
Diversity of Insect Flower Visitors of Xylopia aromatica (Magnoliales, Annonaceae) in a Brazilian Savanna




Abstract
:1. Introduction
2. Materials and Methods
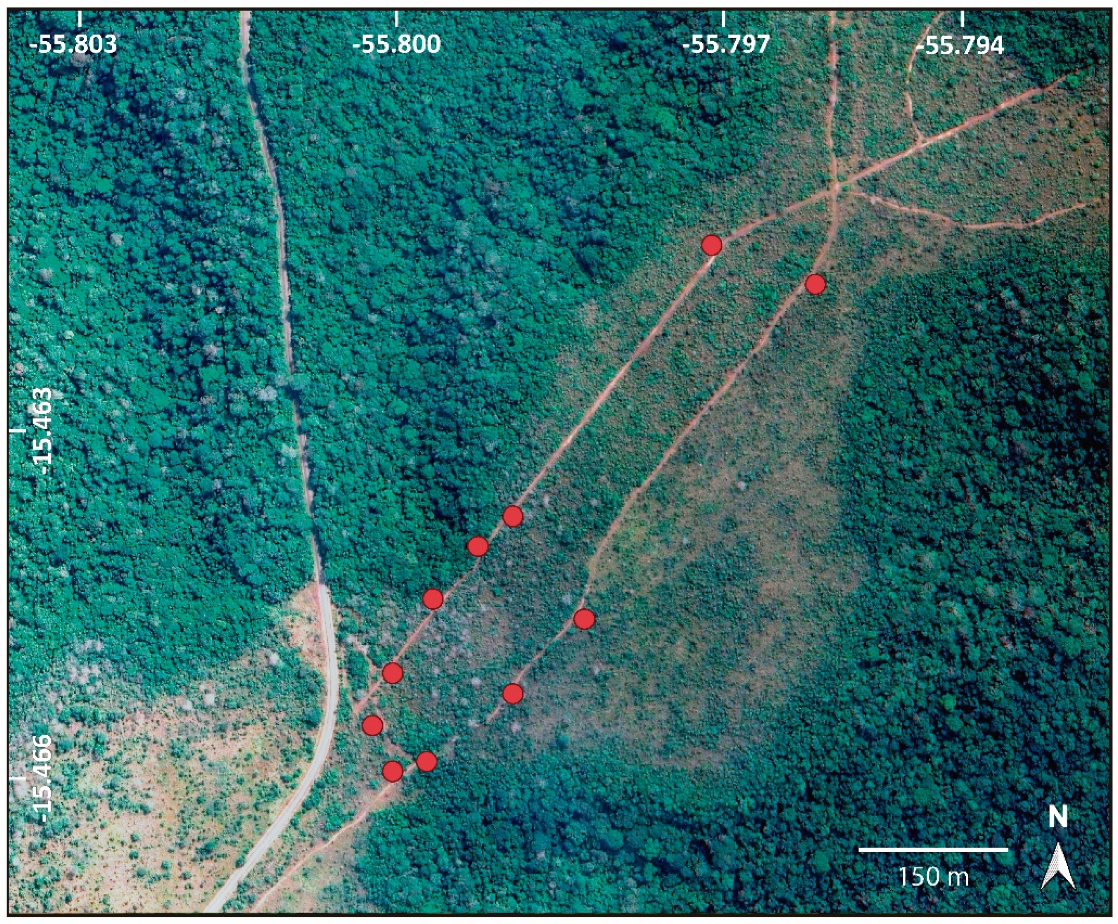
2.1. Study Area
2.2. Sampling of Floral Visitors
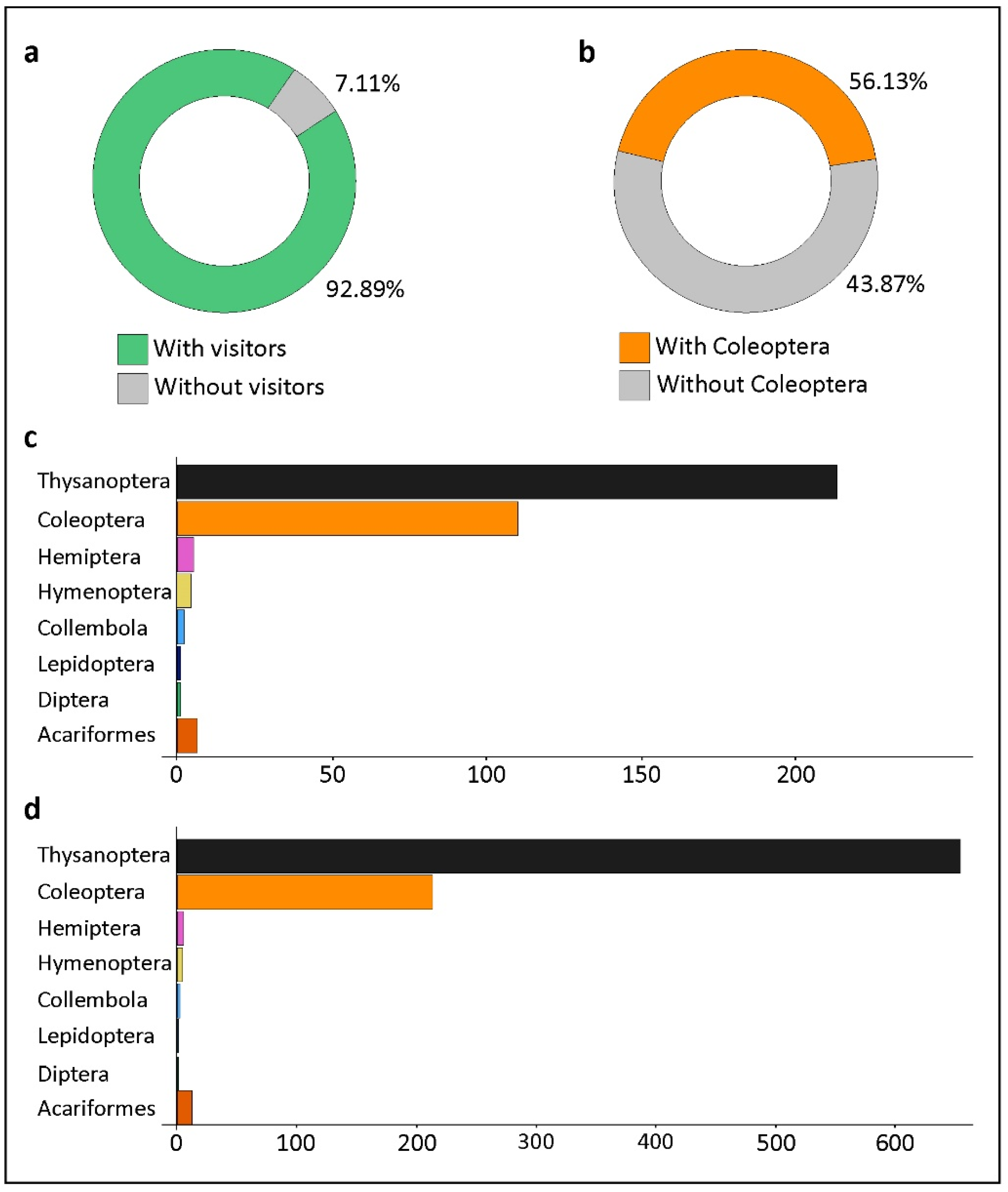
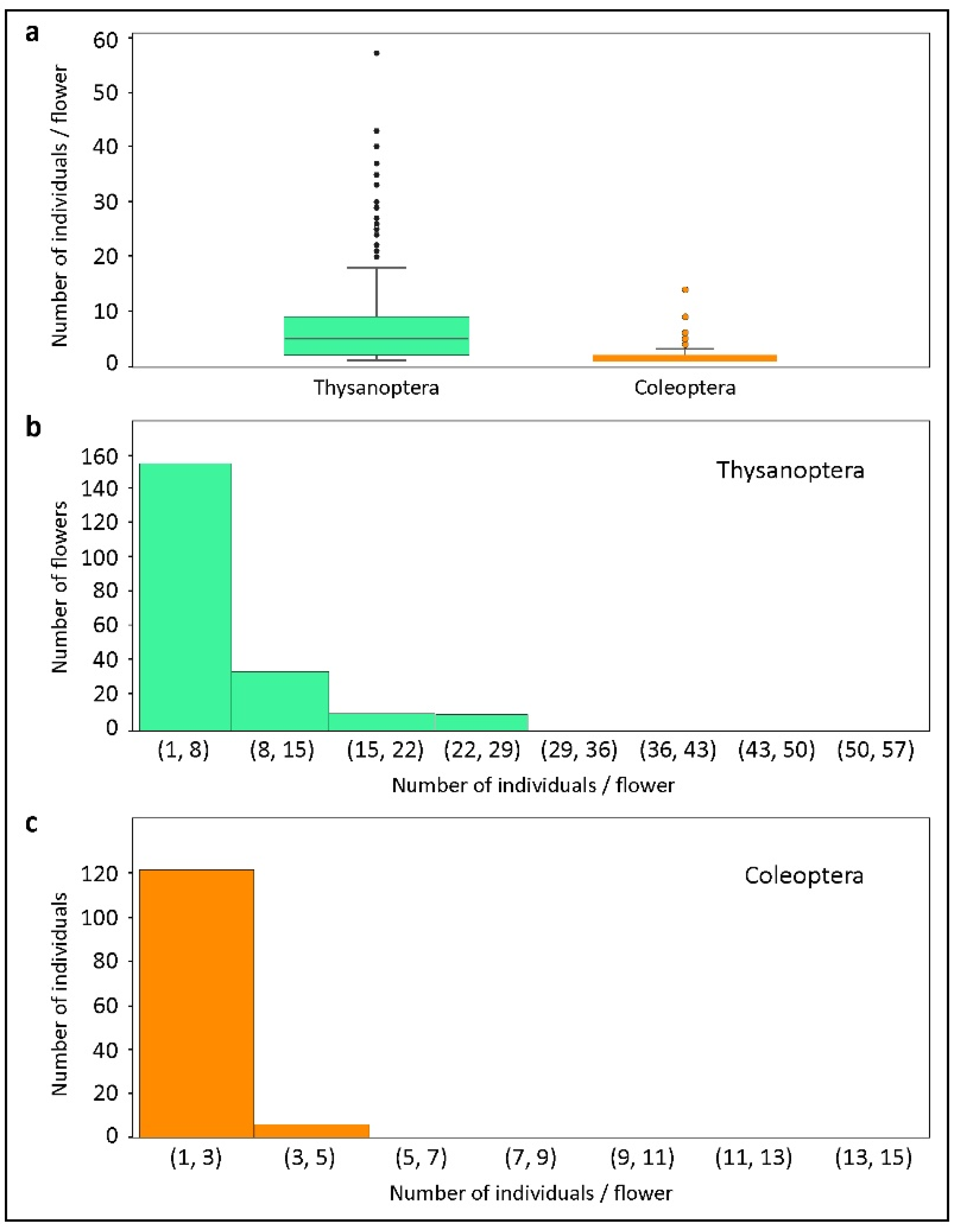
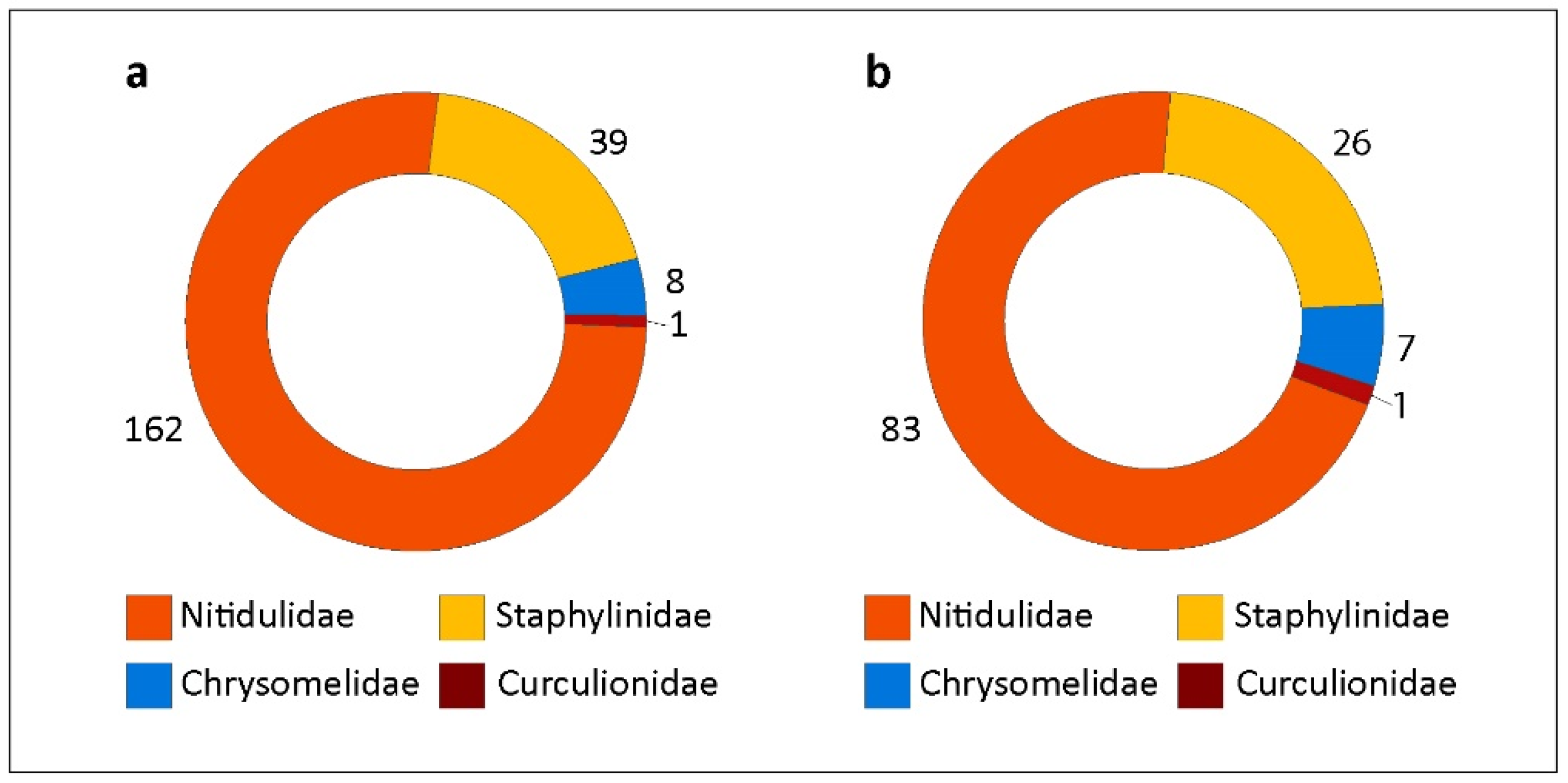
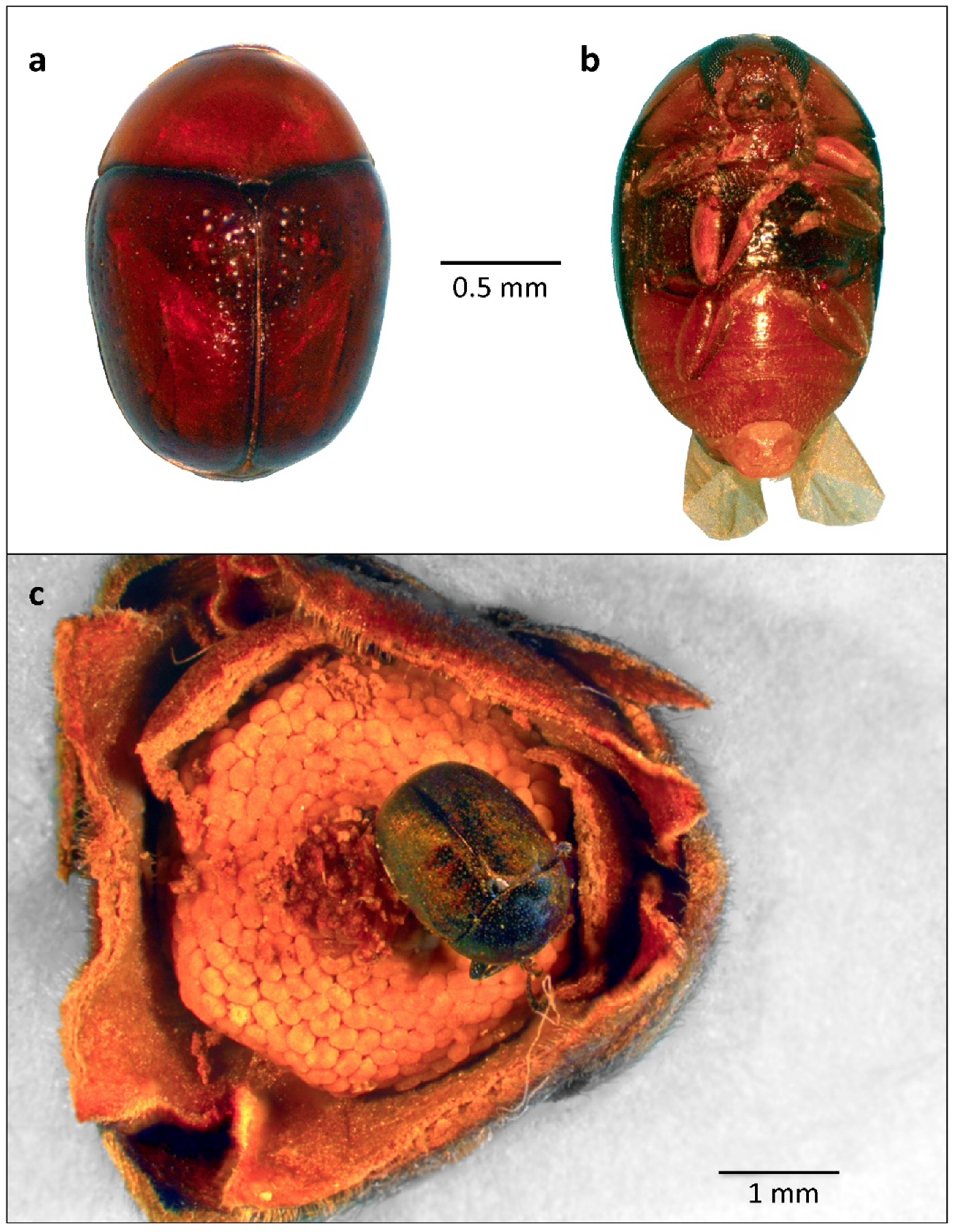
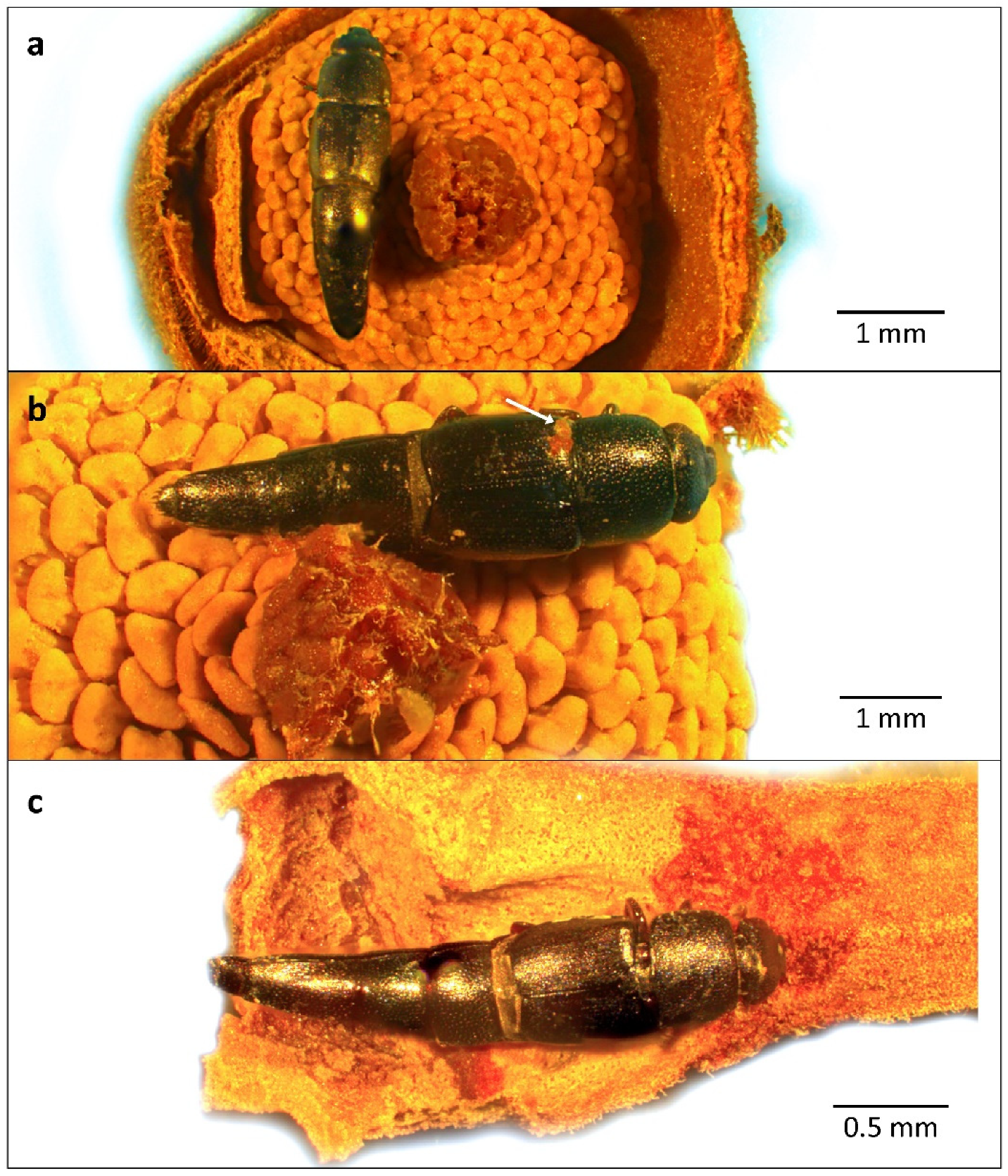
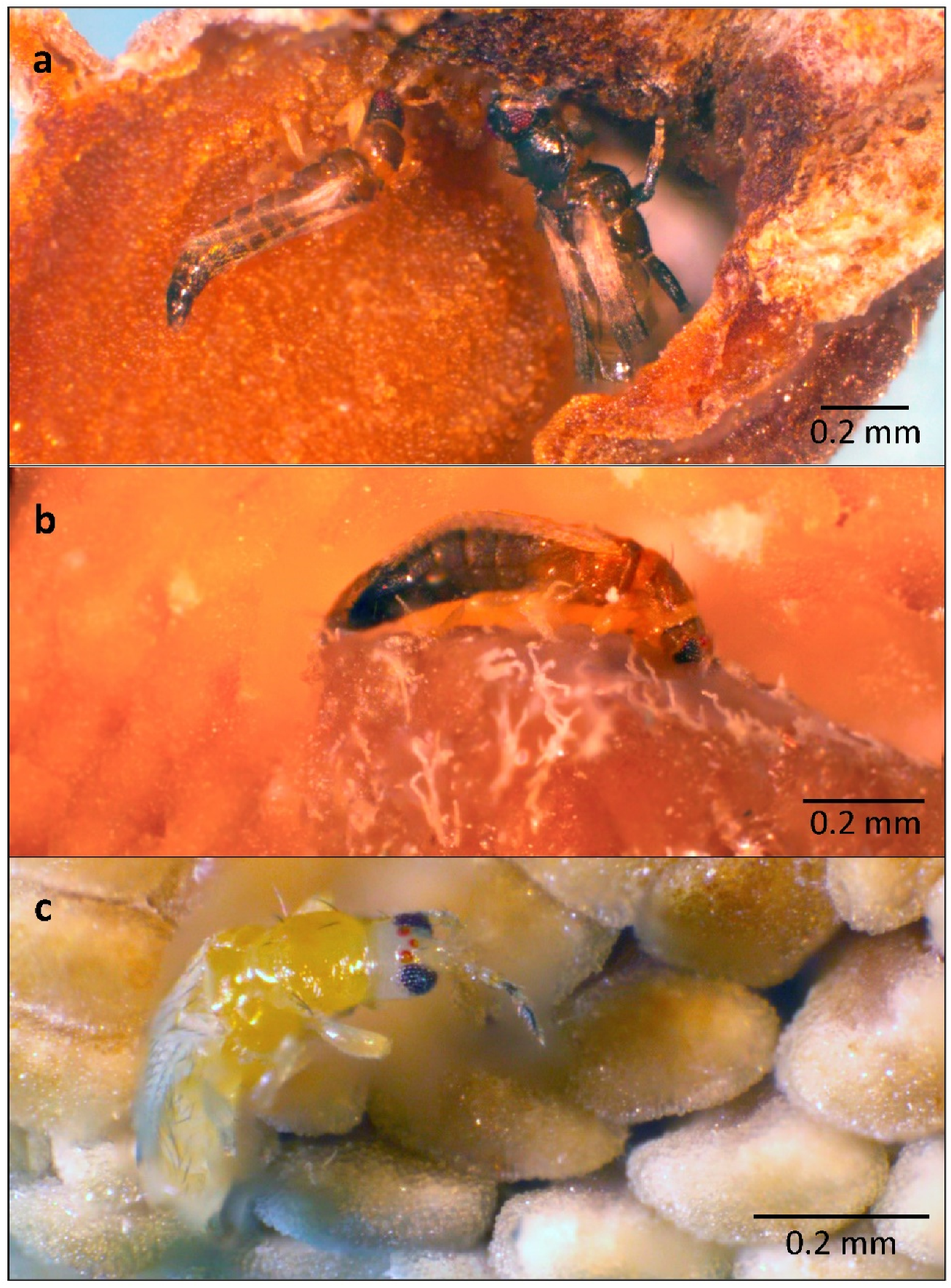
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Del-Claro, K.; Johnson, M.; Torezan-Silingardi, H.M. Plant-Arthropod Interactions: A Behavioral Approach. Psyche 2012, 2012, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Hughes, N.F. Palaeobiology of Angiosperm Origins: Problems of Mesozoic Seed-Plant Evolution; Cambridge University Press: New York, NY, USA, 1976. [Google Scholar]
- Crowson, R.A. The Biology of the Coleoptera; Academic Press: London, UK, 1981. [Google Scholar]
- Jolivet, P. Food habits and food selection of Chrysomelidae: Bionomic and evolutionary perspectives. In Biology of Chrysome-lidae; Jolivet, P., Petitpierre, E., Hsiao, T.H., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1988; pp. 1–24. [Google Scholar]
- Bartelt, R.J.; Kyhl, J.F.; Ambourn, A.K.; Juzwik, J.; Seybold, S.J. Male-produced aggregation pheromone of Carpophilus sayi, a nitidulid vector of oak wilt disease, and pheromonal comparison with Carpophilus lugubris. Agric. For. Entomol. 2004, 6, 39–46. [Google Scholar] [CrossRef]
- Cline, A.R.; Kinnee, S.A. A new species of sap beetle (Coleoptera: Nitidulidae) from Baja California Sur, Mexico, with a review of the genus Lobiopa Erichson. Pan-Pac. Entomol. 2012, 88, 202–211. [Google Scholar] [CrossRef]
- Fernandes, D.R.R.; Bená, D.C.; Lara, R.I.R.; Ide, S.; Perioto, N.W. Nitidulidae (Coleoptera) associados a frutos de café (Coffea arabica L.). Coffee Sci. 2012, 7, 135–138. [Google Scholar]
- Oliveira, M.A.S.; Genú, P.J.C.; Junqueira, M.T.V.; Pinto, A.C.Q. Pragas da Gravioleira no Cerrado; Embrapa-CPAC: Planaltina, Brazil, 1992; pp. 1–11. [Google Scholar]
