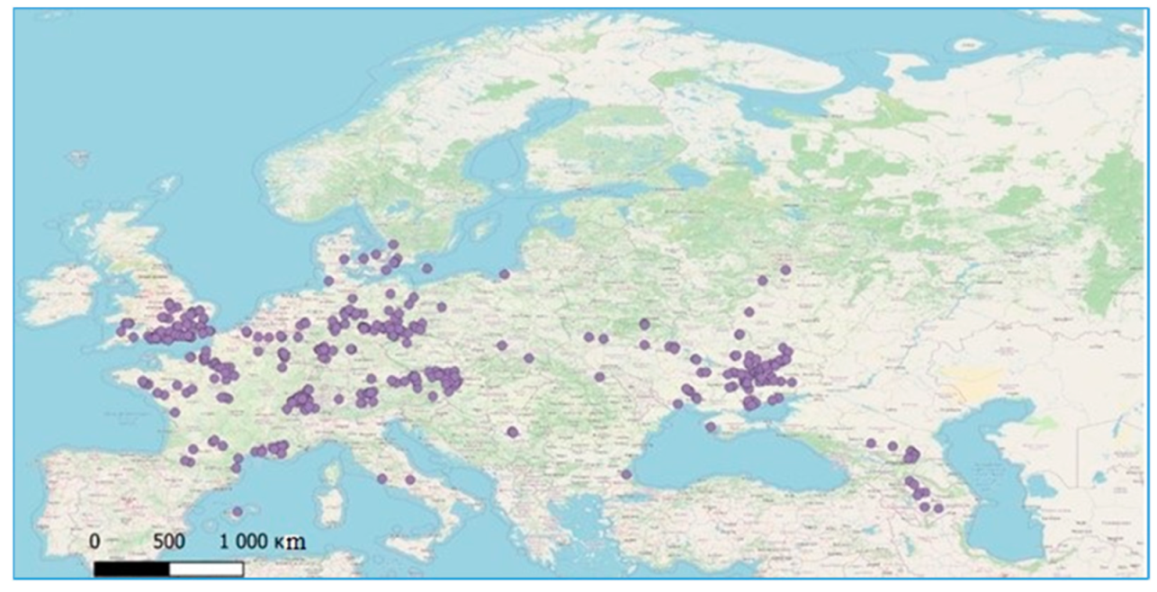
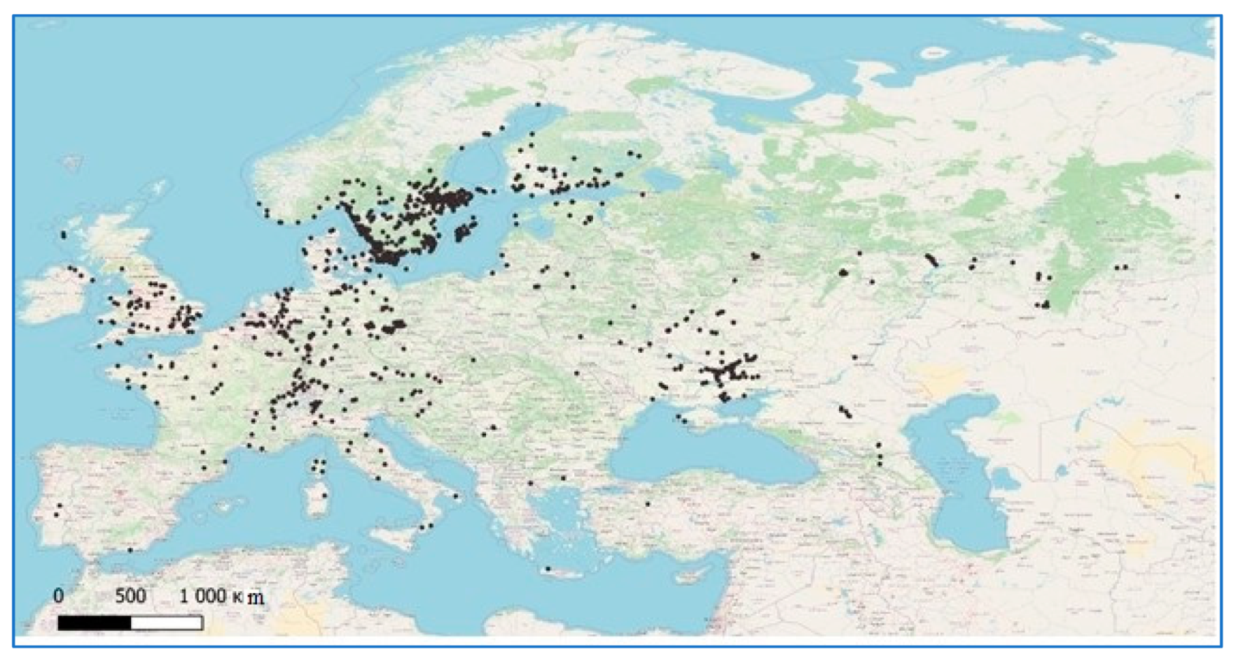
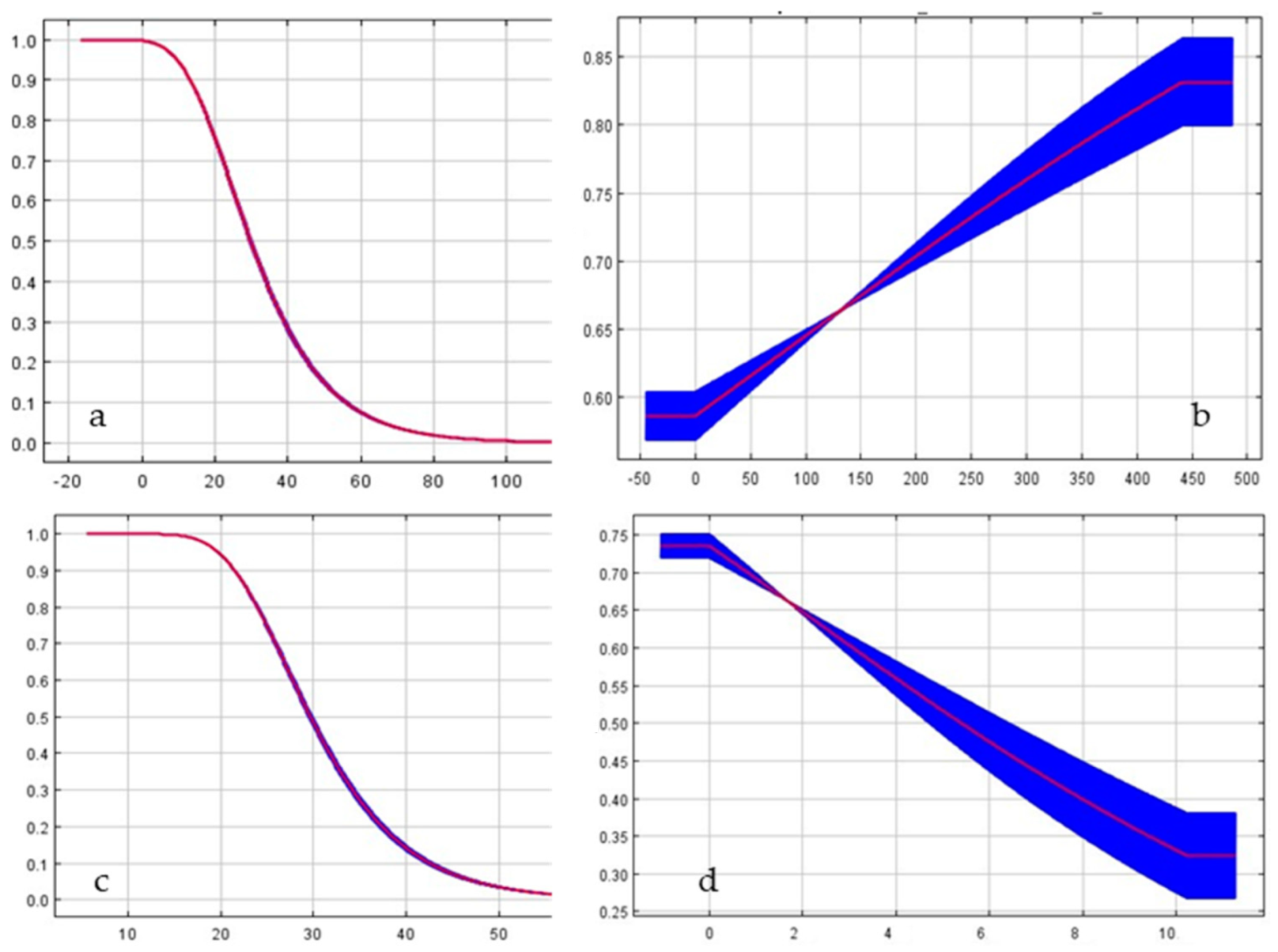
2. Materials and Methods
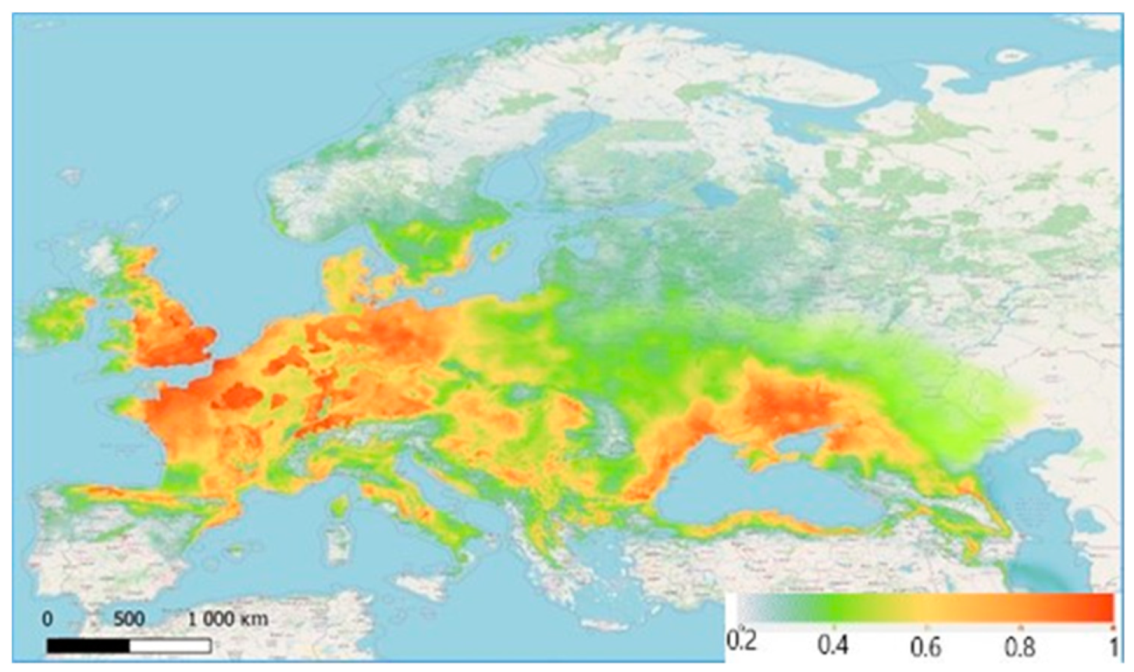
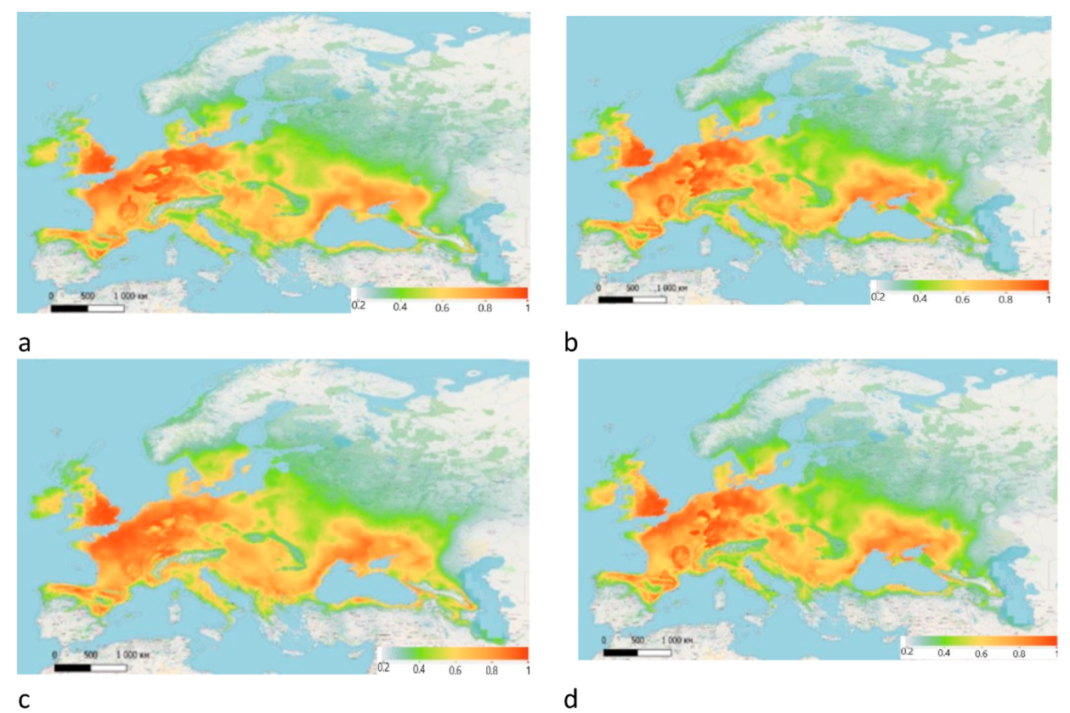
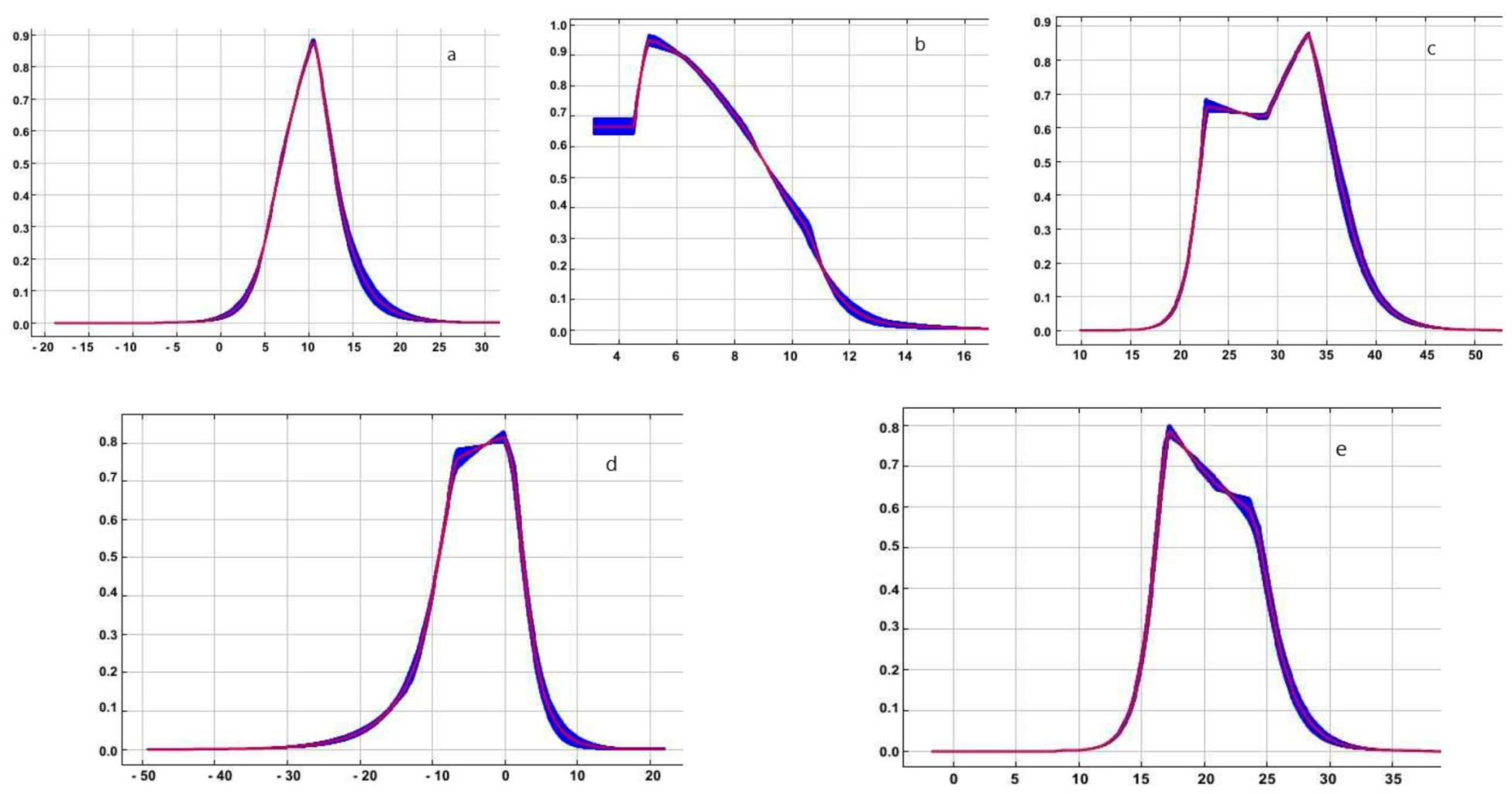
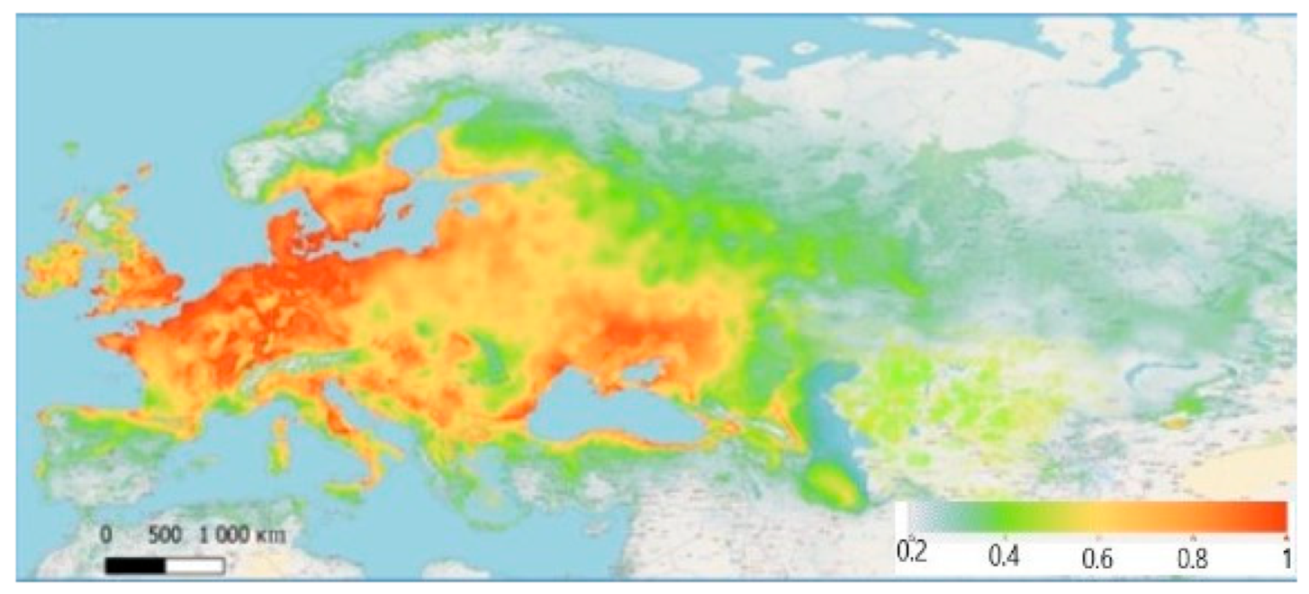
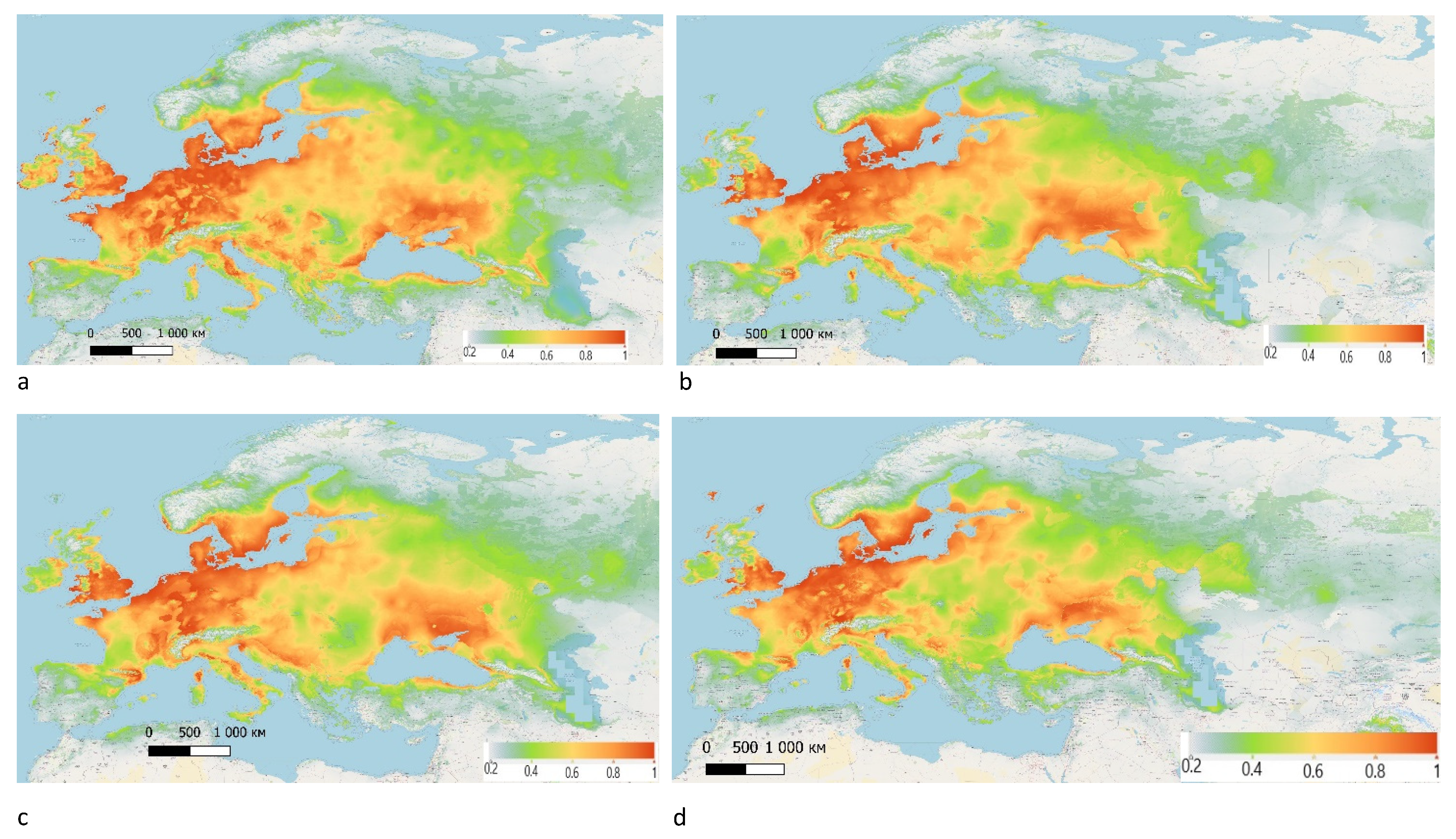
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Irmler, U.; Heller, K.; Meyer, H.; Reinke, H.-D. Zonation of ground beetles (Coleoptera: Carabidae) and spiders (Araneida) in salt marshes at the North and the Baltic Sea and the impact of the predicted sea level increase. Biodivers. Conserv. 2002, 11, 1129–1147. [Google Scholar] [CrossRef]
- Barton, P.S.; Gibbk, H.; Manningk, A.; Lindenmayer, D.B.; Cunningham, S.A. Morphological traits as predictors of diet and microhabitat use in a diverse beetle assemblage. Biol. J. Linn. Soc. 2011, 102, 301–310. [Google Scholar] [CrossRef]
- Langraf, V.; Petrovičová, K.; Krumpálová, Z.; Svoradová, A.; Schlarmannová, J. Dispersion of the epigeic fauna groups in the agricultural landscape. Folia Oecol. 2021, 48, 147–155. [Google Scholar] [CrossRef]
- Loughridge, A.H.; Luff, M.L. Aphid predation by Harpalus rufipes (De Geer) (Coleoptera: Carabidae) in the laboratory and field. J. Appl. Ecol. 1983, 20, 451–462. [Google Scholar] [CrossRef]
- Ishitani, M.; Watanabe, J.; Yano, K. Species composition and spatial distribution of ground beetles (Coleoptera) in a forage crop field. Jpn. J. Entomol. 1994, 62, 275–283. [Google Scholar]
- Holland, J.M.; Luff, M.L. The effects of agricultural practices on Carabidae in temperate ecosystems. Integr. Pest. Manag. Rev. 2000, 5, 109–129. [Google Scholar] [CrossRef]
- Honek, A.; Martinkova, Z.; Jarosik, V. Ground beetles (Carabidae) as seed predators. Eur. J. Entomol. 2003, 100, 531–544. [Google Scholar] [CrossRef] [Green Version]
- Honek, A.; Martinkova, Z.; Saska, P.; Pekar, S. Size and taxonomic constraints determine the seed preferences of Carabidae (Coleoptera). Basic Appl. Ecol. 2007, 8, 343–353. [Google Scholar] [CrossRef]
- Monzó, C.; Sabater-Muñoz, B.; Urbaneja, A.; Castañera, P. The ground beetle Pseudophonus rufipes revealed as predator of Ceratitis capitata in citrus orchards. Biol. Control 2011, 56, 17–21. [Google Scholar] [CrossRef]
- Sasakawa, K. Field observations of climbing behavior and seed predation by adult ground beetles (Coleoptera: Carabidae) in a lowland area of the temperate zone. Environ. Entomol. 2010, 39, 1554–1560. [Google Scholar] [CrossRef]
- Jørgensen, H.B.; Toft, S. Food preference, diet dependent fecundity and larval development in Harpalus rufipes (Coleoptera: Carabidae). Pedobiologia 1997, 41, 307–315. [Google Scholar]
- Di Grumo, D.; Lovei, G. Body size inequality in ground beetle (Coleoptera: Carabidae) assemblages as a potential method to monitor environmental impacts of transgenic crops. Period. Biol. 2016, 118, 223–230. [Google Scholar] [CrossRef]