- Hausmann, C.; Samietz, J.; Dorn, S. Visual Orientation of Overwintered Anthonomus pomorum (Coleoptera: Curculionidae). Environ. Entomol. 2004, 33, 1410–1415. [Google Scholar] [CrossRef] [Green Version]
- Vanin, S.A.; Bená, D.D.C.; Albertoni, F.F. Description of immature stages of Phelypera schuppeli (Boheman, 1834) with comments on natural history (Coleoptera: Curculionidae: Hyperinae). Zootaxa 2012, 3423, 45–60. [Google Scholar] [CrossRef]
- Oliveira, L.J.; Santos, B.; Parra, J.R.P.; Amaral, M.L.B.D.; Magri, D.C. Ciclo Biológico de Phyllophaga cuyabana (Moser) (Scarabaeidae: Melolonthinae). An. Soc. Entomol. Bras. 1996, 25, 431–437. [Google Scholar] [CrossRef]
- Rodrigues, S.R.; Barbosa, C.A.F.; Fuhrmann, J.; Amaro, R.A. Mating behavior and description of immature stages of Cyclocephala melanocephala (Fabricius, 1775) (Coleoptera: Scarabaeidae: Dynastinae), identification key and remarks on known immatures of Cyclocephalini species. Rev. Bras. Entomol. 2018, 62, 205–219. [Google Scholar] [CrossRef]
- Barfod, A.S.; Hagen, M.; Borchsenius, F. Twenty-five years of progress in understanding pollination mechanisms in palms (Arecaceae). Ann. Bot. 2011, 108, 1503–1516. [Google Scholar] [CrossRef] [Green Version]
- Elias, M.A.S.; Franceschinelli, E.V.; Juen, L.; Borges, F.J.A.; Ferreira, G.M.; Carvalho, F.M.V. Reproductive success of Cardiopetalum calophyllum (Annonaceae) treelets in fragments of Brazilian savanna. J. Trop. Ecol. 2012, 28, 317–320. [Google Scholar] [CrossRef]
- Teichert, H.; Dötterl, S.; Frame, D.; Kirejtshuk, A.; Gottsberger, G. A novel pollination mode, saprocantharophily, in Duguetia cadaverica (Annonaceae): A stinkhorn (Phallales) flower mimic. Flora 2012, 207, 522–529. [Google Scholar] [CrossRef]
- Tsukada, M.; Inui, M.; Suzaki, N. Do beetles prefer the odor of female-stage to male-stage flowers in Atemoya, a cantharo-phylous protogynous fruit tree (Annonaceae)? J. Entomol. Res. Soc. 2017, 19, 43–52. [Google Scholar]
- Saravy, F.P.; Marques, M.I.; Schuchmann, K.-L. Coleopteran Pollinators of Annonaceae in the Brazilian Cerrado—A Review. Diversity 2021, 13, 438. [Google Scholar] [CrossRef]
- Silberbauer-Gottsberger, I. Pollination and evolution in palms. Phyton 1990, 30, 213–233. [Google Scholar]
- Eriksson, R. The remarkable weevil pollination of the neotropical Carludovicoideae (Cyclanthaceae). Plant Syst. Evol. 1994, 189, 75–81. [Google Scholar] [CrossRef]
- Lau, J.Y.Y.; Guo, X.; Pang, C.-C.; Tang, C.C.; Thomas, D.C.; Saunders, R.M.K. Time-Dependent Trapping of Pollinators Driven by the Alignment of Floral Phenology with Insect Circadian Rhythms. Front. Plant Sci. 2017, 8, 1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medeiros, B.A.S.D.; Núñez-Avellaneda, A.L.; Hernandez, A.M.; Farrell, B.D. Flower visitors of the licuri palm (Syagrus coronata): Brood pollinators coexist with a diverse community of antagonists and mutualists. Biol. J. Linn. Soc. 2019, 126, 666–687. [Google Scholar] [CrossRef]
- Gibernau, M.; Barabé, D.; Cerdan, P.; Dejean, A. Beetle pollination of Philodendron solimoesense (Araceae) in French Guiana. Int. J. Plant Sci. 1999, 160, 1135–1143. [Google Scholar] [CrossRef] [Green Version]
- Gottsberger, G. Pollination and evolution in neotropical Annonaceae. Plant Species Biol. 1999, 14, 143–152. [Google Scholar] [CrossRef]
- De Almeida-Júnior, E.B.; Collevatti, R.G.; Telles, M.P.D.C.; Chaves, L.J.; Neres, D.F.; Soares, T. Short-distance pollen dispersal in a protogynous Annonaceae tree species from the Brazilian Cerrado. Plant Syst. Evol. 2018, 304, 1091–1099. [Google Scholar] [CrossRef]
- Gottsberger, G. How diverse are Annonaceae with regard to pollination? Bot. J. Linn. Soc. 2012, 169, 245–261. [Google Scholar] [CrossRef] [Green Version]
- Souza, V.C.; Lorenzi, H. Botânica Sistemática: Guia Ilustrado para Identificação das Famílias Nativas e Exóticas no Brasil, Baseado em APG III; Instituto Plantarum: Nova Odessa, Brazil, 2012. [Google Scholar]
- Couvreur, T.L.P.; Helmstetter, A.J.; Koenen, E.J.M.; Bethune, K.; Brandão, R.D.; Little, S.A.; Sauquet, H.; Erkens, R.H.J. Phy-logenomics of the major tropical plant family Annonaceae using targeted enrichment of nuclear genes. Front. Plant Sci. 2019, 9, 1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottsberger, G. As anonáceas do cerrado e a sua polinização. Braz. J. Biol. 1994, 54, 391–402. [Google Scholar]