- Beljaev, V.I.; Volnov, V.V.; Sokolova, L.V.; Kuznecov, V.N.; Matsyura, A.V. Effect of sowing techniques on the agroecological parameters of cereal crops. Ukr. J. Ecol. 2017, 7, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Bezpalko, V.V.; Stankevych, S.V.; Zhukova, L.V.; Zabrodina, I.V.; Turenko, V.P.; Horyainova, V.V.; Poedinceva, A.A.; Batova, O.M.; Zayarna, O.Y.; Bondarenko, S.V.; et al. Pre-sowing seed treatment in winter wheat and spring barley cultivation. Ukr. J. Ecol. 2020, 10, 255–268. [Google Scholar] [CrossRef]
- Coombes, D.S.; Sotherton, N.W. The dispersal and distribution of polyphagous predatory Coleoptera in cereals. Ann. Appl. Biol. 1986, 108, 461–474. [Google Scholar] [CrossRef]
- Goldschmidt, H.; Toft, S. Variable degrees of granivory and phytophagy in insectivorous carabid beetles. Pedobiologia 1997, 41, 521–525. [Google Scholar]
- Jacob, H.S.; Minkey, D.M.; Gallagher, R.S.; Borger, C.P. Variation in postdispersal weed seed predation in a crop field. Weed Sci. 2006, 54, 148–155. [Google Scholar] [CrossRef]
- Harrison, S.; Gallandt, E.R. Behavioural studies of Harpalus rufipes De Geer: An important weed seed predator in northeastern US agroecosystems. Int. J. Ecol. 2012, 2012, 1–6. [Google Scholar] [CrossRef]
- Hengeveld, R. Polyphagy, oligophagy and food specialization in ground beetles (Coleoptera: Carabidae). Neth. J. Zool. 1980, 30, 564–584. [Google Scholar] [CrossRef]
- Putchkov, A.V.; Brygadyrenko, V.V.; Nikolenko, N.Y. Ecological-faunistic analysis of ground beetles and tiger beetles (Coleoptera: Carabidae, Cicindelidae) of metropolises of Ukraine. Biosyst. Divers. 2020, 28, 163–174. [Google Scholar] [CrossRef]
- Avtaeva, T.A.; Sukhodolskaya, R.A.; Skripchinsky, A.V.; Brygadyrenko, V.V. Range of Pterostichus oblongopunctatus (Coleoptera, Carabidae) in conditions of global climate change. Biosyst. Divers. 2019, 27, 76–84. [Google Scholar] [CrossRef]
- Avtaeva, T.; Skripchinsky, A.; Brygadyrenko, V. Changes in the range of Pterostichus melas and P. fornicatus (Coleoptera, Carabidae) on the basis of climatic modeling. Baltic J. Coleopterol. 2020, 20, 109–124. [Google Scholar]
- Avtaeva, T.A.; Sukhodolskaya, R.A.; Brygadyrenko, V.V. Modeling the bioclimating range of Pterostichus melanarius (Coleoptera, Carabidae) in conditions of global climate change. Biosyst. Divers. 2021, 29, 140–150. [Google Scholar] [CrossRef]
- Brandmayr, P.; Pizzolotto, R. Climate change and its impact on epigean and hypogean and hypogean carabid beetles. Period. Biol. 2016, 118, 147–162. [Google Scholar] [CrossRef]
- Oosterbroek, P.; Arntzen, J.W. Area-cladograms of Circum-Mediterranean taxa in relation to Mediterranean paleogeography. J. Biogeogr. 1992, 19, 3–20. [Google Scholar] [CrossRef]
- Pizzolotto, R.; Brandmayr, P.; Mazzei, A. Carabid beetles in a Mediterranean Region: Biogeographical and ecological features. DIAS Rep. 2005, 114, 243–254. [Google Scholar]