- Costa, M.S.; Silva, R.J.; Paulino-Neto, H.F.; Pereira, M.J.B. Beetle pollination and flowering rhythm of Annona coriacea Mart. (Annonaceae) in Brazilian cerrado: Behavioral features of its principal pollinators. PLoS ONE 2017, 12, e0171092. [Google Scholar] [CrossRef] [PubMed]
- Parizotto, D.R.; Grossi, P.C. Revisiting pollinating Cyclocephala scarab beetles (Coleoptera: Melolonthidae: Dynastinae) as-sociated with the soursop (Annona muricata, Annonaceae). Neotrop. Entomol. 2018, 48, 415–421. [Google Scholar] [CrossRef]
- Forzza, R.C.; Baumgratz, J.F.A.; Costa, A.; Hopkins, M.; Leitman, P.M.; Lohmann, L.G.; Martinelli, G.; Morim, M.P.; Coelho, M.A.N.; Peixoto, A.L.; et al. As Angiospermas do Brasil. In Catálogo de Plantas e Fungos do Brasil; Forzza, R.C., Leitman, P.M., Costa, A., de Carvalho, A.A., Jr., Peixoto, A.L., Walter, B.M.T., Bicudo, C., Zappi, D., da Costa, D.P., Lleras, E., et al., Eds.; Instituto de Pesquisas do Jardim Botânico do Rio de Janeiro: Rio de Janeiro, Brazil, 2010; Volume 1, pp. 78–89. [Google Scholar]
- Mendonça, R.C.; Felfili, J.F.; Walter, B.M.T.; Silva-Júnior, M.C.; Rezende, A.V.; Filgueiras, T.S.; Nogueira, P.E.; Fagg, C.F. Flora vascular do bioma Cerrado–Checklist com 12.356 espécies. In Cerrado-Ecologia e Flora; Sano, S.M., de Almeida, S.P., Ribeiro, J.F., Eds.; Embrapa Informação Tecnológica: Brasília, Brazil, 2008; Volume 2, pp. 421–1181. [Google Scholar]
- Ratnayake, R.M.C.S.; Gunatilleke, I.A.U.N.; Wijesundara, D.S.A.; Saunders, R.M.K. Pollination Ecology and Breeding System of Xylopia championii (Annonaceae): Curculionid Beetle Pollination, Promoted by Floral Scents and Elevated Floral Temperatures. Int. J. Plant Sci. 2007, 168, 1255–1268. [Google Scholar] [CrossRef] [Green Version]
- Aragão, D.D.S.; Costa, C.B.N.; Nascimento, V.T.D. Biologia floral, fenologia reprodutiva e polinização de Xylopia aromatica (Lam.) Mart. (Annonaceae) em uma área de Cerrado no oeste da Bahia. Paubrasilia 2019, 2, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Gottsberger, G.; Silberbauer-Gottsberger, I. In the evening when the beetles come: Pollination in Annonaceae and Philodendron. In Life in the Cerrado: A South American Tropical Seasonal Vegetation; Reta Verlag: Ulm, Germany, 2006; Volume 2, Chapter 15; pp. 138–158. [Google Scholar]