- Andersen, J. A re-analysis of the relationship between life cycle patterns and the geographical distribution of Fennoscandian carabid beetles. J. Biogeogr. 1984, 11, 479–489. [Google Scholar] [CrossRef]
- Brygadyrenko, V.V.; Nazimov, S.S. Trophic relations of Opatrum sabulosum (Coleoptera, Tenebrionidae) with leaves of cultivated and uncultivated species of herbaceous plants under laboratory conditions. Zookeys 2015, 481, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Shulman, M.V.; Pakhomov, O.Y.; Brygadyrenko, V.V. Effect of lead and cadmium ions upon the pupariation and morphological changes in Calliphora vicina (Diptera, Calliphoridae). Folia Oecol. 2017, 44, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Makaida, M.V.; Pakhomov, O.Y.; Brygadyrenko, V.V. Effect of increased ambient temperature on seasonal generation number in Lucilia sericata (Diptera, Calliphoridae). Folia Oecol. 2021, 48, 191–198. [Google Scholar] [CrossRef]
- Ganglbauer, L. Revision der Gattung Zabrus Clairv. Münch. Koleopt. Z. 1915, 4, 87–137. [Google Scholar]
- Lindroth, C.H. Coleoptera, Carabidae. Handbooks for the Identification of British Insects; Royal Entomological Society of London: London, UK, 1974. [Google Scholar]
- Lindroth, C.H. The Carabidae (Coleoptera) of Fennoscandia and Denmark. Fauna Entomol. Scand. 1985, 15, 1–226. [Google Scholar]
- Kryzhanovskij, O.L.; Belousov, I.A.; Kabak, I.I.; Kataev, B.M.; Makarov, K.V.; Shilenkov, V.G. A Checklist of the Ground-Beetles of Russia and Adjacent Lands; Pensoft: Sofia-Moscow, Russia, 1995. [Google Scholar]
- Hurka, K. Carabidae of the Czech. and Slovak Republics; Kabourek: Zlín, Czechia, 1996. [Google Scholar]
- Talarico, F.; Giglio, A.; Pizzolotto, R.; Brandmayr, P. A synthesis of feeding habits and reproduction rhythm in Italian seed-feeding ground beetles (Coleoptera: Carabidae). Eur. J. Entomol. 2016, 113, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Sienkiewicz, P.; Lipa, J.J. Gregarina vizri Lipa, 1968 (Apicomplexa, Eugregarinida) recorded in Poland in an expansive plant pest the cereal ground beetle Zabrus tenebrioides (Goeze) (Coleoptera, Carabidae). J. Plant. Prot. Res. 2008, 48, 189–193. [Google Scholar] [CrossRef]
- Kulkarni, S.S.; Dosdall, L.M.; Willenborg, C.J. The role of ground beetles (Coleoptera: Carabidae) in weed seed consumption: A review. Weed Sci. 2015, 63, 335–376. [Google Scholar] [CrossRef]
- Freude, H. Revision der zur Gattung “Zabrus” Clairville, 1806 gehörenden Arten mit Bestimmungstabellen (Col. Carabidae). Atti Mus. Civ. Stor. Nat. Trieste 1986, 39, 133–150. [Google Scholar]
- Freude, H. Revision der zur Gattung “Zabrus” Clairville, 1806 gehörenden Arten mit Bestimmungstabellen (Col. Carabidae). 2. Teil. Atti Mus. Civ. Stor. Nat. Trieste 1988, 41, 133–142. [Google Scholar]