- GBIF-Global Biodiversity Information Facility. Xylopia aromatica (Lam.) Mart. Available online: https://www.gbif.org (accessed on 26 September 2021).
- Ribeiro, J.F.; Walter, B.M.T. As principais fitofisionomias do bioma Cerrado. In Cerrado: Ecologia e Flora; Sano, S.M., de Almeida, S.P., Ribeiro, J.F., Eds.; Embrapa Informação Tecnológica: Brasília, Brazil, 2008; Volume 1, pp. 151–212. [Google Scholar]
- Vieira-Júnior, H.T.; Moraes, J.M.; de Paula, T.L.F. Chapada dos Guimarães (MT). In Geoparques do Brasil: Propostas; Schobben-haus, C., da Silva, C.R., Eds.; CPRM: Rio de Janeiro, Brazil, 2012; Volume 1, pp. 283–316. [Google Scholar]
- Brusca, R.C.; Moore, W.; Schuster, S.M. Invertebrates, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 2016. [Google Scholar]
- Bouchard, P.; Bousquet, Y.; Davies, A.; Alonso-Zarazaga, M.; Lawrence, J.; Lyal, C.; Newton, A.; Reid, C.; Schmitt, M.; Slipinski, A.; et al. Family-Group Names In Coleoptera (Insecta). ZooKeys 2011, 88, 1. [Google Scholar] [CrossRef] [Green Version]
- Habeck, D.H. Nitidulidae Latreille. In American Beetles-Polyphaga: Scarabaeoidea through Curculionoidea; Arnett, R.H., Thomas, M.C., Skelley, P.E., Frank, J.H., Eds.; CRC Press: Boca Raton, CA, USA, 2002; Volume 2, pp. 311–315. [Google Scholar]
- Dieringer, G.; Cabrera, R.L.; Lara, M.; Loya, L.; Reyes-Castillo, P. Beetle Pollination and Floral Thermogenicity in Magnolia tamaulipana (Magnoliaceae). Int. J. Plant Sci. 1999, 160, 64–71. [Google Scholar] [CrossRef]
- Ervik, F.; Knudsen, J.T. Water lilies and scarabs: Faithful partners for 100 million years? Biol. J. Linn. Soc. 2003, 80, 539–543. [Google Scholar] [CrossRef] [Green Version]
- Paulino-Neto, H.F. Polinização por besouros. In Biologia da Polinização; Rech, A.R., Agostini, K., Oliveira, P.E., Machado, I.C., Eds.; Projeto Cultural: Rio de Janeiro, Brazil, 2014; pp. 259–275. [Google Scholar]
- Küchmeister, H.; Webber, A.C.; Silberbauer-Gottsberger, I.; Gottsberger, G. A polinização e sua relação com a termogênese em espécies de Arecaceae e Annonaceae da Amazônia Central. Acta Amazon. 1998, 28, 217–245. [Google Scholar] [CrossRef] [Green Version]
- Jürgens, A.; Webber, A.C.; Gottsberger, G. Floral scent compounds of Amazonian Annonaceae species pollinated by small beetles and thrips. Phytochemistry 2000, 55, 551–558. [Google Scholar] [CrossRef]
- Goodrich, K.R. Floral scent in Annonaceae. Bot. J. Linn. Soc. 2012, 169, 262–279. [Google Scholar] [CrossRef] [Green Version]
- Ervik, F.; Tollsten, L.; Knudsen, J.T. Floral scent chemistry and pollination ecology in phytelephantoid palms (Arecaceae). Plant Syst. Evol. 1999, 217, 279–297. [Google Scholar] [CrossRef]
- Dowd, P.F.; Bartelt, R.J. Aggregation pheromone glands of Carpophilus freemani (Coleoptera: Nitidulidae) and gland distri-bution among other sap beetles. Ann. Entomol. Soc. Am. 1993, 86, 464–469. [Google Scholar] [CrossRef]
- Kono, M.; Tobe, H. Is Cycas revoluta (Cycadaceae) wind- or insect-pollinated? Am. J. Bot. 2007, 94, 847–855. [Google Scholar] [CrossRef]
- Milet-Pinheiro, P.; Gonçalves, E.G.; Navarro, D.M.D.A.F.; Nuñez-Avellaneda, L.A.; Maia, A. Floral scent chemistry and pollination in the Neotropical aroid genus Xanthosoma (Araceae). Flora 2017, 231, 1–10. [Google Scholar] [CrossRef]
- Hemborg, A.M.; Bond, W.J. Different rewards in female and male flowers can explain the evolution of sexual dimorphism in plants. Biol. J. Linn. Soc. 2005, 85, 97–109. [Google Scholar] [CrossRef]
- Meléndez, M.R.; Ponce, W.P. Pollination in the oil palms Elaeis guineensis, E. oleifera and their hybrids (OxG), in tropical America. Pesqui. Agropecuária Trop. 2016, 46, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Kirejtshuk, A.G. On polyphyly of the Carpophilinae with description of a new subfamily, Cillaeinae (Coleoptera: Nitidulidae). Coleopt. Bull. 1986, 40, 217–221. [Google Scholar]
- Kirejtshuk, A.G.; Kovalev, A.V. A new genus of the subfamily Cillaeinae (Coleoptera, Nitidulidae) from the Philippines and New Guinea with notes on the taxonomy and phylogeny of the subfamily. Zootaxa 2016, 4205, 226. [Google Scholar] [CrossRef]
- Paulino-Neto, H.F. História Natural e Interação Flores-Besouros em Espécies de Cerrado. Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil, 2009. [Google Scholar]
- Lau, J.Y.Y.; Pang, C.C.; Ramsden, L.; Saunders, R.M.K. Stigmatic exudate in the Annonaceae: Pollinator reward, pollen ger-mination medium or extragynoecial compitum? J. Integr. Plant Biol. 2017, 59, 881–894. [Google Scholar] [CrossRef] [Green Version]