- Freude, H. Revision der zur Gattung “Zabrus” Clairville, 1806 gehörenden Arten mit Bestimmungstabellen (Col. Carabidae). 3. Teil. Atti Mus. Civ. Stor. Nat. Trieste 1989, 42, 71–153. [Google Scholar]
- Freude, H. Revision der zur Gattung “Zabrus” Clairville 1806 gehörenden Arten mit Bestimmungstabellen (Col. Carabidae). 4. Atti Mus. Civ. Stor. Nat. Trieste 1990, 43, 329–364. [Google Scholar]
- Andújar, A.; Serrano, J. Taxonomic notes on the genus Zabrus (Coleoptera, Carabidae, Zabrini). Boln. Asoc. Esp. Entomol. 2000, 24, 185–193. [Google Scholar]
- Sánchez-Gea, J.F.; Serrano, J.; Galián, J. Variability in rDNA loci in Iberian species of the genus Zabrus (Coleoptera: Carabidae) detected by fluorescence in situ hybridization. Genome 2000, 43, 22–28. [Google Scholar] [CrossRef]
- Sánchez-Gea, J.-F.; Galián, J.; Serrano, J. Phylogeny of Iberian Zabrus (Coleoptera: Carabidae: Zabrini) based on mitochondrial DNA sequence. Eur. J. Entomol. 2004, 101, 503–511. [Google Scholar] [CrossRef] [Green Version]
- Reshetniak, D.Y.; Pakhomov, O.Y.; Brygadyrenko, V.V. Possibility of identifying plant components of the diet of Harpalus rufipes (Coleoptera, Carabidae) by visual evaluation. Regul. Mech. Biosyst. 2017, 8, 377–383. [Google Scholar] [CrossRef]
- El-Danasoury, H.; Cerecedo, C.; Córdoba, M.; Iglesias-Piñeiro, J. Predation by the carabid beetle Harpalus rufipes on the pest slug Deroceras reticulatum in the laboratory. Ann. Appl. Biol. 2017, 170, 251–262. [Google Scholar] [CrossRef]
- Forsythe, T.G. Feeding mechanisms of certain ground beetles (Coleoptera: Carabidae). Coleopt. Bull. 1982, 36, 26–73. [Google Scholar]
- Gaines, H.R.; Gratton, C. Seed predation increases with ground beetle diversity in a Wisconsin (USA) potato agroecosystem. Agric. Ecosyst. Environ. 2010, 137, 329–336. [Google Scholar] [CrossRef]
- Birthisel, S.K.; Gallandt, E.R.; Jabbour, R. Habitat effects on second-order predation of the seed predator Harpalus rufipes and implications for weed seedbank management. Biol. Control 2014, 70, 65–72. [Google Scholar] [CrossRef]
- Leibman, M. Effect of crop habitat and potato management practices on the population abundance of adult Harpalus rufipes (Coleoptera: Carabidae) in Maine. J. Agric. Entomol. 1998, 15, 63–74. [Google Scholar]
- Brygadyrenko, V.V.; Reshetniak, D.Y. Morphological variability among populations of Harpalus rufipes (Coleoptera, Carabidae): What is more important—The mean values or statistical peculiarities of distribution in the population? Folia Oecol. 2014, 41, 109–133. [Google Scholar]
- Cavaliere, F.; Brandmayr, P.; Giglio, A. DNA damage in haemocytes of Harpalus (Pseudophonus) rufipes (De Geer, 1774) (Coleoptera, Carabidae) as an indicator of sublethal effects of exposure to herbicides. Ecol. Indic. 2019, 98, 88–91. [Google Scholar] [CrossRef]
- Langraf, V.; David, S.; Babosová, R.; Petrovičová, K.; Schlarmannová, J. Change of ellipsoid biovolume (EV) of ground beetles (Coleoptera, Carabidae) along an urban–suburban–rural gradient of Central Slovakia. Diversity 2020, 12, 475. [Google Scholar] [CrossRef]
- Thomas, C.F.G.; Green, F.; Marshall, E.J.P. Distribution, dispersal and population size of the ground beetles, Pterostichus melanarius (Illiger) and Harpalus rufipes (Degeer) (Coleoptera, Carabidae), in field margin habitats. Biol. Agric. Hortic. 1997, 15, 337–352. [Google Scholar] [CrossRef]
- Kujawa, K.; Sobczyk, D.; Kajak, A. Dispersal of Harpalus rufipes (Degeer) (Carabidae) between shelterbelt and cereal field. Polish J. Ecol. 2006, 54, 243–252. [Google Scholar]
- Brygadyrenko, V.V.; Reshetniak, D.Y. Trophic preferences of Harpalus rufipes (Coleoptera, Carabidae) with regard to seeds of agricultural crops in conditions of laboratory experiment. Baltic J. Coleopterol. 2014, 14, 179–190. [Google Scholar]
- Brygadyrenko, V.V.; Reshetniak, D.Y. Morphometric variability of Clitellocephalus ophoni (Eugregarinida, Gregarinidae) in the intestines of Harpalus rufipes (Coleoptera, Carabidae). Arch. Biol. Sci. 2016, 68, 587–601. [Google Scholar] [CrossRef]
- Faly, L.I.; Kolombar, T.M.; Prokopenko, E.V.; Pakhomov, O.Y.; Brygadyrenko, V.V. Structure of litter macrofauna communities in poplar plantations in an urban ecosystem in Ukraine. Biosyst. Divers. 2017, 25, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Wallin, H. Spatial and temporal distribution of some abundant carabid beetles (Coleoptra, Carabidae) in cereal fields in adjacent habitats. Pedobiologia 1985, 28, 19–34. [Google Scholar]
- Tamutis, V.; Tamutel, B.; Ferenca, R. A catalogue of Lithuanian beetles (Insecta, Coleoptera). Zookeys 2011, 121, 1–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langraf, V.; Petrovičová, K.; David, S.; Kanská, M.; Nozdrovická, J.; Schlarmannová, J. Change phenotypic traits in ground beetles (Carabidae) reflects biotope disturbance in Central Europe. J. Entomol. Res. Soc. 2018, 20, 119–129. [Google Scholar]
- Langraf, V.; Petrovičová, K.; David, S.; Svoradová, A.; Schlarmannová, J. Prediction of ecological importance of Carabidae biotopes using community index of the ground beetles (iks) in the southern part of Central Slovakia. Appl. Ecol. Environ. Res. 2020, 18, 1197–1210. [Google Scholar] [CrossRef]
- Johnson, N.E.; Cameron, R.S. Phytophagous ground beetles. Ann. Entomol. Soc. Am. 1969, 62, 909–914. [Google Scholar] [CrossRef]
- Zhang, J.; Drummond, F.A.; Liebman, M.; Hartke, A. Phenology and dispersal of Harpalus rufipes DeGeer (Coleoptera: Carabidae) in agroecosystems in Maine. J. Agric. Urban. Entomol. 1997, 14, 171–186. [Google Scholar]
- Kunah, O.M.; Zelenko, Y.V.; Fedushko, M.P.; Babchenko, A.V.; Sirovatko, V.O.; Zhukov, O.V. The temporal dynamics of readily available soil moisture for plants in the technosols of the Nikopol Manganese Ore Basin. Biosyst. Divers. 2019, 27, 156–162. [Google Scholar]