- Gottsberger, G. Comments on flower evolution and beetle pollination in the genera Annona and Rollinia (Annonaceae). Plant Syst. Evol. 1989, 167, 189–194. [Google Scholar] [CrossRef]
- Casari, S.A.; Ide, S. Coleoptera Linnaeus. In Insetos do Brasil: Diversidade e Taxonomia; Albertino, J.A., Melo, G.A.R., de Carvalho, C.J.B., Casari, S.A., Constantino, R., Eds.; Holos Editora: Ribeirão Preto, Brazil, 2012; pp. 454–535. [Google Scholar]
- Asenjo, A.; Klimaszewski, J.; Chandler, D.S.; Fierros-López, H.E.; Vieira, J.S. Staphylinidae (Insecta: Coleoptera) in Latin America: Synopsis, annotated catalog, diversity and distribution. Zootaxa 2019, 4621, 1–406. [Google Scholar] [CrossRef]
- Niño-Pérez, A.E.; Núñez-Avellaneda, L.A. Diversidad y especificidad de estafilinidos (Coleoptera: Staphylinidae) asociados con inflorescencias de palmas silvestres en el Pacífico colombiano. Rev. Biodivers. Neotrop. 2018, 8, 94–107. [Google Scholar]
- Bukejs, A.; Nadein, K. First fossil Lamprosomatinae leaf beetles (Coleoptera: Chrysomelidae) with descriptions of new genera and species from Baltic amber. Zootaxa 2015, 3931, 127–139. [Google Scholar] [CrossRef]
- Webber, A.C. Alguns aspectos da biologia floral de Annona sericea Dun. (Annonaceae). Acta Amaz. 1981, 11, 61–65. [Google Scholar] [CrossRef] [Green Version]
- Andrade, B.M.; Oliveira-Filho, A.T.; Soares, A.R. Pollination and breeding system of Xylopia brasiliensis Sprengel (Annonaceae) in south-eastern Brazil. J. Trop. Ecol. 1996, 12, 313–320. [Google Scholar] [CrossRef]
- Monteiro, R.C.; Mound, L.A. Thysanoptera Haliday. In Insetos do Brasil: Diversidade e Taxonomia; Albertino, J.A., Melo, G.A.R., de Carvalho, C.J.B., Casari, S.A., Constantino, R., Eds.; Holos Editora: Ribeirão Preto, Brazil, 2012; pp. 408–422. [Google Scholar]
- Lorenzi, H. Árvores Brasileiras: Manual de Identificação e Cultivo de Plantas Arbóreas Nativas do Brasil; Plantarum: Nova Odessa, Brazil, 1992. [Google Scholar]
- Silva-Júnior, M.C.; Dos Santos, G.C.; Nogueira, P.E.; Munhoz, C.B.R.; Ramos, A.E. 100 Árvores do Cerrado: Guia de Campo; Rede de Sementes do Cerrado: Brasília, Brazil, 2005. [Google Scholar]