- Eyre, M.D.; Luff, M.L. Ground beetle species (Coleoptera, Carabidae) associations with land cover variables in Northern England and Southern Scotland. Ecography 2004, 27, 417–426. [Google Scholar] [CrossRef]
- Jensen, L.B. Effect of temperature on the development of the immature stages of Bembidion lampros (Coleoptera: Carabidae). Entomophaga 1990, 35, 277–281. [Google Scholar] [CrossRef]
- Knapp, M.; Saska, P. The effects of habitat, density, gender and duration on overwintering success in Bembidion lampros (Coleoptera: Carabidae). J. Appl. Entomol. 2012, 136, 225–233. [Google Scholar] [CrossRef]
- Brygadyrenko, V.; Avtaeva, T.; Matsyura, A. Effect of global climate change on the distribution of Anchomenus dorsalis (Coleoptera, Carabidae) in Europe. Acta Biol. Sibirica 2021, 7, 237–259. [Google Scholar] [CrossRef]
- Gobbi, M.; Fontaneto, D. Biodiversity of ground beetles (Coleoptera: Carabidae) in different habitats of the Italian Po lowland. Agric. Ecosyst. Environ. 2008, 127, 273–276. [Google Scholar] [CrossRef]
- Brygadyrenko, V.V. Parameters of ecological niches of Badister, Licinus and Panagaeus (Coleoptera, Carabidae) species measured against eight ecological factors. Balt. J. Coleopterol. 2015, 15, 137–154. [Google Scholar]
- Kaizuka, J.; Iwasa, M. Carabid beetles (Coleoptera: Carabidae) in coniferous plantations in Hokkaido, Japan: Effects of tree species and environmental factors. Entomol. Sci. 2015, 18, 245–253. [Google Scholar] [CrossRef]
- Langraf, V.; Petrovičová, K.; David, S.; Nozdrovická, J.; Petrovič, F.; Schlarmannová, J. The bioindication evaluation of ground beetles (Coleoptera: Carabidae) in three forest biotopes in the southern part of Central Slovakia. Ekológia (Bratislava) 2019, 38, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Ernsting, G.; Isaaks, J.A. Effects of temperature and season on egg size, hatchling size and adult size in Notiophilus biguttatus. Ecol. Entomol. 1997, 22, 32–40. [Google Scholar] [CrossRef]
- Desender, K.; Maelfait, J.-P. Diversity and conservation of terrestrial arthropods in tidal marshes along the river Schelde: A gradient analysis. Biol. Conserv. 1999, 87, 221–229. [Google Scholar] [CrossRef]
- Desender, K.; Verdyck, P. Geographic scaling and genetic differentiation in two highly mobile European saltmarsh beetles. Belg. J. Zool. 2001, 131, 31–42. [Google Scholar]
- Nietupski, M.; Sowinski, P.; Sadej, W.; Kosewska, A. Content of organic C and pH of bog and post-bog soils versus the presence of ground beetles Carabidae in Stary Dwor near Olsztyn. J. Elem. 2010, 15, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Brygadyrenko, V.V. Evaluation of the ecological niche of some abundant species of the subfamily Platyninae (Coleoptera, Carabidae) against the background of eight ecological factors. Folia Oecol. 2015, 42, 75–88. [Google Scholar]
- Schultz, R. Ground beetles (Coleoptera: Carabidae) as indicators for the efficiency of the revitalization of salt grassland at the Baltic Sea coast. Mitt. Dtsch. Ges. Allg. Angew. Entomol. 2000, 12, 389–394. [Google Scholar]