- Silva, N.A.P.; Frizzas, M.A.; Oliveira, C.M. Seasonality in insect abundance in the “Cerrado” of Goiás State, Brazil. Rev. Bras. Entomol. 2011, 55, 79–87. [Google Scholar] [CrossRef] [Green Version]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Floral Phase | No. of Flowers | % of Flowers | Time of Collection | |
|---|---|---|---|---|
| Female | 234 | 94.46 | All hours | (1st day) |
| Male | 19 | 5.54 | 07:05 to 16:05 | (2nd day) |
| Family/Subfamily/Species | Ni | Nf (%) | Ni/f | P.F.C. (%) |
|---|---|---|---|---|
| Nitidulidae | ||||
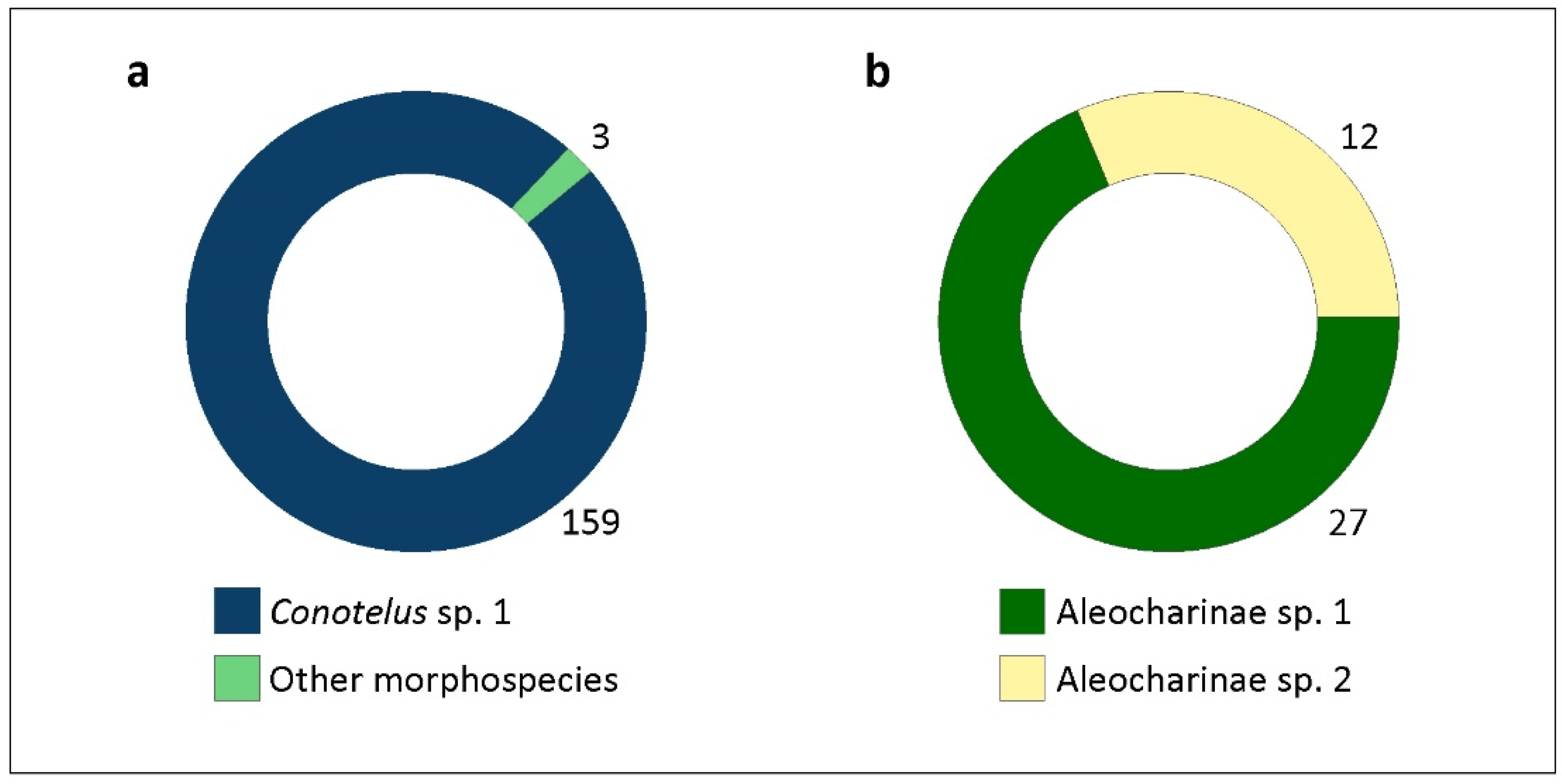
| Conotelus sp. 1 | 159 | 33.2 | 1.91 | 83.64 |
| Carpophilinae sp. 1 | 1 | 0.39 | 1 | 100 |
| Nitidulinae sp. 1 | 1 | 0.39 | 1 | 1 |
| Meligethinae sp. 1 | 1 | 0.39 | 1 | 100 |
| Staphylinidae | ||||
| Aleocharinae sp. 1 | 27 | 8.3 | 1.28 | 70.37 |
| Aleocharinae sp. 2 | 12 | 4.74 | 1 | 91.66 |
| Chrysomelidae | ||||
| Lamprosomatinae sp. 1 | 2 | 0.79 | 1 | 100 |
| Lamprosomatinae sp. 2 | 4 | 1.18 | 1.33 | 0 |
| Chrysomelidae sp. 1 | 1 | 0.39 | 1 | 100 |
| Chrysomelinae sp. 1 | 1 | 0.39 | 1 | 0 |
| Curculionidae | ||||
| Curculionidae sp. 1 | 1 | 0.39 | 1 | 0 |
| Total | 210 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinheiro Saravy, F.; Schuchmann, K.-L.; Marques, M.I. Diversity of Insect Flower Visitors of Xylopia aromatica (Magnoliales, Annonaceae) in a Brazilian Savanna. Diversity 2021, 13, 661. https://doi.org/10.3390/d13120661
Pinheiro Saravy F, Schuchmann K-L, Marques MI. Diversity of Insect Flower Visitors of Xylopia aromatica (Magnoliales, Annonaceae) in a Brazilian Savanna. Diversity. 2021; 13(12):661. https://doi.org/10.3390/d13120661
Chicago/Turabian StylePinheiro Saravy, Fábio, Karl-L. Schuchmann, and Marinez I. Marques. 2021. "Diversity of Insect Flower Visitors of Xylopia aromatica (Magnoliales, Annonaceae) in a Brazilian Savanna" Diversity 13, no. 12: 661. https://doi.org/10.3390/d13120661
APA StylePinheiro Saravy, F., Schuchmann, K. -L., & Marques, M. I. (2021). Diversity of Insect Flower Visitors of Xylopia aromatica (Magnoliales, Annonaceae) in a Brazilian Savanna. Diversity, 13(12), 661. https://doi.org/10.3390/d13120661