- Komlyk, V.O.; Brygadyrenko, V.V. Morphological variability of Bembidion aspericolle (Coleoptera, Carabidae) populations in conditions of anthropogenic impact. Biosyst. Divers. 2019, 27, 21–25. [Google Scholar] [CrossRef]
- Komlyk, V.O.; Brygadyrenko, V.V. Morphological variability of Bembidion minimum (Coleoptera, Carabidae) populations under the influence of natural and anthropogenic factors. Biosyst. Divers. 2019, 27, 250–269. [Google Scholar] [CrossRef] [Green Version]
- Rodda, G.H.; Jarnevich, C.S.; Reed, R.N. Challenges in identifying sites climatically matched to the native ranges of animal invaders. PLoS ONE 2011, 6, e14670. [Google Scholar] [CrossRef] [Green Version]
- Maharaj, S.S.; New, M. Modelling individual and collective species responses to climate change within small island states. Biol. Conserv. 2013, 167, 283–291. [Google Scholar] [CrossRef]
- Fitzpatrick, M.C.; Gotelli, N.J.; Ellison, A.M. MaxEnt versus MaxLike: Empirical comparisons with ant species distributions. Ecosphere 2013, 4, 55. [Google Scholar] [CrossRef]
- Norris, D. Model thresholds are more important than presence location type: Understanding the distribution of lowland tapir (Tapirus terrestris) in a continuous Atlantic forest of Southeast Brazil. Trop. Conserv. Sci. 2014, 7, 529–547. [Google Scholar] [CrossRef] [Green Version]
- Namyatova, A.A. Climatic niche comparison between closely related trans-palearctic species of the genus Orthocephalus (Insecta: Heteroptera: Miridae: Orthotylinae). PeerJ 2020, 8, e10517. [Google Scholar] [CrossRef]
- Zhang, V.M.; Punzalan, D.; Rowe, L. Climate change has different predicted effects on the range shifts of two hybridizing ambush bug (Phymata, Family Reduviidae, Order Hemiptera) species. Ecol. Evol. 2020, 10, 12036–12048. [Google Scholar] [CrossRef]
- Fedorenko, V.P. Fitosanitarnoe sostoyanie agrocenozov Ukrainy. Zashchita Karantin Rastenij 2012, 4, 48–52. (In Russian) [Google Scholar]
- Beleckij, E.N.; Stankevich, S.V. Policiklichnost, Sinhronnost i Nelinejnost Populyacionnoj Dinamiki Nasekomykh i Problemy Prognozirovaniya; Premier Publishing: Vienna, Austria, 2018. [Google Scholar]
- Zhavoronkova, T.N. Certain structural peculiarities of the Carabidae (Coleoptera) in relation to their feeding habits. Entomol. Rev. 1969, 48, 462–471. [Google Scholar]
- Trepashko, L.I.; Bojko, S.V. Invaziya obyknovennoj hlebnoj zhuzhelicy na territoriju Belarusi. Zashchita i Karantin Rastenij 2016, 11, 32–35. (In Russian) [Google Scholar]
- Beleckij, E.N. Massovye Razmnozhenija Nasekomyh: Istoriya, Teoriya, Prognozirovanie; Majdan: Kharkov, Ukraine, 2011. (In Russian) [Google Scholar]
- Beleckij, E.N. Fitosanitarnoe prognozirovanie na Ukraine: Istorija, metodologija, puti sovershenstvovanija. Zashchita Karantin Rastenij 2015, 12, 14–19. (In Russian) [Google Scholar]
- Hilevskij, V.A. Khlebnaja zhuzhelica na Severnom Kavkaze. Zashchita Karantin Rastenij 2013, 8, 21–23. (In Russian) [Google Scholar]
- Skrebcova, T.I.; Chenikalova, E.V.; Mukhina, O.V. Khlebnaja zhuzhelica na ozimykh Stavropolija. Zashchita Karantin Rastenij 2011, 8, 15–16. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioclimatic Parameters | Bio 17 | Bio 15 | Bio 7 | Bio 19 | Bio 2 | Bio 9 | Bio 18 | Bio 8 |
|---|---|---|---|---|---|---|---|---|
| AUC | 0.89 | 0.87 | 0.86 | 0.85 | 0.78 | 0.63 | 0.62 | 0.25 |
| Climatic Scenario | Coefficient of Presence of Species | |||||
|---|---|---|---|---|---|---|
| 0.90–1.00 | 0.70–0.89 | 0.60–0.69 | 0.40–0.59 | 0.30–0.39 | Total | |
| Potential range, 2021 | 478,230 | 551,626 | 725,497 | 1,105,574 | 1,774,793 | 4,635,720 |
| RCP2.6, 2050 | 493,059 | 528,935 | 800,890 | 1,273,991 | 1,873,638 | 4,970,512 |
| RCP2.6, 2070 | 210,635 | 488,371 | 806,108 | 919,945 | 1,015,639 | 3,440,697 |
| RCP8.5, 2050 | 247,320 | 463,139 | 807,477 | 1,081,459 | 1,015,723 | 3,615,117 |
| RCP8.5, 2070 | 206,632 | 619,587 | 945,061 | 1,041,712 | 1,124,416 | 3,937,408 |
| Bioclimatic Parameters | Bio 1 | Bio 6 | Bio 2 | Bio 10 | Bio 3 |
|---|---|---|---|---|---|
| AUC | 0.86 | 0.83 | 0.78 | 0.77 | 0.73 |
| Climatic Scenario | Coefficient of Presence of Species | |||||
|---|---|---|---|---|---|---|
| 0.90–1.00 | 0.70–0.89 | 0.60–0.69 | 0.40–0.59 | 0.30–0.39 | Total | |
| Potential range, 2021 | 45,983 | 543,186 | 689,134 | 1,371,795 | 1,557,676 | 4,207,775 |
| RCP2.6, 2050 | 74,731 | 455,780 | 405,665 | 827,129 | 1,665,526 | 3,428,831 |
| RCP2.6, 2070 | 76,567 | 346,328 | 577,383 | 979,503 | 1,727,848 | 3,707,630 |
| RCP8.5, 2050 | 68,762 | 346,852 | 554,552 | 1,011,087 | 1,737,403 | 3,718,656 |
| RCP8.5, 2070 | 47,303 | 293,142 | 574,259 | 731,451 | 1,422,442 | 3,068,597 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avtaeva, T.; Petrovičová, K.; Langraf, V.; Brygadyrenko, V. Potential Bioclimatic Ranges of Crop Pests Zabrus tenebrioides and Harpalus rufipes during Climate Change Conditions. Diversity 2021, 13, 559. https://doi.org/10.3390/d13110559
Avtaeva T, Petrovičová K, Langraf V, Brygadyrenko V. Potential Bioclimatic Ranges of Crop Pests Zabrus tenebrioides and Harpalus rufipes during Climate Change Conditions. Diversity. 2021; 13(11):559. https://doi.org/10.3390/d13110559
Chicago/Turabian StyleAvtaeva, Tamara, Kornélia Petrovičová, Vladimir Langraf, and Viktor Brygadyrenko. 2021. "Potential Bioclimatic Ranges of Crop Pests Zabrus tenebrioides and Harpalus rufipes during Climate Change Conditions" Diversity 13, no. 11: 559. https://doi.org/10.3390/d13110559
APA StyleAvtaeva, T., Petrovičová, K., Langraf, V., & Brygadyrenko, V. (2021). Potential Bioclimatic Ranges of Crop Pests Zabrus tenebrioides and Harpalus rufipes during Climate Change Conditions. Diversity, 13(11), 559. https://doi.org/10.3390/d13110559